БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
2. СТРОЕНИЕ И УЛЬТРАСТРУКТУРА КЛЕТКИ
2.3. Строение прокариотической клетки
Прокариоты в экологическом, физиологическом и структурном плане очень гетерогенны. Ниже приведем только беглый обзор общих особенностей строения клетки прокариот, который тем не менее ясно покажет, сколь велики различия между прокариотическими и эукариотическими клетками. Никаких переходных форм между этими двумя типами клеток в современном мире организмов неизвестно. Только сходство последовательностей семантических молекул (ДНК, РНК, белков) указывают на общее происхождение в ходе эволюции про- и эукариот.
Уже внешне принципиальное различие между про- и эукариотами проявляется в сильно различающихся размерах клеток (см. рис. 2). Размеры клетки кишечной палочки Escherichia coli составляют 2 — 4x1 мкм, а соответственно объем — около 2,5 мкм3. Объем цитоплазмы средней эукариотической клетки без вакуолей составляет порядка 1500 — 3 000 мкм3, оказываясь, таким образом, примерно на 3 порядка больше. Этому соответствует и значительно меньшее содержание ДНК в
прокариотической клетке. Если общая длина ядерной ДНК в гаплоидной клетке человека приближается к 1 м, то у Е. coli она едва превышает 1 мм. С малыми размерами прокариотических клеток связано и то, что продолжительность жизни одного поколения может быть очень мала; у Е. соli, например, 20 мин. Эукариотические клетки в образовательных тканях, напротив, делятся в большинстве случаев не чаще чем один раз в сутки. Из одной-единственной бактериальной клетки уже через 10 ч может возникнуть свыше миллиарда клеток — обстоятельство, которое наряду с прочими делает понятным огромное экологическое значение бактерий. Настоящих многоклеточных организмов средипрокариот нет.
Малые размеры прокариот имеют следствием особенно простую внутреннюю дифференцировку клетки. В столь малой клетке не было бы места обширной внутренней мембранной системе (рис 2.93). В большинстве прокариотических клеток плазматическая мембрана является единственной биомембраной, клетка представляет собой один-единственный компартмент Внутриклеточные неплазматические компартменты у бактерий образуются редко, и в каждом случае нет ни ЭР, ни диктиосом, ни пузырьков, ни настоящих вакуолей, а также таких окруженных мембранами компартментов, как пластиды или митохондрии.
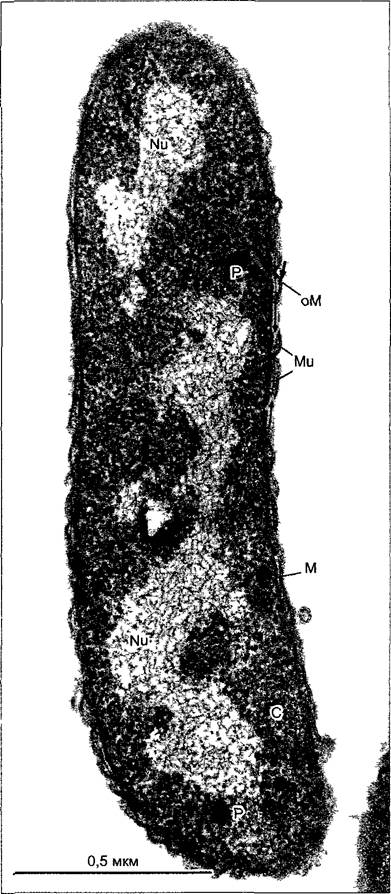
Рис. 2.93. Ультраструктура типичной бактериальной (грамотрицательной) клетки Rhodo- spirillum rubrum (препарат R.Ladwig, электронная микрофотосъемка R.Marx).
Аморфный нуклеоид, в котором четко различимы тяжи ДНК, окружен богатой рибосомами цитоплазмой, в которой находятся полифосфатные гранулы. Клетка отграничена плазматической мембраной от клеточной стенки В клеточной стенке, помимо тонкого муреинового мешка (пептидогликанового слоя, тонкие стрелки) находится мембраноподобный слой — так называемая внешняя мембрана (outer membrane); она отсутствует у грамположительных бактерий, у которых муреиновая часть стенки значительно толще и многослойная (ср. рис. 2.98). С—цитоплазма; М—плазматическая мембрана; Мu—слой муреина, Nu—нуклеоид, оМ—внешняя (outer) мембрана, Р—полифосфатные гранулы
Так называемые тилакоиды цианобактерий (рис 2.94, А) не являются компартментами ограниченных мембранами пластид, как у эукариотических водорослей и высших растений Здесь, скорее, мы имеем плоские двойные мембраны в цитоплазме, которые снабжены фотосинтетическими пигментами и осуществляют световые реакции с расщеплением воды Они возникают путем выпячивания плазматической мембраны У некоторых бактерий имеются разной формы выпячивания плазматической мембраны (рис 2 94, В, С), которые, однако, длительное время остаются соединенными с ними Их называют интрацитоплазматическими мембранами (ИЦМ). Эти мембранные пузырьки, кармашки или трубочки также несут фотосинтетические пигменты.
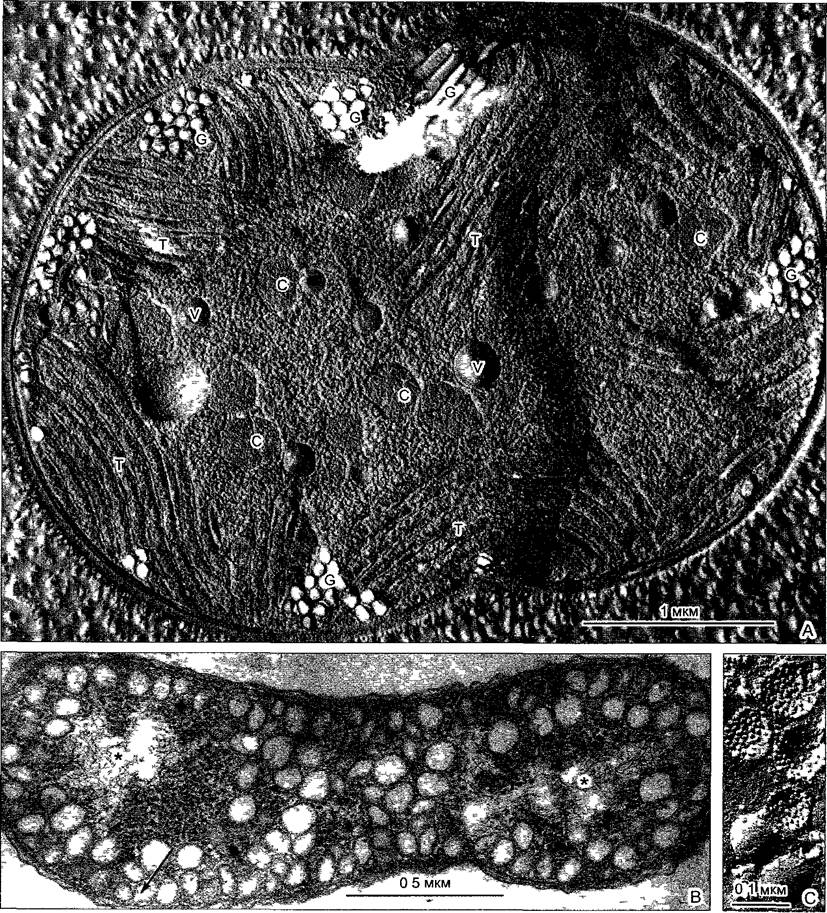
Рио. 2.94. Внутрицитоплазматические мембраны (intracytoplasmatic membranes, ICМ) прокариот (препараты и электронная микрофотосъемка J R Golecki)
А—цианобактерия Microcystis aeruginosa (здесь после замораживания-скалывания) содержит несколько типов внутрицитоплазматических мембран тилакоиды Т, вакуоли V с запасными веществами, карбоксисомы С в качестве запасных структур для фермента фотосинтеза RubisCO (см 6 5 1), так называемые газовые вакуоли G— заполненные газом цилиндрические полости, которые обеспечивают плавучесть клеток в толще воды, — окружены не липопротеиновыми мембранами, а белковыми оболочками, которые могут синтезироваться de novo в цитоплазме Клетка находится в начале деления. В—у грамотрицательной бактерии Rhodospiriffum rubrum на свету при анаэробных условиях образуется система внутрицитоплазматических мембран, похожих на хроматофоры в форме пузырьков которые осуществляют фотосинтез с помощью бактериохлорофилла (разумеется, без расщепления воды), «хроматофоры» возникают из впячиваний клеточной мембраны (стрелка) и связаны отчасти с ней, отчасти между собой Звездочкой (*) обозначены нуклеоиды С—соответствующие «хроматофоры» после замораживания-скалывания у Rhodobacter capsulatus, с поверхности (ср бокс 2 3, рис А) они выглядят гладкими, на сколе видны многие внутримембранные частицы, соответствующие пигмент-белковым комплексам для фотосинтеза
Огромное, далеко не полностью изученное многообразие прокариот выражается не только в многочисленных необычных путях обмена веществ, которые не встречаются у эукариот. Оно проявляется и в том, что имеются сильные отклонения от только что перечисленных типичных характеристик. Например, бактериальные клетки иногда достигают размеров, которые соответствуют размерам эукариотических. Максимальные значения были отмечены для Epilopiscum fishelsonit — грамположительной кишечной бактерии из тропических морских рыб, палочковидные клетки которой имеют размер 600 х 80 мкм. Еще крупнее недавно открытая в морских осадках у берегов Намибии шаровид ная серная бактерия Thiomargarita namibiensis диаметром до 750 мкм, которая в своих вакуолизированных клетках накапливает значительные количества серы и нитрата. Эта бактерия образует цепочки, объединяющие до 50 отдельных клеток. Сходные объединения клеток являются правилом у цианобактерий (см рис 5 1, 11 15), а у миксобактерий дело доходит даже до образования комплексных «плодовых тел»
2.3.1. Размножение и генетический аппарат
ДНК прокариот является кольцевой, она не располагается в разных линейных структурах, которые могли бы соответствовать хромосомам эукариот. Несмотря на это, и кольца ДНК бактерий обычно называют бактериальными хромосомами. Эти кольца ДНК имеют специфический локус для прикрепления к мембране и только одно место начала репликации они монорепликонны. Доля некодирующих участков последовательностей невелика. Несмотря на малую длину (от 0,2 мм у микоплазм до 37 мм у некоторых цианобактерий), ДНК должна быть компактно свернута, чтобы поместиться в центральной зоне клетки — нуклеоиде. Нуклеоиды не отделены простыми или двойными мембранами от содержащей рибосомы цитоплазмы, однако четко от нее отграничены. Структур типа ядрышек в нуклеоидах нет. У клеток цианобактерий (сине- зеленых водорослей), более крупных по сравнению с прочими прокариотами, расположенный в центре нуклеоид может быть виден даже при световой микроскопии. Он был описан как хромидиальный аппарат. Нуклеотид расположен в центроплазме, которая окружена хроматоплазмой, окрашенной «тилакоидами» (см. рис. 2.94, А).
В нуклеоидах прокариотических клеток гистонов нет (исключение: метаногенные архебактерии). Нейтрализация и упаковка генетического материала осуществляется другими основными белками, аминами и неорганическими катионами. При транскрипции и трансляции становится особенно ясно, что нуклеоиды не ограничены никакими мембранами: еще до завершения транскрипции одного гена или группы соседних генов (оперона) у сначала синтезированного 5'-конца тРНК уже начинается трансляция. Процессинг этой РНК не происходит. Котранскрипциональная трансляция происходит на 70S-рибосомах (субъединицы 50S и 30S; см. рис. 2.42), активность которых ингибируется антибиотиками, отличными от тех, которые действуют на 80S-рибосомы эукариот (см. 2.2.4). 70S-рибосомы меньше размером и имеют более простое строение по сравнению с 808-рибосомами, а соотношение РНК/белок у них составляет примерно 1,5: 1 (у рибосом эукариот — 1:1).
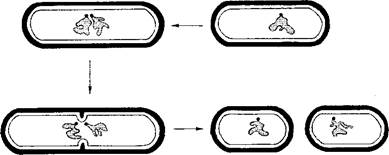
Процессов, которые бы соответствовали митозу или мейозу, у прокариот нет. Они не располагают ни микротрубочками, ни актином или миозином, и у них нет ничего, что было бы непосредственно сравнимо с аппаратом веретена у эукариот. Распределение генетического материала по дочерним клеткам достигается за счет того, что после удвоения кольцевой молекулы ДНК стартовые места репликации в нуклеоиде насколько возможно далеко расходятся друг от друга, а места прикрепления к мембране также отодвигаются друг от друга за счет роста мембраны. Между ними начинается образование септы (поперечной стенки) (рис. 2.95). Для клеточного деления цитоплазма клетки перешнуровывается в плоскости септы сократительным кольцом. В этом кольце ведущую роль играет белок FtsZ, который является гомологом тубулина и при подходящих условиях образует нити и кольцевые структуры.
Несмотря на отсутствие сингамии и мейоза, у бактерий имеются процессы типа полового, т.е. происходит перенос генетической информации от одной клетки к другой, а также рекомбинация (парасексуальность). При этом прежде всего переносятся плазмиды — (чаще всего) мелкие кольцевидные молекулы ДНК, которые могут автономно размножаться в клетке хозяина. Они содержат не гены для основного обмена веществ, а так называемые адаптивные гены, которые, например, обеспечивают устойчивость к антибиотикам (гены устойчивости в R-плазмидах), служат для конъюгации (F-плазмиды) или кодируют токсины.
Рис. 2.95. Сегрегация геномов и клеточное деление у бактерии, Схема (кольцевая ДНК и комплекс прикрепления к клеточной мембране показаны серым цветом)
2.3.2. Жгутики бактерий
Многие эубактерии имеют жгутики, которые, однако, устроены совершенно иначе, чем сложные жгутики или цилии эукариот. Бактериальный жгутик (рис. 2.96) имеет толщину только 20 нм, т.е. даже не достигает диаметра одной микротрубочки. Он построен из единственного структурного белка — флагеллина. Бактериальный жгутик спиральной формы и жесткий. У основания он имеет структуру из четырех коаксиальных колец, вставленных в плазматическую мембрану и клеточную стенку (рис. 2.97). Сам жгутик располагается вне клетки. В противоположность жгутику эукариот, в 10 раз более толстому и способному менять форму, прокариотический жгутик не одет мембраной. При плавании бактериальной клетки вперед или назад (эти направления движения постоянно сменяют друг друга) весь жгутик без изменения формы поворачивается по часовой стрелке или против нее, он действует как корабельный винт. Мотор для этого вращательного движения находится у основания жгутика. В движение он приводится не АТФ, а непосредственно протонным градиентом на плазматической мембране. При движении бактериального жгутика градиент расходуется за счет перемещения протонов в клетку.
Рис. 2.96. Бактериальные жгутики (Agroba cterium tumefaciens негативный контраст) (электронная микрофотосъемка Н Falk)
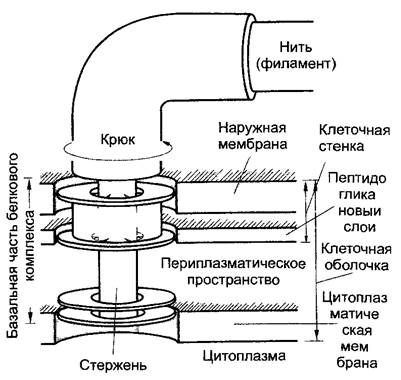
Стрелка на вставке (большее увеличение) указывает на «крючок» жгутика где находится мотор вращательного движения (рис 2.97)
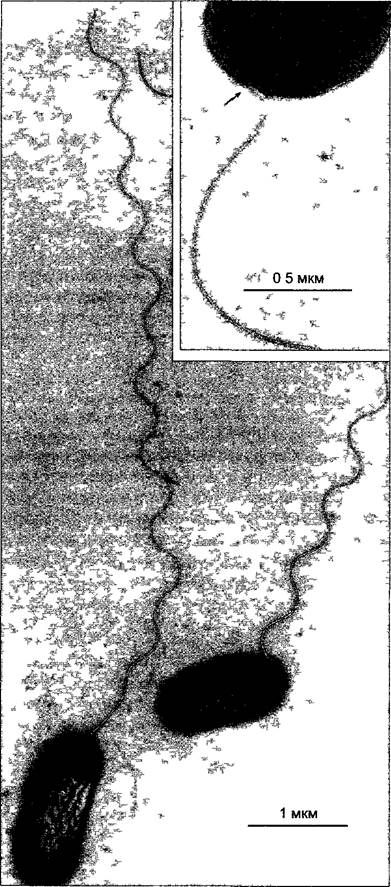
Рис. 2.97. Схема основания жгутика Eschen chia coli (по J Adler)
Четыре кольца базального белкового комплекса, который функционирует как двигательный аппарат имеют диаметр примерно 20 нм. Оба наружных кольца отсутствуют у грамположительных бактерий
2.3.3. Структуры стенки
Клеточные стенки прокариот могут выглядеть очень по-разному. У особенно мелких и просто устроенных клеток микоплазм — они представляют собой самый низкий уровень клеточной организации — клеточные стенки совсем отсутствуют. Большинство других прокариотических клеток, напротив, одеты клеточной стенкой, которая служит не только для их защиты, но и для осмотической стабилизации, придания формы и контролируемого контакта с окружающей средой. Стенка функционирует как внешний скелет. Если прокариотические клетки искусственно лишить стенки, они округляются (сферо- и протопласты), становятся осмотически лабильными и могут снова делиться только после регенерации стенки.
Рис. 2.98 показывает слоистое строение клеточных стенок у бактерий. (Клеточные стенки архебактерий сильно уклоняются как по своему строению, так и по молекулярному составу.) Компонент, определяющий структуру стенок, — пептидогликановый, или муреиновый, слой. Он построен из неразветвленных полисахаридных цепочек, которые переплетены олигопептидными цепочками. Поскольку весь муреиновый слой соответствует одной- единственной гигантской молекуле, его называют также муреиновым мешком. Он может за счет локальных включений новых компонентов увеличиваться и таким образом принимать участие в росте клетки, не переставая выполнять свои опорную и защитную функции. Биосинтез пептидогликана блокируется пенициллином, поэтому данный антибиотик убивает бактериальные клетки, но не клетки эукариот — у них пептидогликаны не встречаются.
Рис. 2.98. Примеры клеточных стенок бактерий (по U. J. Jürgens): А—схема строения клеточной стенки у грамположительной бактерии Bacillus. Цитоплазматическая мембрана (клеточная мембрана) покрыта многослойным пептидогликаном; в плоскости клеточной стенки проходят тейхоевые кислоты (линейные полимеры из остатков фосфоглицерина или рибитолфосфата, которые ковалентно связаны с пептидогликаном; напротив, липотейхоевые кислоты зафиксированы в цитоплазматической мембране и тянутся перпендикулярно плоскости стенки. Весь комплекс клеточной стенки покрыт S-слоем (англ, surface — поверхность), с которым через боковые радикалы ковалентно связаны направленные наружу цепочки полисахаридов капсулы; В—соответствующая схема для грамотрицательной бактерии, например, Escherichia coli. Пептидогликан здесь однослойный. Наружная мембрана (англ, outer membrane) жестко связана с ним липопротеиновыми единицами (показаны серым). Она пронизана тримерными поринами и содержит белок А наружной мембраны (outermembrane protein А, показан точками) в качестве интегрального структурного белка. Наружный одинарный слой наружной мембраны состоит из липополисахаридов с ориентированными внутрь жирными кислотами липида А и изогнутыми наружу полисахаридными цепочками (О-антигены), а также из амфиполярных единиц ЕСА (enterobacterialcommon antigen — общий энтеробактериальный антиген) с более длинными вытянутыми полисахаридными цепочками. Кроме того, здесь закреплены полисахариды капсулы (К-антиген)
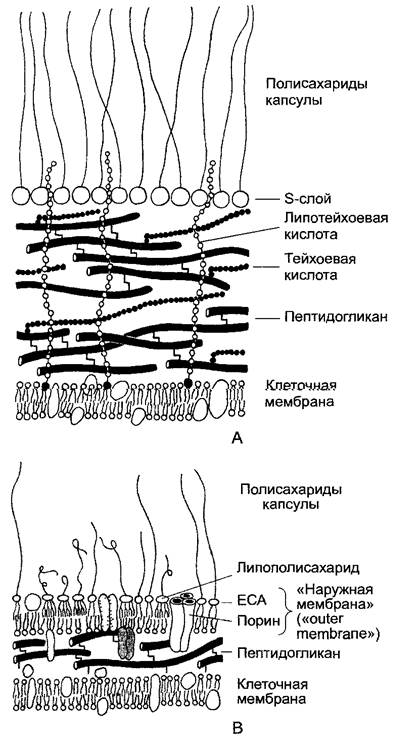
Грамположительные и грамотрицательные бактерии по строению клеточной стенки четко отличаются друг от друга. (Окраска по Граму — генцианвиолет + йод — может у грамотрицательных бактерий вымываться этанолом, а у грамположительных не может.) У грамположительных бактерий пептидогликановый слой грубый, он
состоит из многих муреиновых пластов. У грамотрицательных бактерий и цианобактерий, напротив, муреиновый мешок сравнительно тонкий. Однако здесь кроме мешка имеется еще характерный слой, который по внешнему виду его среза в электронном микроскопе называют наружная мембрана(outer membrane). По своему молекулярному строению он сходен с биомембраной в том отношении, что представляет собой двойной липидный слой, внутренний слой липидов которого состоит преимущественно из фосфолипидов. Внешний слой липидов, напротив, построен из липополисахаридов — комплексных полимеров с остатками жирных кислот в качестве липофильного компонента и характерными олиго- и полисахаридными цепочками, которые отходят наружу. Вместе они образуют гидрофильный защитный слой вокруг прокариотических клеток, через который не могут проникать липофильные молекулы. Гидрофильные частицы, напротив, проходят. В липидном двойном слое наружной мембраны находятся трехмерные комплексы трансмембранного белка (порина), который образует гидрофильные поры диаметром примерно 1 нм. (Порины наружной мембраны оболочки митохондрий и пластид имеют соответствующую функцию, однако по аминокислотным последовательностям они не показывают близкого родства с бактериальными поринами, которые в свою очередь очень разнородны.) Наружная мембрана — это слой клеточной стенки, а не настоящая биомембрана. В противоположность настоящим биомембранам она может также образовываться de novo; например, она снова регенерирует после полной утраты стенки. Наружная мембрана нигде не граничит с цитоплазмой и не обладает также транслокаторами для специфического или вообще активного транспорта. Пространство между клеточной и наружной мембранами называют периплазматическое пространство.
Большинство прокариот может при неблагоприятных условиях образовывать споры с особенно прочными и непроницаемыми стенками.