БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
2. СТРОЕНИЕ И УЛЬТРАСТРУКТУРА КЛЕТКИ
2.4. Теория симбиогенеза и водородная гипотеза
Пластиды и митохондрии занимают в клетках эукариот особое положение: они постоянно отделены оболочкой из двух мембран от цитоплазмы и сливаются (насколько это вообще бывает возможно) только с себе подобными. Они имеют собственную кольцевую ДНК, а также собственные молекулярные комплексы для транскрипции и трансляции, которые показывают бактериальные признаки (см. ниже). Характер деления также напоминает деление бактерий. Теория симбиогенеза смогла эти особенности объяснить исходя из предположения, что митохондрии и пластиды произошли от бактерий, которые на раннем этапе эволюции жизни на Земле могли внедриться в качестве внутриклеточных симбионтов в примитивные эукариотические клетки1 (эндосимбионтов). На современных примерах эндосимбиоза можно проверить эту теорию возникновения пластид и митохондрий.
1 Теория эндосимбиоза впервые предложена русским ученым М.Д. Мережковским, дальнейшее развитие она получила в трудах американской исследовательницы Маргелис. — Примеч. ред.
2.4.1. Эндосимбиоз
У многих протистов, грибов и растений обнаруживаются эндосимбионты, которые в клетках своих хозяев играют роль органелл. Например, бактерии из родов Rhizobium и Bradyrhizobium, когда они живут в клетках корневых клубеньков бобовых растений, ассимилируют азот из воздуха и таким образом делают свое растение-хозяина независимым от почвенного азота или азотных удобрений (см. 2.2.1). У мадрепоровых кораллов эндосимбионтные динофлагелляты (зооксантеллы) (см. рис. 11.69), осуществляя фотосинтез, ускоряют рост животного-хозяина примерно в десять раз. У амеб, различных инфузорий, у грибов и пресноводных полипов Hydra имеются формы, в которых одноклеточные эндосимбиотические зеленые водоросли могут осуществлять фотосинтез и которые за счет этого частично или полностью могут становиться фотоавтотрофными. Образование стабильных эндосимбиозов в любом случае — широко распространенное, экологически значительное явление у современных организмов (см. 9.2).
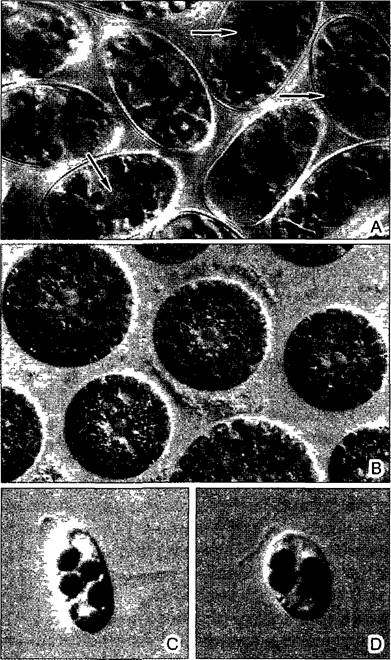
Некоторые эндосимбионты могут выживать независимо от своих хозяев. В других случаях взаимная зависимость партнеров по симбиозу друг от друга столь сильна, что в природе они могут встречаться только совместно. Крайний пример такой ситуации — синцианозы, в которых одноклеточные цианобактерии живут как постоянные внутриклеточные симбионты (рис. 2.99). Эндосимбиотические цианобактерии играют роль хлоропластов. Их называют цианеллы (греч. kyanos — синий). Ци- анеллы не могут выживать вне своих хозяев. Их ДНК имеет лишь 1/10 длины и соответственно информационной емкости генома свободноживущих цианобактерий. Большинство специфичных для цианелл белков кодируется не этой ДНК, а ядерной ДНК клеток хозяина. Этим у цианелл, которые еще сохраняют остатки прокариотной клеточной стенки, достигается ситуация, которая и в генетическом плане соответствует таковой у пластид.
Рис. 2.99. Синцианозы: А— aucocystis nostochinearum с цианеллами вытянутой формы; стрелки — клеточные ядра (900х); В—Glaucosphaera vacuolata с округлыми цианеллами, в которых хорошо различимы периферическая хроматоплазма и непигментиpованная центроплазма; в центре сидячих клеток — ядро с ядрышком (900х); С, D—жгутиконосец Cyanophora paradoxa с цианеллами, которые находятся на разных стадиях деления (1700х)
2.4.2. Возникновение пластид и митохондрий путем симбиогенеза
Теория симбиогенеза основывается, как уже упоминалось, прежде всего на ряде особых признаков пластид и митохондрий, которые наблюдаются также и у бактерий:
✵ кольцевая ДНК без часто повторяющихся последовательностей, сконцентрированная в нуклеоидах, без гистонов и соответственно нуклеосом;
✵ репликация независимо от времени S-фазы клеточного цикла;
✵ сходство последовательностей (например, рРНК у митохондрий с α-пурпурными бактериями, у пластид с цианобактериями);
✵ только одна чувствительная к рифамици- ну РНК-полимераза (в клеточном ядре их, напротив, три, и они в разной степени чувствительны к аманитину);
✵ на мРНК: нет кэпирования у 5’-конца, нет поли-А-последовательности у З'-конца (см. 7.2.2.2);
✵ рибосомы, которые (в том числе) по размеру и чувствительности к ингибиторам соответствуют бактериальному 70S-типy;
✵ начало трансляции с формилметионина (а не с метионина, как у цитоплазматических 80S-рибосом)
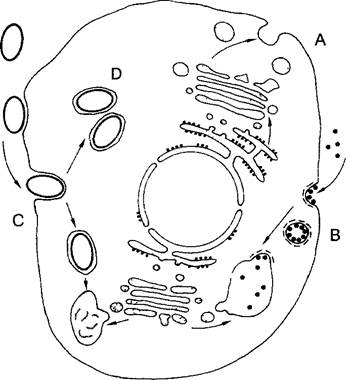
Помимо указанных, имеются явные признаки родства органелл с бактериями. Внутренняя мембрана митохондрий содержит, например, встречающийся помимо этого только у бактерий кардиолипин, тогда как в ней отсутствуют типичные для эукариотических мембран стероидные липиды (рис. 2.100). Постулируемое теорией симбиогенеза внедрение эндосимбионтов, по-видимому, происходило путем фагоцитоза — распространенного у простейших (но также, например, и у гранулопитов и макрофагов млекопитающих и человека) механизма поглощения твердых частиц (рис. 2.101). При фагоцитозе неизбежно возникает компартментация, известная у пластид и митохондрий: заглатываемые путем фагоцитоза клетки в заглотившей их клетке окружаются оболочкой из двух мембран, причем внутренняя мембрана соответствует плазматической мембране захваченной клетки, а внешняя, напротив, мембране фагосомы (эндосомы), которая в свою очередь произошла из плазматической мембраны захватившей клетки. После фагоцитоза захваченная частица (пищи) обычно переваривается лизосомами (см. рис. 2.56, А). Однако это не происходит при установлении эндосимбиозов — здесь захваченные одноклеточные организмы выживают в клетке хозяина в качестве симбионтов или же паразитов, что подтверждают и подобные случаи среди ныне живущих организмов (см. выше).
Рис. 2.100. Кардиолипин (А)—фосфолипид, встречающийся у свободноживущих бактерий. В эукариотических клетках его можно обнаружить только во внутренней мембране митохондрий. Стероидные липиды — например, холестерин (В)—наоборот, можно обнаружить только в составе эукариотических мембран, у свободноживущих бактерий и во внутренней мембране митохондрий он отсутствует
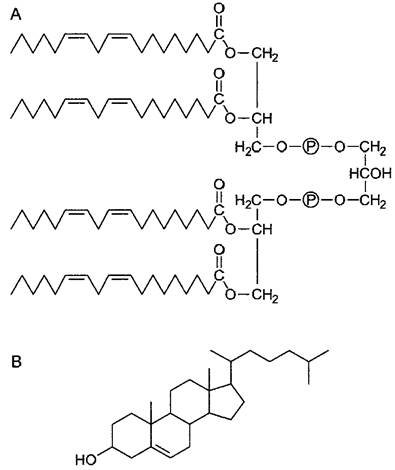
Рис. 2.101. Фагоцитоз и эндосимбиоз. Эукариотический фагоцит (например, амеба) способен путем экзо- и эндоцитоза обмениваться со средой мелкими частицами и молекулами (А, В), а также может поглощать целые клетки (показано тонировкой), образуя в процессе фагоцитоза сначала складки наружной мембраны (С), а затем — пищеварительные вакуоли (фагосомы) При слиянии с первичными лизосомами в вакуолях происходит переваривание захваченных клеток. Если внедрившаяся клетка выживает, может образоваться стабильный симбиоз (D) с клеткой-хозяином, где происходит размножение захваченных клеток, играющих роль симбионтов (или паразитов)
Гипотетические организмы, в клетки которых встроились гипотетические предки содержащих ДНК органелл, называют протоэукариотами. Фактически еще и ныне известны простейшие, которые не имеют митохондрий и показывают примитивные особенности последовательностей ДНК. Такие архезои соответствуют этим гипотетическим протоэукариотам. К ним относятся систематические группы Мetamonada, Мicrosporidiа и Раrabasaliа. В последнее время показано, что ядерная ДНК архезой содержит участки последовательностей, которые могли произойти только от митохондрий. Эти паразитические одноклеточные организмы, возможно, сначала имели митохондрии и вторично их утратили.
В настоящее время обсуждается и альтернативная гипотеза — так называемая водородная гипотеза. В основе ее лежит положение, что протоэукариот в качестве самостоятельной эволюционной линии на ранних этапах развития жизни на Земле вообще не было, а первые эукариотические клетки уже были продуктом симбиоза метаногенных архебактерий и α-протео- бактерий. Протеобактерии при недостатке кислорода образуют водород, в котором архебактерии нуждаются для продукции метана. Таким образом, симбиоз мог сделать архебактерии независимыми от абиотических источников Н2. Путем обрастания архебактерии могли затем полностью присоединить к себе своих партнеров. В свою очередь эти партнеры в ходе дальнейшей эволюции либо развивались в гидрогеносомы (эквиваленты митохондрий у анаэробных одноклеточных эукариот), либо — при доступности O2 — в митохондрии. Согласно водородной гипотезе, в состав клеток примитивных эукариот должны были входить α-протеобактерии. В противоположность теории симбиогенеза водородная гипотеза не предполагает внедрения симбионтов путем фагоцитоза. Водородная гипотеза, наряду с прочим, основывается на том, что именно метаногенные архебактерии (как и эукариоты) содержат гистоны и образуют нуклеосомы.
Важное общее следствие из теории симбиогенеза состоит в том, что организмы нового типа возникли в ходе эволюции не за счет мутации, генетической рекомбинации или горизонтального переноса генов, а путем образования стабильных внутриклеточных симбиозов. Новые надорганизмы, возникшие путем такой межтаксонной комбинации,представляют собой в клеточном и генетическом плане химеры. (Химера в греческих мифах — это чудовище с головой льва, туловищем козы и хвостом дракона. В биологии химерой называют генетически неоднородный организм.) Современные эукариотические клетки представляют собой мозаичные клетки, составленные из клеток представителей разных царств (империй) организмов. При очень длительной совместной эволюции клеток хозяина и эндоцитоби- онтов — называемой симбиогенезом — симбионты постепенно превращались в органеллы, которые наблюдаются у современных эукариот. К изменениям, наряду с прочим, относятся утрата стенки, приуроченность размножения к специфическим потребностям клетки хозяина; развитие транслокаторных систем в мембранах оболочек для интенсивного обмена веществами вплоть до появления способности выводить АТФ или триозофосфаты через эти мембраны; и, наконец, передача генетической информации из симбионтов/органелл в ядра хозяина, в комбинации со специфическим импортом белков (и тРНК) из цитоплазмы в органеллы.