БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
4. МОРФОЛОГИЯ И АНАТОМИЯ ЛИСТОСТЕБЕЛЬНЫХ РАСТЕНИЙ
4.3. Листовые органы. Строение и метаморфозы
Многообразие строения листьев крайне велико. Оно варьирует от едва заметных чешуек до метровых многократно перистых вай древовидных папоротников и листьев пальм, от зеленых игольчатых хвоинок и разной формы листовых пластинок фотоассимилирующих листьев до ярко окрашенных лепестков цветков, от листовых колючек до изощренных кувшинчатых ловчих листьев насекомоядных растений. Но во всех этих случаях речь идет о видоизменениях единственного типа органа — филлома. Его первоначальные функции — фотосинтез и транспирация. Их выполняет собственно лист, который можно охарактеризовать как ассимилятор и транспиратор.
В морфологическом отношении филлом представляет собой наиболее развитую форму листа. Остальные формы по сравнению с ним выглядят упрошенными благодаря редукции.
4.3.1. Типичный лист
4.3.1.1. Расчленение и симметрия
Сложившееся в процессе филогенеза морфологическое строение типичного листа с нерасчлененной листовой пластинкой показано на рис. 4.56.
Нижняя часть листа (Unterblatt)1 включает листовое основание и, если имеются, прилистники(Stipulae, ед. число Stipula, от лат. stipula — соломинка). Листовое основание часто выглядит как расширение основания черешка, однако у однодольных оно обычно столь широко, что целиком охватывает стебель в узле. В таких случаях листовое основание обычно удлиняется в листовое влагалище, как это можно наблюдать у большинства злаков.
1 Понятия Unterblatt и Оberblatt используют только в немецкоязычной литературе; они не имеют точных соответствий в других языках. — Примеч. ред.
Рис. 4.56. Лист ивы козьей (Salix саргеа) как пример типичного филлома
Расширенным основанием лист прикрепляется к стеблю с боков основание фланкировано прилистниками непосредственно над ним располагается пазушная почка (стрелка) Основание листа и прилистники составляют нижнюю зону листа. Верхняя зона состоит из черешка и листовой пластинки (12х). У перистых листьев (ср рис 4 21) черешок в виде рахиса продолжается в область листовой пластинки и несет парные листочки и верхушечный листочек (у непарноперистосложных листьев)

Такое листовое влагалище функционирует как опорный орган для соломины, тонкого стебля злаков. Утолщенными листовыми влагалищами являются также запасающие чешуи луковиц. В других случаях из удлиненных, охватывающих друг друга листовых влагалищ образуется ложный стебель (рис. 4.57, А). В типичном виде он свойствен банану, но его можно наблюдать на ранних стадиях развития и у местных европейских однодольных (например, у чемерицы, Veratrum; или злака Molinia). Истинные, несущие соцветия стебли прорастают позже внутри трубчатого ложного стебля.
Рис. 4.57, Ложные стебли и ложная кора
А—ложный стебель лука репчатого (Allium сера) образованный полыми цилиндрическими листовыми влагалищами (0 6х) В, С—стеблеобъемлющее основание листа типично для многих однодольных, истинный стебель часто невидим примеры — популярные комнатные растения Aloe spmosissima и Dracaena mar- gmata (0 6x) D E— ложная кора, представленная листовыми основаниями у хвойных D—Thuja onentahs (накрест-супротивное листорасположение 2 1х) Е—вертикальный побег ели Picea abies с очередным листорасположением, слева олиственный, справа после опадения хвои удлиненные листовые основания вплотную прилегают друг к другу — в противоположность пихте где округлые листовые основания не покрывают сплошь стебель (ср рис 4 16, Е)
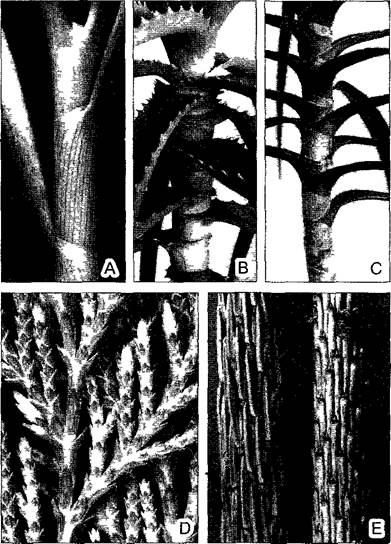
У многих хвойных деревьев листовое основание не стеблеобъемлющее, но оно удлиняется вдоль побега и срастается со стеблем. Если к тому же эти основания соприкасаются друг с другом боковыми сторонами, образуя плотный рельеф на стебле, то говорят о ложной коре (рис. 4.57, D, Е).
Прилистники у многих растений вообще не образуются или существуют недолго и рано опадают (как у лещины и граба, где они играют роль почечных чешуй), однако могут быть и весьма крупными и даже брать на себя функции типичного листа (рис 4.58) Нередко прилистники превращены в колючки, как у робинии или белой акации.
Верхняя часть листа включает черешок и листовую пластинку. Черешок выносит на некоторое расстояние от стебля листовую пластинку, которая собственно и служит главным органом ассимиляции и транспирации. За счет ростовых или тургорных движений он может изменять ее положение для оптимальной экспозиции к солнечному свету. Как несущий орган черешок часто обладает более или менее округлым поперечным сечением, напоминая этим осевые органы. Но бывает, что расширенные и уплощенные черешки перенимают функции листовой пластинки (рис 4.59). В таких случаях говорят о филлодиях. Если черешок отсутствует, листья называются сидячими
Многообразие филломов проявляется прежде всего в разнообразии формы листовой пластинки (от лат lamina — пластинка, см рис 4 2). Наряду с формой крайне вариабельна и величина листьев — от миллиметровых до почти 20-метровых, как перистые листья пальмы Raphia /armífera.
Рис. 4.58. Листовое основание и прилистники
А—листовидные прилистники у гравигата Geum urbanum (1,6х) У некоторых растений прилистники полностью берут на себя функции листьев, например, у чины прилистниколистной (ср. рис 44.69); В—подмаренник Galhum mollugo как будто имеет мутовчатые листья, но стебель четырехгранный и только из пазух двух противостоящих листьев вырастают пазушные побеги, только эти органы — настоящие листовые пластинки, а остальные — сходные с ними листовидные прилистники В качестве альтернативы можно было бы рассматривать листья как сидячие, то есть не имеющие черешков супротивные пальчатые листья (2,1х), С—листовое основание без прилистников у ореха грецкого (Juglans regia, 1,6х), D, Е—одревесневшие, превратившиеся в колючки прилистники белой акации Robinia (D—0,3х; Е—1,7х)

Рис. 4.59. Филлодии у Acacia heterophyíía (по J. Reinke).
После дважды перистых первых листьев образовались перистые листья с крылатыми черешками и, наконец, у следующих листьев без пластинок — уплощенные черешки филлодии перенимают функции листьев
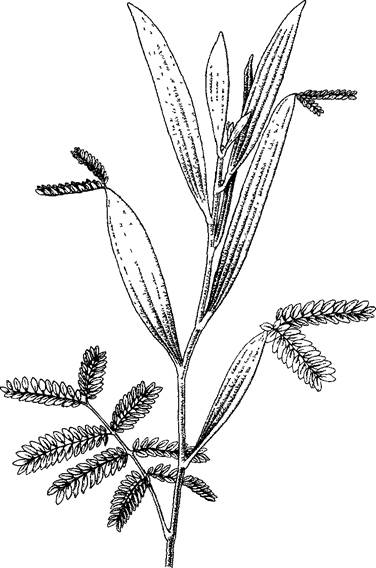
Многие народные названия растении даны по форме листьев (селезеночник, печеночница, лапчатка, стрелолист, манжетка и т.п.). В определителях приведены диагностически важные различия растении и их названия. Морфологически особенно интересны перистые листья. У них черешок продолжается в рахис, на котором располагается несколько пар боковых листочков и (чаще всего) непарный конечный листочек. У вай папоротников особенно часто встречается многократная перистость, когда перышки первого порядка расчленены на перышки второго порядка, а те в свою очередь — на перышки третьего порядка. Если продольный рост рахиса однажды перистых листьев заторможен, то все перышки кажется прикрепленными на конце черешка; такие листья называют пальчатыми. Особые формы вроде шитовидных, вальковатых или мечевидных листьев, а также трубчатые ловчие листья хищных растений мы еще рассмотрим при обсуждении развития диета.
Типичный лист билатерально симметричен; его медианная плоскость совпадает с направлением черешка и соответственно рахиса, в ней располагается средняя жилка. Отклонения от билатеральной симметрии редки и поэтому особенно заметны (косой лист у бегонии). У типичных листьев чаще всего также отчетливо выражена дорсовентральность: их обращенная к стеблю (по крайней мере исходно) ада- ксиальная, или морфологически верхняя, сторона отличается многими особенностями от абаксиалъной, нижней, стороны. Различия касаются, например, числа устьиц (большинство листьев гипостома- тичны: у них более 90 % устьиц расположены в нижней эпидерме), опушенности, накопления пигментов в вакуолях клеток эпидермы и, наконец, анатомии: палисадная паренхима преимущественно адаксиальна, а губчатая — абаксиальна. В листовых проводящих пучках — жилках, или нервах листа (оба эти обиходные названия в сущности ошибочны), ксилема расположена сверху, а флоэма снизу. Дорсовентральность листьев — следствие полярности стеблей, на которых листья расположены сбоку, так что поверхность листа ориентирована поперек градиента полярности.
Уже при рассматривании листа невооруженным глазом заметен характер их жилкования, т.е. узор из проводящих пучков в листовой пластинке (рис. 4.60). Более мощные пучки (главные жилки, англ, major veins) снабжают листья водой и обеспечивают обратный транспорт продуктов фотосинтеза из листьев. Они окружены обкладкой, которая контактирует с системой тканей мезофилла и контролирует обмен веществ между проводящими пучками и мезофиллом. Такие обкладки пучков иногда достигают эпидермы и выполняют также опорную функцию. Главные жилки часто выступают с нижней стороны листа в виде ребер жесткости, укрепляющих пластинку. Крайний случай отмечен у гигантских плавающих листьев Victoria amazónica (рис. 4.61). Но основная функция жилок листа — снабжение водой и минеральными солями особенно активно фотосинтезирующих и транспирирующих клеток мезофилла, а также обеспечение быстрого оттока продуктов фотосинтеза. В проводящих элементах идет конвективный1 поток содержимого; за пределами пучков транспорт веществ ограничен диффузией. Эффективность диффузионного транспорта падает как квадрат преодолеваемого расстояния и фактически недостаточна уже на расстоянии в несколько клеток. Сама вода течет по сосудам в миллион раз легче, чем через живую ткань. Соответственно тонкие жилки, осуществляющие непосредственный контакт с тканью мезофилла, образуют столь густую сеть, что в расположенных между ними ареолах ни одна клетка мезофилла не удалена от ближайшего пучка далее, чем на 7 других клеток. (Обратите внимание термин «ареола» употреблен здесь совсем в другом значении, ср рис 4 7, С). Общая длина проводящих пучков листа бука достигает примерно 30 м.
1 Механизм дальнего транспорта по проводящим тканям не соответствует процессу конвекции в том смысле, как его понимают физики, — Примеч. ред.
Рис. 4.60. Жилкование листьев (узор из проводящих пучков в листовой пластинке)
А, В—дихотомическое (вильчатое) жилкование в листьях папоротников А—Adiantum capillus-venens (3,5х), В—Asplénium nidus (2,4х), С—сочетание параллельного и сетчатого жилкования у маранты (Maranta), листовая пластинка (показана только часть) окрашена так что напоминает облиственный стебель — предположительно мимикрия для того чтобы помешать отложить яйца насекомым-вредителям (1 4х), D—параллельное жилкование у пальмы Sabal umbracufifera (0 7х) Е—сетчатое жилкование у девичьего винограда Parthenocissus tncuspidata (3 5х)

Рис. 4.61. Ребра на нижней стороне плавающего листа Victoria anazonica (= regia); лист в диаметре почти 2 м (фото W. Barthlott)

Эти функциональные задачи могут быть решены по-разному. У однодольных преобладает параллельное жилкование все главные жилки тянутся продольно. Особенно четко выражено такое расположение проводящих пучков в линеиных листьях злаков. В ланцетных листьях большинства других однодольных жилки проходят ровными дугами в соответствии со столь же ровным дуговым краем листа, ти- пичного для однодольных (см. рис. 4.14, А). При параллельном жилковании главные пучки связаны между собой более мелкими поперечными пучками, так что в действительности имеется правильная сеть жилок. (Это можно хорошо видеть невооруженным глазом, например, в листьях Clivia.)
У настоящих двудольных образуется более сложная сеть жилок: сетчатое жилкование. Оно допускает почти любую форму листовой пластинки и особенно ее края. Различия в жилковании отражаются и на расположении устьиц: у однодольных растений они чаще всего ориентированы параллельно, тогда как у магнолиевых и настоящих двудольных — беспорядочно (см. рис. 3.13).
Третий тип жилкования, вильчатое, или веерное, жилкование, имеется у папоротников и у причисляемого к голосеменным гинкго. Здесь более мощные проводящие пучки ветвятся дихотомически и заканчиваются слепо у края листа. Поэтому это жилкование как «открытое» раньше противопоставляли якобы «закрытому» жилкованию однодольных и настоящих двудольных растений (сравните замкнутое и незамкнутое кровообращение у животных). Впрочем, и при сетчатом жилковании тончайшие жилки слепо заканчиваются в мезофилле.
4.3.1.2. Развитие и особые формы листьев
Зачатки листьев, листовые примордии, возникают на конусе нарастания экзогенно как боковые выросты за счет разделения апикальной меристемы (см. рис. 3.3;
3.5).
У папоротников в мелкоклеточной зоне фланговой меристемы сначала возникает двухстороння верхушечная клетка листового зачатка. Из нее развивается линейная краевая меристема, в которой исходная верхушечная клетка уже незаметна. Для большинства листьев папоротников типичен акропластический рост, когда верхушечная часть листа продолжает нарастать, а клетки в основании листа уже дифференцированы. Расчленение листьев папоротников обусловлено фракционированием краевой меристемы вследствие локального прекращения делений ее клеток.
У покрытосеменных только что образованные листовые примордии проявляют ясную тенденцию к расширению своих оснований перпендикулярно оси побега. Так возникает широкое листовое основание, которое может полностью охватывать стебель и приводить к образованию листового влагалища. Гребневидная краевая меристема затем образует листовую пластинку. При этом в противоположность папоротникам здесь преобладает базипластический рост, т.е. активность краевой меристемы сначала угасает на верхушке и в последнюю очередь — в основании листовой пластинки. Перистые листья чаще всего возникают, как и у папоротников, в результате фракционирования краевой меристемы. Главные жилки дифференцируются от основания, тогда как тонкие жилки сначала полностью развиваются в дистальной части пластинки.
Черешки листа образуются путем интеркалярного роста, т.е. благодаря меристеме, находящейся между уже сформированными частями листа. Сходным образом цельнокрайние параллелонервные листовые пластинки однодольных, прежде всего злаков, обязаны своим возникновением базальной интеркалярной меристеме. (Как, впрочем, и медленно отмирающие с верхушек, а у основания продолжающие нарастать более 500 лет листья своеобразного представителя голосеменных Welwitschia; см. рис. 11.211, А.)
Дорсовентральность листовой пластинки выражается в том, что большинство листьев бифациальны, т.е. их верхняя и нижняя стороны выглядят по-разному (от лат. facies — вид, облик; рис. 4.62, A—D). Бывают также эквифациальные листья, особенно у обитателей солнечных мест. Обе стороны таких листьев устроены одинаково: например, они имеют одинаковую плотность устьиц и палисадный мезофилл под абаксиальной эпидермой (рис. 4.62, F, I). Такие листья часто утолщены или игловидны и повернуты ребром к стеблю. Если к этому добавляется суккулентность, то развиваются эквифациальные вальковатые листья, как, например, у очитка (Sedum; см. рис. 4,70, А). Другой способ образования вальковатых листьев состоит в том, что нижняя сторона листа растет интенсивнее верхней, так что последняя в конце концов просто исчезает: унифациальность. Черешки листьев часто приближаются к такой унифациальности, приобретая округлую в поперечном сечении форму, напоминающую стебель. Но и листовые пластинки некоторых однодольных (ситники, некоторые виды лука, например, шнитт-лук) унифациальны и радиально симметричны. Своеобразный и особый случай — листья ирисов (Ins): речь идет о вторично уплощенных унифациальных листьях, причем это уплощение шло не перпендикулярно к оси корневища, а в том же направлении («мечевидные листья)») (рис. 4.62, Е).
Рис. 4.62. Поперечные срезы листьев разных типов (схемы А—Н—по W Troll и W Rauh, I—по Volkens)
Палисадная паренхима показана крапом, нижняя сторона листа — толстой линией, ксилем- ные части проводящих пучков черные А—нормальный бифациальный плоский лист (ср рис 4 64); В—обратно-бифациальный плоский лист (например, черемша, или медвежий лук Allium ursinum), С, D—происхождение унифа- циального вальковатого листа (например, Alliumsativum, Juncus effusus), E—унифациальный мечевидный лист (Iris), F—эквифациальный плоский лист, G—эквифациальный игольчатый лист (см рис. 4 65, А), Н—эквифациальный вальковатый лист (например, Sedum, см рис 4.70, А), I—поперечный срез эквифациального листа пустынного растения Reamuna hirtella, из Tamaricaceae (ЗОх)
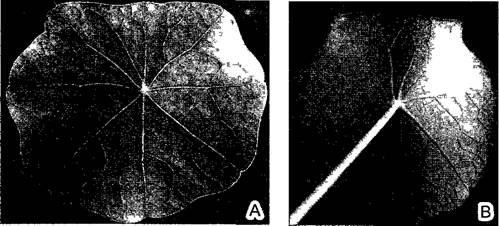
У пельтатных — щитовидных — листьев (от греч. pelte — щит) черешок отходит не от нижнего конца, а почти от центра листовой пластинки (рис. 4.63). Это обусловлено тем, что из-за ярко выраженной базипластии краевая меристема листовой пластинки мощно нарастает непосредственно от места отхождения черешка, причем благодаря его унифациальности правый и левый края пластинки прилегают вплотную друг к другу и срастаются Подобным образом развиваются трубчатые листья некоторых специализированных растений-хищников из семейств Sarracentaceae и Nepenthaceae (бокс 4 4 рис А).
Рис.4.63. Щитовидный лист настурции Tropaeolum majus (0,7х), вид сверху (А) и снизу (В)
У некоторых растений срастаются края не одного и того же листа, а разных листьев одного узла (гамофилия). Это явление довольно редко встречается в вегетативной части растения, но широко распространено в цветках сросшиеся чашелистики и лепестки, а также ценокарпные завязи
4.3.1.3. Анатомия
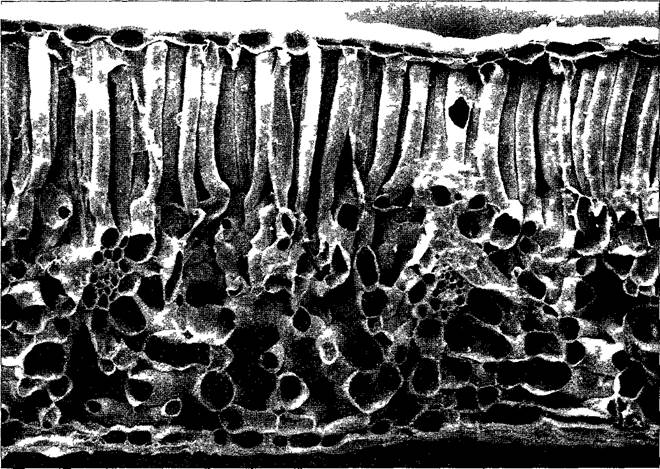
Типичная картина поперечного сечения бифациального листа показана на рис 4.64. Однослойные эпидермы окружают хлоренхиму мезофилла, дифференцированную на палисадную и губчатую паренхиму. Более плотная, одно-, двух- или трехслойная палисадная паренхима содержит около 4/5 всех хлоропластов листа, это основная ассимиляционная ткань. Напротив, очень рыхлую губчатую паренхиму (см рис 3 7, В) можно охарактеризовать как транспирационную ткань. За счет многочисленных, частью очень больших межклетников (они занимают до 90 % объема мезофилла) общая поверхность всех клеток мезофилла почти в 100 раз больше поверхности листа. Система межклетников губчатой паренхимы у гипостоматических листьев облегчает также диффузию СО2 к палисадной паренхиме Клетки эпидермы содержат лейкопласты, иногда с немногочисленными тилакоидами и малым содержанием хлорофилла. Более крупные проводящие пучки окружены эндодермой, которую здесь называют обкладкой пучка. Под ней обычно находится кольцо передаточных клеток, соответствующее перициклу (см 4 4 2 1). Этот клеточный слой, как и сама эндодерма, имеет железистый характер и служит для контролируемого обмена веществ между пучком и мезофиллом. Часто пучки сопровождают и склеренхимные волокна.
Рис. 4.64. Анатомия бифациального листа поперечный срез листа морозника вонючего (Heleborus foetidus, 100х) (СЭМ-фотография Н О Ihlenfeldt)
Под верхней эпидермой палисадная паренхима ниже — рыхлая губчатая паренхима с двумя перерезанными поперек проводящими пучками снизу лист покрыт нижней эпидермой В таких листьях на эпидерму приходится около 12 % объема на проводящие ткани — 5 % и на мезофилл (палисадную и губчатую паренхиму) — 68% около 16% занимают межклетники
Особая структура листа присуща так называемым С4-растениям, фотосинтез которых приспособлен к особенно сильному солнечному освещению в сухих местообитаниях (см 6 5 8) У этих растений окончательная фиксация СO2 происходит в клетках обкладок пучков, которые, соответственно, особенно велики и богаты пластидами («венечный тип» листовых проводящих пучков, см рис 6 81) Пластиды клеток обкладок пучков не образуют гран1, но содержат много ассимиляционного крахмала. Напротив, хлоропласты ассимиляционной паренхимы с гранами, но без крахмала (диморфизм хлоропластов, см, рис. 6.82). Клетки обкладок пучков и мезофилла связаны многочисленными плазмодесмами. Весь комплекс тканей действует как насос для СО22: пластиды клеток обкладок получают достаточно СO2 даже в том случае, если устьичные щели сужены для уменьшения транспирации и поэтому концентрация углекислого газа в межклетниках падает.
1 Только у некоторых С4-растений хлоропласты в клетках обкладок не имеют гран, у большинства растений граны многочисленные и крупные. — Примеч. ред.
2 Это крайне упрощенное и искаженное описание функционирования хлоренхимы С4-растений. — Примеч. ред.
У многих растений расположение тканей в листовой пластинке более или менее сильно отклоняется от показанного на рис. 4.64 (см. рис. 4.62, В—I). Нередко внутри от нижней эпидермы также находится палисадная паренхима (см. рис. 4.71, А). В вертикально стоящих листьях злаков мезофилл гомогенный, он не подразделен на палисадную и губчатую паренхиму, а устьица расположены с одинаковой частотой на обеих сторонах листа. Листья водных растений (например, водяной чумы Еlоёеа) часто бывают толщиной всего в два слоя клеток, а у растений крайне влажных местообитаний встречаются листья даже с одним слоем клеток (Нуmenophyllum).
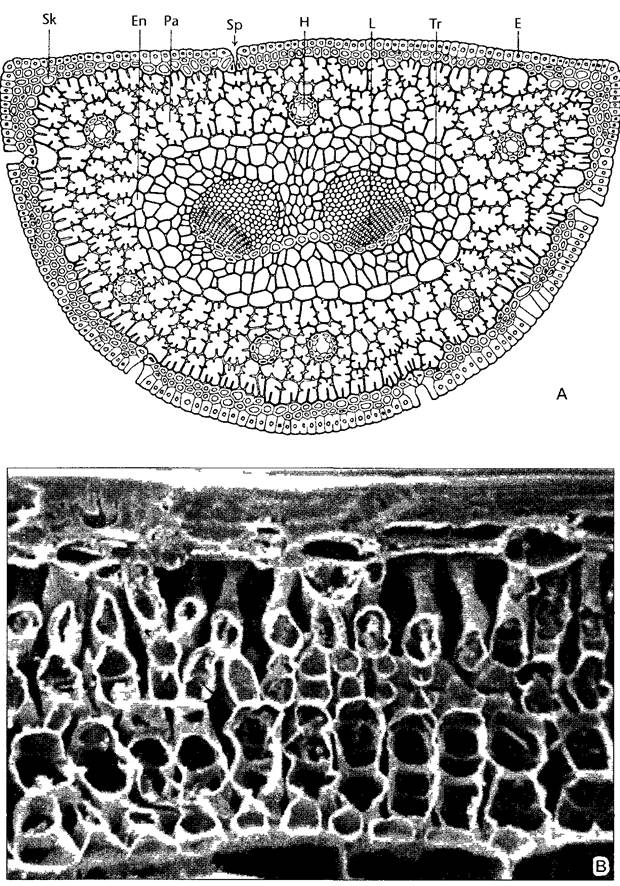
Внутреннее строение эквифациального листа показано на рис. 4.65 на примере игольчатого листа. У листьев этого типа разделение на губчатую и столбчатую паренхиму выражено в общем нечетко, и часто вовсе отсутствует, как и в представленном случае. На поперечном срезе сосновой хвоинки видны клетки мезофилла полигональных очертаний. Поверхность клеток увеличивают гребневидные утолщения стенок1 (дланевидная паренхима). Отсутствие межклетников только кажущееся: дисковидные слои ассимиляционной ткани в одну клетку толщиной, ориентированные перпендикулярно продольной оси хвоинки, отделены друг от друга межклетниками. Между ассимиляционной тканью и эпидермой, клетки которой после мощного утолщения стенок отмирают, находится мертвая, лишифицированная механическая ткань (гиподерма). Устьица, замыкающие клетки которых нуждаются в соединении с живой тканью, погружены до уровня ассимиляционной ткани. В мезофилле хвоинки проходит несколько продольных смоляных ходов. Один или два неразветвленных проводящих пучка игольчатого листа окружены трансфузионной тканью и общей эндодермой. Транспорт веществ между проводящими элементами и мезофиллом осуществляет трансфузионная ткань, образованная паренхимными клетками — к флоэме примыкают типичные клетки Страсбургера с выраженным железистым характером, — а также мертвыми, короткими трахеидами.
1 На самом деле это не утолщения, а инвагинации стенок. — Примеч. ред.
Рис. 4.65. Поперечный (А) и продольный (В) срезы эквифациального игольчатого листа сосны черной (Pinus nigra, 40х) (А—по R von Wettstein, В—СЭМ-фотография, 285х)
На продольном срезе видны межклетники Sk — склеренхимная гиподерма, Еn—эндодерма, Ра—ассимиляционная паренхима, Sp—устьице, Н—смоляной канал, L— проводящий пучок, ксилема сверху, Тг—трансфузионная ткань, Е—эпидерма
4.3.2. Листовая серия
Как было показано выше (рис. 4.5; 4.6), развитие филломов у покрытосеменных меняется в онтогенезе одного и того же растения — то есть при одном и том же генотипе — в очень широких пределах, соответствующих ниже приведенной (максимальной) схеме.
Сравнение разных листьев (см. листовую серию на рис. 4.66) показывает, что упрощенные их формы, такие как низовые листья, почечные чешуи, верховые листья и филломы цветка возникают в результате торможения развития верхней части листа и усиленного развития нижней части. Листовая серия — впечатляющее проявление способности органа к изменениям путем изменения пропорций.
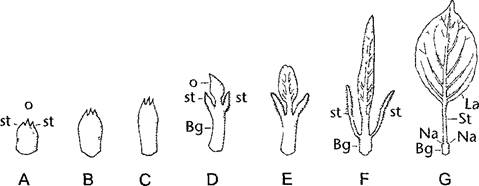
Рис. 4,66. Развитие верхней части листа (Oberblatt) при переходе от почечных чешуй (А—С) к типичному листу (G) у яблони Malus bacata (по W.Troil)
D—Е — переходные листья, F—типичный лист накануне распускания (А—F — почти в натуральную величину; G—0,2х); st—прилистники; Вg—основание листа; Nа— рубцы оставшиеся после опадения прилистников; St—черешок листа; Lа—листовая пластинка, о—верхняя часть листа
Если семядоли в семени служат «мясистыми» резервуарами запасных веществ, то во время прорастания они обычно остаются внутри растрескивающейся семенной кожуры и поэтому или оказываются на поверхности почвы или остаются в земле — подземное прорастание (дуб, конский каштан, горох, фасоль кроваво-красная и др.). Чаще встречается надземное прорастание, когда семядоли благодаря удлинению гипокотиля выносятся на свет и зеленеют (ель, бук, клен, горчица, подсолнечник, фасоль обыкновенная и др.).
1 Предлистья — самые первые листья на боковом побеге, они никогда не следуют за почечными чешуями. — Примеч. ред.
Насколько сильно может измениться листовая серия, показывает пример известного декоративного растения Streptocarpus hydridus. Оно образует только две семядоли, поначалу одинаковые. Позднее одна из них очень сильно увеличивается в единственный, долго живущий «лист», в пазухе которого затем развивается соцветие.
И наоборот, листовая серия может дополнительно усложняться, когда, например, различаются молодые и старые типичные листья, как у плюща. Об анизофилии говорят, если соседние листья, порой даже одного узла, оказываются разного размера/мощности вследствие дорсовентральности плагиотропного побега (рис 4.67) Под гетерофилией следует понимать такое явление, когда в зависимости от внешних или внутренних условий развиваются совершенно иначе устроенные листья с разными функциями (рис. 4.68, особые соотношения у плавающего папоротника Salvinia см. рис 11.158) Общеизвестны различные формы листьев молодых ползучих и отстоящих от субстрата цветоносных побегов плюща.
Рис. 4.67. Анизофиллия (по W Troll, В — по К Goebel)
А—индуцированная анизофиллия у клена остролистного, или платановидного (Acer pfatanoides) листья двух соседних мутовок косо растущей ветви, стрелка вектор силы тяжести (0,25х), В—облигатная анизофиллия у селагинеллы Дугласа (Selagmetta doudlasu) каждый узел несет один крупный вентральный и один маленький дорсальный лист (5х) (Листья у всех селагинелл не супротивные, а очередные — Примеч. ред.)
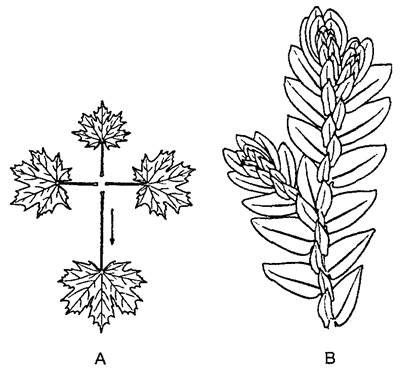
Рис. 4.68. Модификационная гетерофиллия у лютика водяного {Ranunculus aquatihs) (по W Troll)
А—цветущая, симподиально нарастающая ветвь с плавающими и тонко рассеченными подводными листьями (w), В—лист переходной формы
Отдельные филломы листовой серии отличаются не только по форме и функциям, но и по продолжительности жизни. Особенно короткоживущими обычно являются семядоли и листочки околоцветника. Сильно редуцированные и чаще всего быстро опадающие верховые листья называют брактеями (от лат. bractеа — листочек). Типичные листья живут дольше, но у летне-зеленых растений (лиственные деревья, а из хвойных лиственница) опадают в конце каждого вегетационного сезона. Перед листопадом прежде всего азотосодержащие соединения преобразуются и транспортируются из листьев. В ходе этих драматических изменений хлоропласты становятся геронтопластами1, которые окрашиваются в желтый цвет сохраняющимися каротиноидами, обычно образующими сложные эфиры с жирными кислотами. Листья/хвоя вечнозеленых растений и кустарников функционируют по несколько лет (сосна — 2 года; пихта — 5 — 6 лет; араукария — до 15 лет). Листья опадают вследствие образования особой отделительной ткани (см. рис. 7.61).
1 В ботанике такие пластиды принято называть хромопластами. — Примеч. ред.
4.3.3. Видоизменения листьев
4.3.3.1. Метаморфозы
Уже упоминалось, что листья, как и побеги, могут превратиться в колючки (см. 4.1.1) и усики (см. 4,2,6). Примеры таких метаморфозов показаны на рис. 4.69 и 4.7. Часто листья функционируют как запасающие органы, и наряду со стеблевыми суккулентами есть также и листовые. Крупные водоносные клетки располагаются либо субэпидермально, либо в глубине листа (например, у Lithops, — эти растения называют «живые камни» южноафриканских пустынь; см. 15.2.6).
У некоторых растений клетки самого мезофилла увеличиваются за счет необычно крупных вакуолей. В этом случае речь идет о морфологической корреляции с особым приспособлением фотосинтеза к солнечному, жаркому и сухому местообитанию. Такое приспособление известно под названием САМ-метаболизм (см. 6.5.9). Crassulaceae — «толстолистные» растения (лат. crassus — толстый), к ним относят среди прочих молодило (Sempervivum) и очиток (Sedum, рис. 4.70). САМ-растения имеются не только в семействе Сrassulaсеае. но и в 27 других семействах, даже среди суккулентных папоротников Они фиксируют СO2 ночью при открытых устьицах в промежуточной форме. Синтезируемая в этих условиях яблочная кислота накапливается в крупных вакуолях клеток мезофилла. Днем устьица из-за опасности слишком больших потерь влаги закрываются, но теперь высвобождаемый из яблочной кислоты СO2 может быть окончательно ассимилирован с помощью энергии света.
Рис. 4.69. Усики (А, В — по H.Schenck; С — по F. Noll): А—усики перистого листа гороха посевного (Pisum sativum); В—листовой усик чины Lathyrus aphaca, С—побеговый усик с дисками-прицепками девичьего винограда (Par- thenocissus tricuspidata) (все 0,6 1); s—стебель; n—прилистники; г—листочки сложного листа, превращенные в усики; а—пазушный цветонос; b—рахис сложного листа
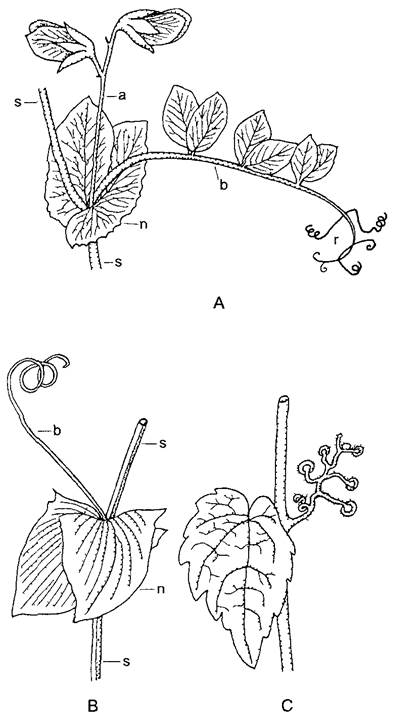
Рис. 4.70. Листовые суккуленты (0,75х): А—очиток белый (Sedum album)- В—молодило Sempervivum schnittspahmi, розетка с очередными листьями

Метаморфоз листьев наблюдается прежде всего у растений, приспособившихся к необычным условиям обитания или к особому образу жизни. В таких случаях соответствующие изменения претерпевают чаще всего не только листья, но и все растение, — возникает приспособительный синдром. Три синдрома, касающиеся и строения листа, коротко представлены в двух следующих разделах и в боксе 4.4 (экологическая морфология).
4.3.3.2. Ксероморфные листья
Для растений сухих (аридных) областей (степи, пустыни) или местообитаний (скалы, песчаные почвы; от греч. xeros — сухой) критическое значение приобретает обеспеченность водой. Водообеспечение невозможно стабилизировать путем усиленного поглощения воды, поэтому остается лишь ограничить водоотдачу, т е. транспирацию, если вообще не отказаться от активной жизни в периоды засухи. Мы уже видели, что многие ксерофиты преобразуют свои листья в колючки или редуцируют их до маленьких чешуек, а фотосинтезируют уплощенными стеблями, которые не содержат транспирируюшей ткани1 и обладают сравнительно малой поверхностью относительно объема Кутикулярная транспирация крайне ограниченна; во многих случаях образуется система накопления воды (стеблевые суккуленты; см рис. 4.35).
1 Это неверное утверждение, так как любая фотосинтезирующая ткань, будь она в листе или в стебле, транспирирует. — Примеч, ред.
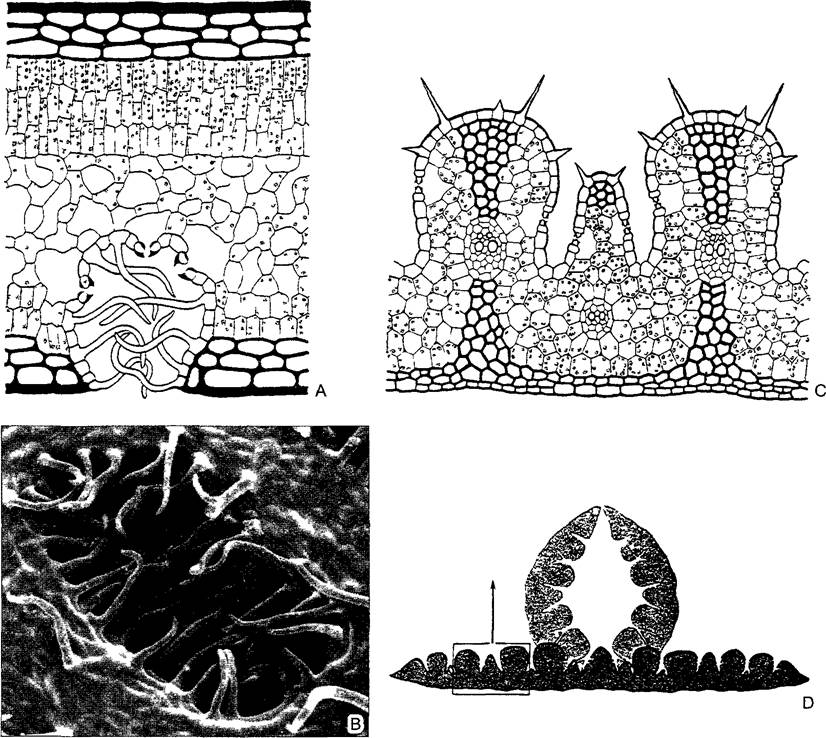
Многие ксерофиты, впрочем, сохраняют ассимилирующие листья. Эти листья «ксероморфны», т.е. устроены так, что их транспирация мала и/или может быть снижена при необходимости. Действительно, листья ксерофитов отчетливо отличаются от листьев мезофитов и гигрофитов, населяющих места с достаточным или обильным увлажнением. В то время как листья гигрофитов (впрочем, и «теневые» листья, например, у бука (см. рис. 7.73), тонкие и чаще всего голые, с непогруженными, а часто даже приподнятыми над поверхностью эпидермы устьицами, ксероморфные листья обычно кожистые, жесткие (жестколистные растения — лавр, мирт, оливковое дерево), с погруженными устьицами (рис. 4.71). Листья, скручивающиеся в засуху, вообще изолируют свои устьица от контакта с окружающей средой. Кроме того, потерю воды можно ограничить с помощью очень толстой кутикулы и мощного воскового налета, а часто дополнительно — и густого опушения, благодаря которому у самой поверхности листа создается слой неподвижного, более влажного воздуха. Жесткость листьев ксерофитов, исключающая их увядание, обусловлена развитием склеренхимных волокон или одиночных звездчатых склереид. Ксероморфные листья часто эквифациальны.
Рис. 4.71. Анатомия ксероморфных листьев (А, С, D—по О Stocker, В—СЭМ-фотография W Barthlott)
А—олеандр с многослойной эпидермой (черная) трехслойной палисадной паренхимой и глубоко погруженными устьицами, в углублениях (криптах) движению воздуха препятствуют волоски (80х), В—крипта вид снаружи (170х), CD—листья ковыля Stipa capillata эпистоматичны, т е устьица расположены только на верхней стороне. При засухе листья скручиваются на верхнюю сторону, укрывая тем самым устьица от наружного воздуха, при хорошем водоснабжении листовая пластинка разворачивается и становится плоской Мезофилл не дифференцирован на палисадную и губчатую паренхиму, как и у других злаков (С—80х, D—10х)
Эквифациальные игольчатые листья (см. рис. 4.65) также явно ксероморфны. Это приспособление к сильной абсорбции инфракрасной части солнечного излучения и связанному с ней разогреву темной хвои на прямом солнечном свету. Из-за сравнительно низкой пропуск ной способности трахеид древесины хвойных в игольчатых листьях такое прогревание уже в обычных условиях может приводить к дефициту воды. При длительных морозах, когда поглощение воды из промерзшей почвы прекращается, водный баланс в прогретых на свету хвоинках легко становится критическим — «морозная сухость».
Сокращением транспирации водный обмен удается стабилизировать, но острой остается проблема перегрева паренхимы листа/стебля (рис 4.72, 4 73, 4 74) (Транспирация создает мощный охлаждающим эффект из-за сравнительно высоких энергетических затрат на испарение воды — около 41 кДж/моль.) У многих растений чрезмерному нагреванию листовых пластинок препятствует их поворот ребром к направлению света. Известны не дающие тени австралийские эвкалипты, чьи серповидные листья отвесно свисают с ветвей. На стволы охлаждающе действуют выступающие ребра и корка с глубокими трещинами (см. рис. 4.54, А; 4.55, G-I).
Рис. 4.72. Кувшинчатые листья Dischidia major (средний разрезан вдоль) с придаточным корнем, который врастает в полость листа через отверстие кувшинчика (0,8х) (фотография WBarthlott)
4.3.3.3. Листья эпифитов
В противоположность лазящим растениям, всегда укореняющимся в почве, эпифиты с самого начала поселяются в кронах деревьев, чтобы занять место под солнцем. Деревья служат им только поддерживающим субстратом; их могут заменить скалы, крыши домов, даже телефонные провода. Таким образом, большинство эпифитов — не паразиты. Правда, при мощном развитии они могут «задушить» свою живую опору. Лишь немногие эпифиты, например, омела, являются паразитами.
У растений, обитающих на бедных питательными веществами, особенно азотом, субстратах (например, на верховых болотах) встречается удивительная пищевая специализация эти растения способны жить как фотоавтотрофы, но дополнительно оснащены приспособлениями для ловли и удержания мелких животных, прежде всего насекомых. Эти хищные, или насекомоядные, растения используют добычу как дополнительный источник азота, переваривая ее внеклеточно (см 9 1.2). Их листья преобразованы различным, часто весьма причудливым образом для ловли животных.
Сравнительно просто действуют клейкие ловушки росянок (Drosera). На их листьях расположены снабженные тяжем трахеид эмер- генны-«щупальца» (ср рис. 3 31) Их железистые головки выделяют блестящие капельки клейкого секрета, который привлекает мелких животных. Насекомое прилипает к железке и, пытаясь освободиться, касается соседних железок и все крепче увязает. Раздражение «щупалец» вызывает их сгибание — насекомое оказывается прижатым к поверхности листа. Здесь его тело (за исключением хитина) химически разрушается выделяемыми специальными железками секретами и в растворенной форме всасывается.
Венерина мухоловка Dionaea (см рис. 8 28, Е, F) способна мгновенно закрыть ловушку-капкан, сложив пополам лист. Это движение осуществляется с помощью шарнирного сочленения на главной жилке и регулируется осмотическими процессами. Такое движение происходит, как только насекомое, садясь на открытый лист, касается одного из чувствительных волосков (см. 8.3.2.4) Сложенные пополам края листовой пластинки усажены зубцами наподобие капкана и могут удерживать даже таких сильных насекомых, как оса или шмель, которые перевариваются выделяющимися ферментами.
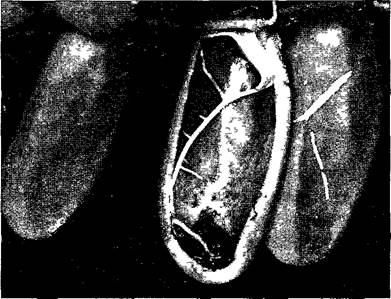
У Nepenthes, Cephalotus, Sarracenia и Darlingtonia скользкими ловушками служат урно- образные, имеющие форму кувшина или мешочка листья. Кувшинчики Nepenthes (рис. А) содержат водянистую кислую пищеварительную жидкость, выделяемую железками, расположенными на стенках. Привлеченные животные соскальзывают по гладкому краю кувшина, «вымощенному» восковыми пластинками, тонут в его содержимом и перевариваются ферментами.
Рис. А. Ловчий кувшинчик у Nepenthes (С — фото W Barthlott)
Кувшинчик образован мешковидной листовой пластинкой В кувшинчике высотой несколько сантиметров (А, 0,3х) скапливается несколько миллилитров пищеварительного секрета выделяемого особыми щитовидными железками (D, 260х) Добыча, чаще всего насекомые, садится на ярко окрашенный край кувшинчика, скользкий из-за эпикутикулярных восковых пластиночек (В, 1,2х), под завернутым краем внутри находятся нектарные железки Насекомое, пытаясь добраться до нектара, случайно соскальзывает в ловушку Крышка кувшинчика во время его развития закрыта и препятствует попаданию внутрь дождевой воды, позднее она остается постоянно открытой Черешок листа может функционировать как усик (стрелка на А), подвешивая тяжелый кувшинчик на ветвях Удлиненная и расширенная базальная часть листа (G—на А) берет на себя функции листовой пластинки
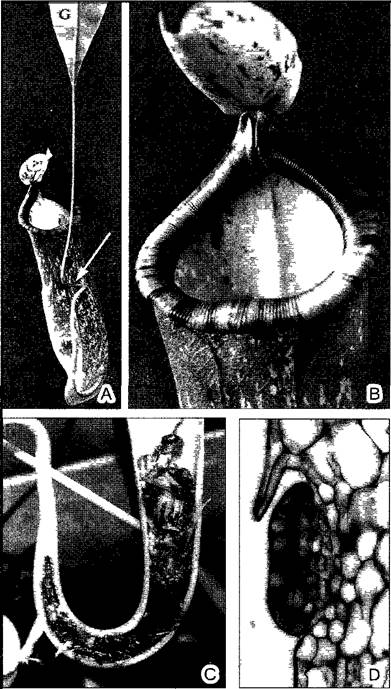
Живущие в стоячих водоемах погруженные растения видов Utricularia несут на своих рассеченных листьях маленькие зеленые пузырьки (рис. В), наполненные водой: это заглатывающие (засасывающие) ловушки. «Рот» такой ловушки сначала водонепроницаемо закрыт клапаном. Если мелкое водяное животное натолкнется на один из волосков на внешней стороне клапана, действующих по типу рычага, то клапан откроется и с потоком воды засосет жертву (это прежде всего мелкие рачки, личинки насекомых, коловратки и простейшие) в пузырек величиной около 2 мм. Процесс заглатывания происходит путем ослабления напряжения в стенках пузырька, первоначально эластично вдавленных. Затем клапан возвращается в исходное положение и снова запирает ловушку.
Рис. В. Ловчий пузырек пузырчатки (Utricularia) (В—СЭМ-фотография W Barthiott)
А—ловчий пузырек U vulgaris в продольном разрезе (10х), В—сложный аппарат чувствительных антенн U sandersonii (100х)
Для более крупных листостебельных эпифитов обеспечение водой и минеральными солями представляет существенную проблему. Благоприятные условия для жизни эти растения находят только в регионах с обильными дождями и постоянно высокой влажностью воздуха, главным образом в дождевых тропических лесах. При этом эпифиты демонстрируют тем более выраженное ксероморфное строение, чем выше сухость воздуха.
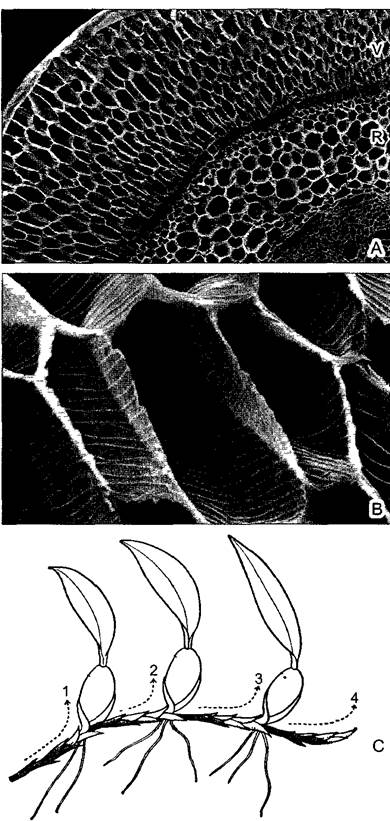
В свободно свисающих, нередко зеленых воздушных корнях обычно развивается особая поглощающая воду ткань — веламен (рис. 4.73, А, В). У других эпифитов воздушные корни, растущие вверх, формируют густое ветвистое сплетение, в котором скапливаются влага и гумус. Папоротник костенец гнездовидный (Asplenium nidus) образует густые розетки крупных вай, в их воронковидных пространствах постепенно накапливается гумус. У папоротника оленьи рога (Рlatycerium) через определенные промежутки времени образуются особые щитовидные листья, прилегающие к субстрату, за ними могут скапливаться вода и гумус — пример гетерофиллии (см. рис. 11.162). Еще дальше зашло преобразование части листьев у Dischidiа из семейства Asclepiadасеае: за счет очень сильного плоскостного роста пластинок при одновременном торможении ее краевого роста отдельные листья становятся мешковидными с узким отверстием (см. рис. 4 72). В них живут колонии муравьев, натаскивающие туда почву, а влага накапливается благодаря конденсации водяного пара. В полость каждого такого листа врастает придаточный корень, развивающийся на том же узле. Таким образом, растение «высаживает» само себя в созданные им же «цветочные горшочки».
Рис. 4.73. Приспособления эпифитных орхидей тропических дождевых лесов (А, В—СЭМ- фотография S. Porembski и W Barthlott; С—по W. Troll): А—поперечный срез воздушного корня Dertdrobium nobile, между веламеном (лат. — покров) из заполняющихся водой при дожде мертвых клеток и корой находится однослойная экзодерма с пропускными клетками; паренхима коры отделена от стелы (на фото справа внизу) однослойной эндодермой (60х); В—клетки веламена со спиральными утолщениями стенок (460х) (Аналоги гиалиновых клеток в листьях торфяного мха, сфагнума, ср. рис. 11.120, G.) С—Coelogyne sp., симподиальная система четырех генераций побегов, завершающихся клубнями (1—4, 0,2х). (Правильнее называть эти структуры туберидиями. — Примеч ред.) V—веламен; R—кора
В других случаях образуются стеблевые клубни, запасающие воду во время дождей (см. рис. 4.73, С). Широко распространены и другие специфические приспособления для эффективного улавливания осадков. У бромелиевых корни представляют собой короткие жесткие органы прикрепления; у некоторых видов, как у часто свисающих с телефонных проводов представителей рода Tillandsia, корни могут совсем отсутствовать. Воду эти эпифиты получают исключительно через поглощающие волоски на листьях (рис. 4.74). Часто у этих растений плотно прилегающие друг к другу базальные части розеточных листьев образуют цистерны, где собирается дождевая вода.
Рис. 4.74. Щитовидные всасывающие волоски (чешуйки) эпифитных бромелиевых (Bromeliaceae, 170х) (СЭМ-фотография W. Barthlott): А—Tillandsia rauhir, В— Acanthostachys. Мертвые клетки щитков волосков при дожде заполняются водой, которая по живым клеткам ножки волоска передается в лист
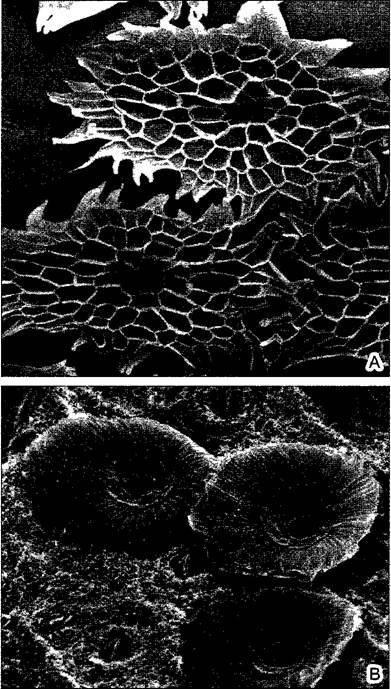
В общем, созданные эпифитами мини-водоемы дают пристанище микроорганизмам, водным улитками, насекомым и даже некоторым лягушкам. На Ямайке в цистернах бромелиевых живет даже пресноводный краб Metopaulias depressus.
4.4. Корни
Корневая система обычно выполняет две функции; заякоривание растения в почве и поглощение из нее воды и минеральных питательных веществ.
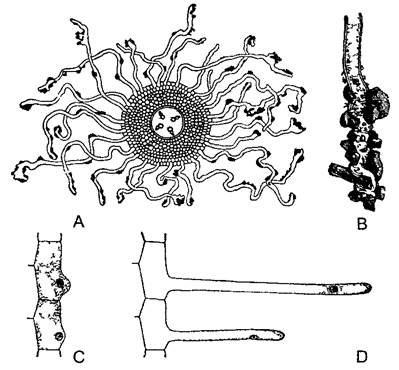
Второй задаче часто соответствует чрезвычайное увеличение всасывающей поверхности корня. Многие клетки самого наружного, не имеющего кутикулы слоя (ризодермы =эпидермы корня1) вырастают в длинные (от нескольких миллиметров до нескольких сантиметров) корневые волоски (рис. 4.75). Корневые волоски, как и весь корень в целом, растут верхушкой и поэтому способны внедряться между частицами почвы. Они живут недолго (3—9 дней), зона корневых волосков растущего корня достигает всего 1—2 см в длину. Подсчитано, что, несмотря на это, взрослое растение ржи имеет свыше 10 млрд корневых волосков, общая длина которых достигает 10 000 км, а общая поверхность — 400 м2. Это примерно в 50 раз больше, чем вдвое более тяжелая надземная часть растения, включая листья
1 Ризодерма не гомологична эпидерме. — Примеч. ред.
Рис. 4.75. Корневые волоски (А—по Frank В—по F Noil, С, D—по Rothert)
А—поперечный срез через зону всасывания корня с тетрархным центральным цилиндром корневые волоски с частицами почвы (10х) В—окончание корневого волоска при большем увеличении (50х), С, D—ризодерма на продольном срезе, начальные и последующие стадии образования корневого волоска (обратите внимание на положение клеточного ядра) (50х)
Наряду с заякориванием и поглощением воды и минеральных веществ корни часто выполняют и другие функции В них, к примеру, синтезируются такие важные вещества, как растительные гормоны (цитокинины, гибберел лины, см 7 6 2, 7 6 3) Обычно они функционируют и как запасающие органы (см бокс 4 5)
4.4.1. Корневые системы
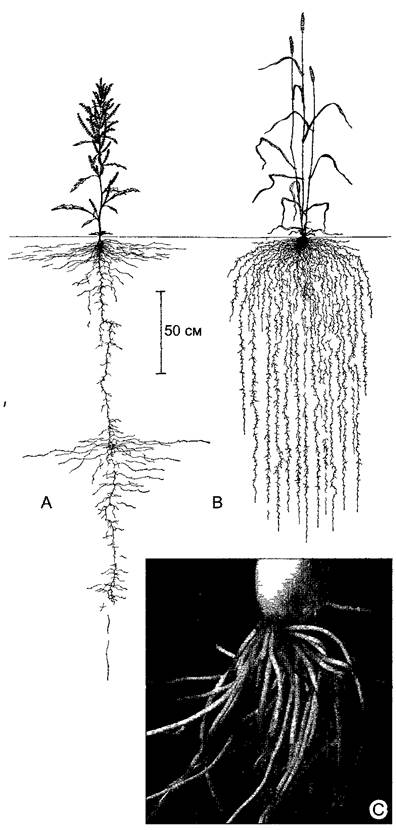
Корневые системы растений, как и все осевые системы, устроены весьма по-разному в зависимости от предпочитаемого местообитания. У молодых или быстро распространяющихся с помощью ползучих побегов растений корневая система часто больше, чем побеговая (рис 4 76). И напротив, особенно слабо развита корневая система у многих кактусов, растущих в сухих, жарких биотопах, где почва почти всегда (по крайней мере в течение дня) совершенно сухая. В зависимости от вертикального распространения корней в почвенном профиле можно выделить две группы растений — с глубокой корневой системой и с поверхностной. Самые глубокие корневые системы у растений из мест, где поверхностный слой почвы сухой, но в глубине проходят водоносные жилы грунтовых вод (например, Welwitschia; см. рис. 11.211, А; стержневые корни тамарисков достигают предположительно глубины 30 м, а у дерева Prosopis juliflora из североамериканских пустынь даже более 50 м). У деревьев размер корневой системы в целом определяется размером кроны: самая наружная зона корневой системы слегка выходит за пределы площади проекции кроны.
Рис. 4.76. Аллоризия и гоморизия (А, В—по L Kutschera)
А—Rumex crispus настоящее двудольное растение образует гетерогенную (аллоризную) корневую систему первичный корень которой внедряется в почву более чем на 3 м В—вторичная гоморизная корневая система пшеницы с характерным для большинства злаков мочковатым типом корневой системы, С—ярко выраженная гоморизия имеется у луковичных растений (например, у молодого лука Аllium fistufosum) многочисленные, довольно мясистые корни все одинаковой толщины и почти не ветвятся (0,7х)
По особенностям развития и дефинитивного строения можно выделить два типа корневой системы: гетерогенную (аллоризия) и гомогенную (гоморизия).
Гетерогенные корневые системы. У многих растений зародышевой корешок вырастает в главный, или первичный, корень, образуя вертикально внедряющийся в почву стержневой корень. От него отходят вторичные, боковые корни первого порядка, растущие в почве косо или горизонтально, которые в свою очередь ветвятся (боковые корни второго, третьего и последующих порядков). Боковые корни высоких порядков растут независимо от вектора силы тяжести и поэтому могут пронизывать почву во всех направлениях. Такую иерархически построенную корневую систему называют аллоризной, или гетерогенной (от греч. alios — различный, rhiza — корень; см. рис. 4.76, А).
Большинство деревьев аллоризны, некоторые сохраняют исходную стержневую корневую систему и позже: пихта, сосна, дуб. У других древесных пород (например, у лиственницы, березы, липы) в дополнение к исходному главному корню образуются все новые мощные корни, идущие в почве косо, так что под основанием ствола возникает полусферическая система корней, примерно одинаковых по мощности, которую называют кистевидной. Деревья с поверхностной корневой системой (к ним относят ель и рябину) обладают системой мощных, растущих горизонтально почти под поверхностью почвы вторичных корней, от которых вертикально вниз отходят существенно более слабые и короткие корни.
У некоторых видов деревьев, например, у ели и вяза полевого, в загущенных популяциях корни срастаются с корнями других растений. Над толстыми, словно канаты, тяжами корней, тянущимися иногда на многие метры, несколько деревьев могут создать действительно «крепко спаянный» коллектив.
Гомогенные корневые системы. Гоморизные системы состоят полностью или преимущественно из корней одного порядка ветвления, сходного строения, не ветвящихся или слабо ветвящихся (греч. homos — одинаковый, похожий; см. рис. 4.76, В, С). Гомогенной корневой системой обладают все папоротникообразные. Как споровые растения они не образуют семян, так что у них нет зародышевого корешка; их осевая система вообще не имеет корневого полюса и закладывается униполярно, поэтому у Pteridophyta все корни придаточные (первичная гоморизия). Кроме того, расположение их корней четко коррелирует с положением листьев: под основанием каждого листа возникают один или несколько корней, у крупных древовидных папоротников их бывает более 100.
Первичная гоморизия типична для папоротникообразных. У семенных растений с их биполярными зародышами имеется не первичная, а вторичная гоморизия. У однодольных она возникает вследствие того, что от нижнего узла побега отрастают многочисленные корни одного порядка ветвления. Они функционально дополняют слабо развитую систему первичного корня, которая, как и базальная часть побега, не претерпевает первичного утолщения (см. рис. 11.220). Эти корни часто выполняют дополнительно опорную функцию (например, у кукурузы; см. также бокс 4.5, рис. В). Они возникают, таким образом, в ходе регенерационного процесса и представляют пример придаточных корней (см. рис. 7.21; 7.24). И у настоящих двудольных часто бывают придаточные корни, например, на усах и у всех корневищных растений. Крайне своеобразный случай представляют мангровые растения (Rhizophora; бокс 4.5, рис. В), а также тропический фикус Ficus bengalensis, чья раскидистая крона покрывает площадь до 2 га (диаметр 170 м), опираясь на сотни придаточных колонновидных воздушных корней.
Строгая приуроченность корней к листьям, характерная для папоротникообразных, у семенных растений встречается редко. Тем не менее и у них придаточные корни часто развиваются на узлах побега. Однако из этого правила есть много исключений1 (ср. бокс 4.5, рис. А).
1 Число таких примеров ничтожно мало по сравнению с числом видов, образующих узловые придаточные корни. — Примеч, ред,
4.4.2. Анатомия корня
4.4.2.1. Первичное строение
Схема поперечного среза корня на рис. 4.77 показывает радиальную симметрию на стадии первичного строения. К ризодерме изнутри примыкает более жесткий и долговечный, часто слабо опробковевающий слой клеток — гиподерма; в оболочках ее клеток часто образуются пояски Каспари, благодаря чему она становится экзодермой. Этот слой окружает мощно развитую паренхиму первичной коры, которая внутри ограничена эндодермой (см. 3.2.2.3). Последняя представляет собой морфологическую и физиологическую границу центрального цилиндра, в котором объединены механические и проводящие элементы. Центральное положение этой прочной ткани в менее прочной оболочке обеспечивает гибкость при высокой прочности на растяжение (кабельное строение, см. 3.2.3) и соответствует выполняемой корнем функции заякоривания растения в субстрате.
Рис. 4.77. Расположение тканей на поперечном срезе корня.
Центральный цилиндр окружен перициклом, ксилема показана черным, флоэма — крапом, между ними полоски паренхимы. Здесь ксилема трехлучевая, соответственно центральный цилиндр триархный. (Более правильно считать гиподерму экзодермой. — Примеч. ред.)
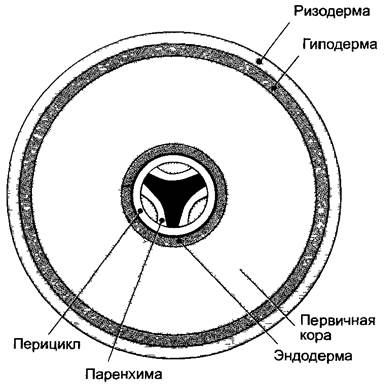
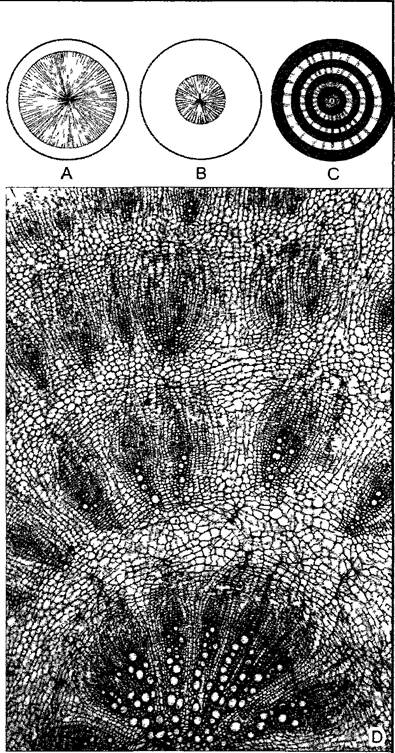
Самый наружный слой центрального цилиндра, перицикл, состоит из тонкостенных, богатых цитоплазмой клеток, которые длительное время сохраняют способность к делению. Поэтому этот клеточный слой, в котором, как и в ризо-, экзо- и эндодерме, мало межклетников, называют также перикамбием1 (см. 4.4.2.2). Середина центрального цилиндра, устроенная как актиностела2, обычно занята ксилемой, которая в виде двух или большего числа радиальных гребней доходит до пери цикла. По числу таких ксилемных нолюсовразличают 2-, 3- ... полиархные центральные цилиндры3. У папоротникообразных, магнолиид и настоящих двудольных преобладают 2 —4-архные центральные цилиндры (рис. 4.78, А; 4.79, А), в то время как однодольные часто обладают полиархным центральным цилиндром (рис. 4.78, В). В промежутках между гребнями ксилемы располагаются тяжи флоэмы. Ксилема и флоэма отделены друг от друга слоями паренхимы, которая по бокам от гребней ксилемы доходит до пери цикла.
1 Этот термин широко применяли до середины XX в., а затем он вышел из употребления. — Примеч. ред.
2 На самом деле сравнивать следует с радиальным проводящим пучком, но не с актиностелой. — Примеч. ред.
3 Понятие архности правильно относить к ксилеме радиального пучка, а не к центральному цилиндру. — Примеч. ред
Рис. 4.78. Центральный цилиндр (А—по D von Denffer, В—СМ-фотография Н Fafk)
А—поперечный срез тетрархного проводящего пучка корня лютика едкого (Ranunculus асег) (160х), В—поперечный срез додекархного центрального цилиндра корня Iris germaniса, на котором легко различимы те же ткани, что показаны на фрагменте А, за исключением участков тонкостенной флоэмы, расположенных непосредственно под перициклом между 12 ксилемными полюсами. Стрелка — пропускная клетка, эндодерма (как на А) в третичном состоянии (120х), Еп—эндодерма с пропускными клетками D, X—ксилема с сосудами, Ph—флоэма с ситовидными трубками S и показанными темным клетками-слутницами Ре—перицикл
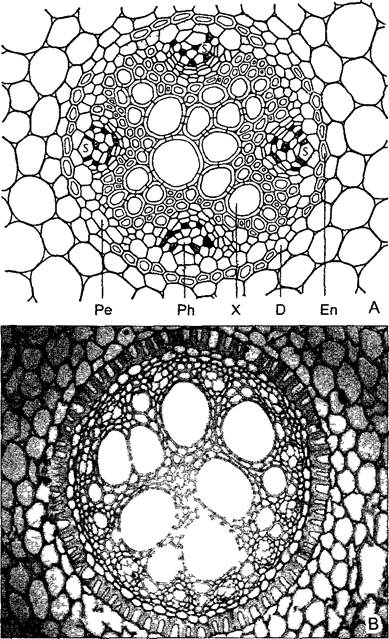
Рис. 4.79. Эндогенное возникновение боковых корней (А—по О Stocker): А—поперечный срез корня настоящего двудольного; над ксилемным полюсом центрального цилиндра из делящихся клеток перицикла (слева вверху) образуется конус нарастания будущего бокового корня, который позднее (вверху справа) прорастает наружу сквозь ткань коры (120х); В—поперечный срез корня Vicia faba с пентархным центральным цилиндром и растущим боковым корнем; меристематические клетки выглядят более темными, чем сильно ва- куолизированные клетки постоянных тканей (75х); en—эндодерма; pz—перицикп, с—камбий в паренхиме (ра), х—ксилема; р—флоэма
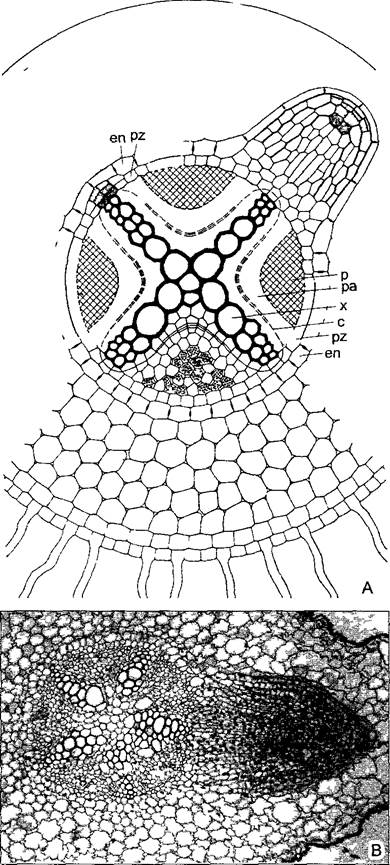
При развитии центрального цилиндра его ткани в противоположность тканям стебля дифференцируются снаружи внутрь1, поэтому протофлоэма и протоксилема располагаются непосредственно под перициклом, а самые крупные сосуды мета ксилемы — в центре. Иногда метаксилема в самом центре корня не развивается. В таких случаях и в корнях настоящих двудольных в центре центрального цилиндра находится паренхимная сердцевина, у однодольных сердцевина обычная. В особенно мощных корнях высоких однодольных растений центральный цилиндр содержит и склеренхиму.
1 Флоэма в стебле также дифференцируется снаружи внутрь — Примеч. ред.
В гипокотиле, пограничной зоне между корнем и стеблем, актиностела корня акропетально переходит в эвстелу или атактостелу стебля (бокс 4 3) Строение этой переходной зоны различно у разных покрытосеменных.
Обычно от корня вверх центральный цилиндр разделяется на отдельные сектора проводящих тканей в каждом с одним ксилемным полюсом и двумя половинками соседних тяжей флоэмы, примыкающими к ксилеме с боков. Эти сектора выше смещены на периферию осевого органа, здесь между ними расположена паренхима — сердцевинные лучи, а в центре — сердцевина. Ксилема каждого выделившегося таким образом проводящего пучка разворачивается так, что участки протоксилемы, лежавшие в центральном цилиндре корня на периферии, теперь оказываются обращенными внутрь, к сердцевине, а метаксилема, соответственно, — наружу.
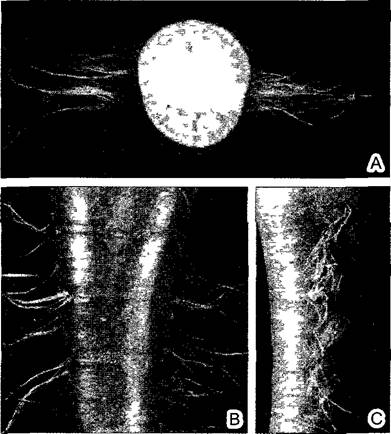
Поскольку корень не несет листьев, у него нет характерной для побегов метамерии — продольного расчленения на узлы и междоузлия. За покрытым корневым чехликом (калиптрой) (см. 3.1.1.2) и, таким образом, субапикальным конусом нарастания следует (за областью покоящегося центра) зона клеточных делений1 и далее — зона растяжения длиной 3—10 мм. Максимальная частота клеточных делений в образующейся первичной коре (периблеме) приходится на область рядом с конусом нарастания, в возникающем центральном цилиндре (плероме), отступя от него, а в молодой ризодерме (дерматоген, или эпиблема) — в наиболее удаленной от инициалей области. Много клеточных делений происходит также в зоне растяжения. К ней примыкает зона корневых волосков и, наконец, область образования боковых корней — зона ветвления. Образованием зоны корневых волосков завершается первичный рост корня. Продольный рост корня здесь заканчивается (рис. 4.80), в противном случае, протискиваясь между частицами почвы, он обрывал бы отходящие вбок корневые волоски. Таким образом, корни растут только на самом конце (см. рис. 7.3).
Рис. 4.80. Ризостихи у редьки: А—поперечный срез (1,2х), В, С—внешний вид, 0,8х).
Каждая из двух ортостих корня (они указывают на диархный центральный цилиндр) в действительности двойная, так как у крестоцветных, или капустных (к которым относится редька) снаружи от каждого ксилемного полюса центрального цилиндра образуются две тесно сближенные ортостихи Диаметр главного корня, образующего корнеплод, в 100 раз больше диаметра боковых корней
1 Следует уточнить: покоящийся центр и зона клеточных делений составляют конус нарастания корня, а не располагаются позади него. — Примеч. ред.
Зона растяжения корня значительно короче, чем у развивающегося в воздухе стебля, где она часто достигает нескольких сантиметров. Это связано со значительно большим сопротивлением, которое приходится преодолевать растущим в земле верхушкам корней: соответственно и у воздушных корней зона растяжения почти в десять раз длиннее, чем у обычных корней.
Бокс 4.5. Метаморфозы корня
Известны многочисленные приспособления корней к выполнению особых функций. Уже функция заякоривания растения в особых условиях может потребовать видоизменения корней. Известные примеры — придаточные цепляющиеся корни лазящих растений (рис. А) и эпифитов. Ходульные корни мангровых растений способствуют закреплению их в подвижном грунте приливной зоны тропических морских побережий (рис. В). В принципе сходную роль играют — правда в совсем других условиях существования — и адвентивные корни высоких злаков. Досковидные корни образуются при усиленном вторичном утолщении верхней стороны корней, растущих горизонтально непосредственно под поверхностью почвы. У некоторых высоких тропических деревьев они образуют вокруг ствола метровой высоты контрфорсы (цв. табл. 15.1, Е, т. 4).
Своеобразна функция втягивающих корней, заглубляющих подземные побеги — корневища, клубни или луковицы (рис. С). Сокращение этих корней основано на том, что стенки продольно вытянутых клеток первичной коры имеют продольно ориентированные фибриллы целлюлозы, так что на повышение тургора они отвечают укорочением (при одновременном утолщении).
Немало растений образуют особые запасающие корни (рис. D); у многих корнеплодов запасающие ткани также хотя бы частично принадлежат корню (рис. Е). Аномальное вторичное утолшение1 приводит к возникновению толстых, но слабо разветвленных участков корня; в некоторых случаях даже образуются шаровидные корневые клубни, не имеющие боковых корней. В качестве запасных веществ встречаются преимущественно ди-, олиго- и полисахариды (сахароза, крахмал, инулин) 2.
1 Подавляющее большинство запасающих корней обладают либо вторичным утолщением нормального типа (двудольные), либо первичным утолщением (однодольные). — Примеч. ред.
2 Корни некоторых растений запасают воду. — Примеч. ред.
Корневые колючки — короткие, полностью одревесневшие, остроконечные боковые корни на придаточных воздушных корнях. У некоторых пальм они защищают основание ствола.
Воздушные корни, как уже говорилось, часто выполняют функцию поддержания по- беговой системы над субстратом. Воздушными корнями поглощают воду эпифиты, которые не могут добраться до почвенной влаги (хотя в некоторых случаях делают это с помощью листьев). Такие корни имеют особую наружную ткань (веламен, ср. рис. 4.73, А, В). Он развивается из протодермы путем периклинальных клеточных делений. Веламен состоит из многочисленных, рано отмирающих крупных клеток со спиральными утолщениями стенок и отверстиями в стенках. Подобно водяным клеткам в листьях торфяного мха сфагнума эти пустые клетки заполняются дождевой водой, которую веламен впитывает и удерживает, словно губка.
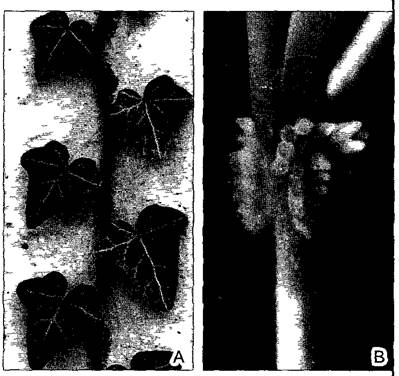
Рис. А—Придаточные скопляющиеся корни: А—у плюща цепляющиеся корни служат не для поглощения воды и минеральных веществ, а исключительно для закрепления на субстрате (здесь — на бетоне; 0,7х). Двухрядное расположение листьев типично для ювенильных форм; В—у кампсиса укореняющегося (Саmpsis radicans) цепляющиеся корни образуются только в узлах (2,6х)
В постоянно переувлажненных почвах мощные корневые системы испытывают дефицит кислорода из-за малой его растворимости в воде. Поэтому многие деревья и кустарники тропических заболоченных лесов и мангровы образуют растущие вверх (отрицательно геотропные) дыхательные корни — пневматофоры, которые поднимаются над поверхностью почвы (воды) так, что система межклетников в тканях коры контактирует с воздухом Особая форма — коленчатые корни, которые сначала растут вверх, но достигнув поверхности почвы, изгибаются и снова уходят вниз, под землю Благодаря одностороннему утолщению (как у досковид- ных корней) на таких изгибах часто образуются торчащие в воздух вздутия — корневые шишки.
Рис. В. Ходульные корни (фото А—D Lupmtz, В—W Barthiott)
А—Rhizophora mucronata, одно из настоящих двудольных растений мангровых зарослей на заливаемом морском пляже (острова Тонга, Юго-Западная Полинезия), В— относящийся к однодольным западноафриканский панданус, или винтовая пальма (Pandanus candelabrum)

Рис. С. Втягивающие корни у аронника пятнистого (Arum maculatum) (А — по Rimbach)
А—заглубление клубня за счет сокращения корня I—прорастание, II—начало, III—конец второго года, IV—взрослое растение, клубень на глубине 10 см от поверхности почвы (0,4х), В—клубень и втягивающие корни, наружные ткани которых не участвуют в сокращении корня и пассивно собираются в поперечные складки (1,8х)1
1 Укорочение корня обусловлено как раз активным сокращением кольцевых зон периферийных тканей корня — Примеч. ред.
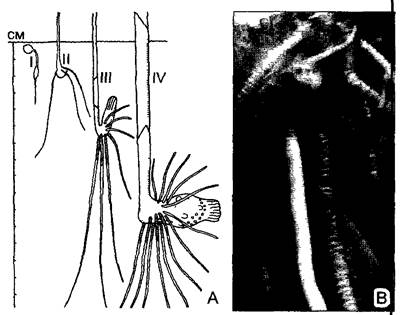
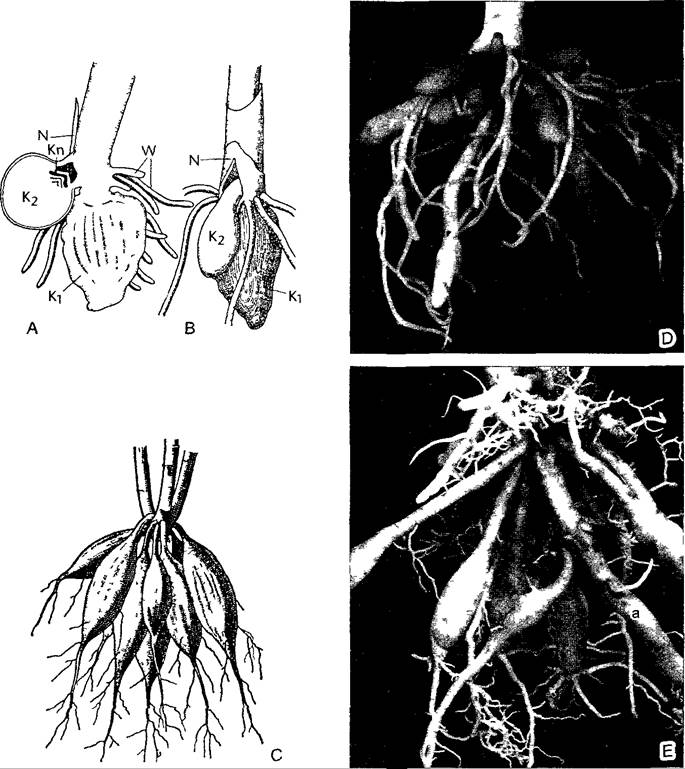
Рис. D. Корни как запасающие органы (А, В—по R von Wettstein, С—по Weber)
А, В—корневые клубни Orchis militans (0,7х), К1—прошлогодний клубень, из которого вырос цветонос текущего года, в пазухе самого нижнего чешуевидного низового листа N на пазушном побеге развивается новый корневой клубень К2, W—нормальные придаточные корни, Кn—почка пазушного побега следующего вегетационного периода, С—придаточные запасающие корни георгины (0,15х), D—придаточные корневые клубни гоморизной корневой системы чистяка весеннего (Ranunculus ficaria), клубни легко отрываются у основания и затем вырастают в новое растение (2х), Е—менее выраженные, чем у георгины, корневые клубни лилейника, или красоднева (Hemerocafhs), но и здесь боковые корни образуются только в дистальной, не запасающей части корня, а — подземный столон (0 5х)
Уже было упомянуто, что корни эпифитных орхидей берут на себя даже функции листьев (см рис 4 4) К сильным морфологическим изменениям корней приводят паразитический и симбиотический образ жизни (см 9 2 9 3).
Полупаразиты — это зеленые растения, которые сами еще способны к фотосинтезу, но воду и питательные соли получают от растения-хозяина, в ксилему которого внедряются с помощью корневых гаусториев. К этой категории относятся, к примеру, очанка, погремок, марьянник и мытник из норичниковых, а также вечнозеленая омела Она прорастает как паразитический эпифит на ветвях определенных деревьев. Затем ее корневая система распространяется под лубом ветви хозяина, а от корней в заболонь хозяина внедряются гаустории, через характерные короткие сосуды подсоединяющиеся к водопроводяшей системе ветви (см. рис. 4.39, В).
Полными паразитами считают растения, не образующие хлоропластов и питающиеся полностью за счет хозяина. Петров крест (Lathraea) через корневые гаустории потребляет пасоку из ксилемы корней древесных хозяев. Виды рода Огоbanche, напротив, присасываются к флоэме корней своей жертвы. Гаустории этих желтоватых, красноватых или лиловых паразитов внедряются сбоку в корень хозяина и так интенсивно поглощают флоэмный сок, что это приводит к отмиранию дистального участка корня. Поэтому кажется, что паразит сидит на конце корня хозяина.
Рис. Е. Анатомия корнеплодов (поперечные срезы) (А—С — по W Franke)
А—в «древесинных» корнеплодах мощно развита главным образом ксилема, которая, однако, состоит преимущественно из древесинной паренхимы; пример — редька, В — в «лубяных» корнеплодах, наоборот, запасающей тканью становится вторичная флоэма; пример — морковь, С—в корнеплодах свеклы (культурные формы Beta vulgaris: сахарная, кормовая, столовая) образуются чередующиеся кольца из ксилемы (светлые) с флоэмой1 и паренхимы (темные); это происходит в результате аномального вторичного утолщения с повторным образованием добавочных камбиев в коре1. Как это выглядит в световом микроскопе, показано на D (внизу исходный центральный цилиндр; 48х).
1 В корнеплоде свеклы образуются отдельные коллатеральные пучки, а не кольца ксилемы с флоэмой. — Примеч. ред.
2 Добавочные камбии образуются не в коре, а в пролиферирующем перицикле. — Примеч. ред.
При симбиозе с азотфиксирующими бактериями образуются корневые клубеньки — локальные разрастания тканей коры (см. 9 2.1). В особых вакуолях увеличенных полиплоидных паренхимных клеток прокариотические симбионты живут в форме «бактероидов».
Гораздо шире распространен называемый микоризой симбиоз с почвенными грибами (см. 9.2.3). Растение использует чрезвычайно мощную всасывающую способность грибных гиф прежде всего для снабжения минеральными солями. Характерно, что корни с микоризой вообще не образуют корневых волосков.
4.4.2.2. Боковые корни
В отличие от боковых побегов боковые корни образуются эндогенно, т.е. внутри тела корня, точно на границе между центральным цилиндром и первичной корой (рис. 4.79). При этом клетки перицикла реэмбрионизуются и, делясь пери- и антиклинально, образуют конус нарастания нового корня. Первые боковые корни всегда закладываются позади зоны корневых волосков Таким образом, речь идет о настоящем новообразовании конусов нарастания; фракционирования материнской апикальной меристемы, как в побеге, здесь нет. Придаточные корни также закладываются внутри коры стебля1. Поэтому проводящие ткани боковых корней рано устанавливают связь с проводящей системой материнского органа, сквозь ткани коры которого новый корень прорастает наружу. Место отхождения бокового корня часто бывает окружено выпяченным краем прорванной коры корня или стебля подобно воротнику.
1 Как правило, закладываются в области камбия — Примеч. ред
Эндогенное возникновение боковых корней и субапикальное положение инициалей конуса нарастания корня имеет важные морфологические и анатомические последствия. В обоих случаях поверхность корня образует внутренняя ткань, первоначально окруженная другой тканью. (Ризодерма первичных корней первоначально ведь также покрыта недолговечными клетками корневого чехлика.) С этим, очевидно, связано то, что ризодерма не покрыта кутикулой и не имеет устьиц.
Боковые корни часто располагаются на материнском корне отчетливыми продольными рядами — ризостихами (рис. 4.80), так как новообразование конусов нарастания в перицикле чаще всего происходит против ксилемных полюсов центрального цилиндра. Следовательно, по числу ризостих можно уже по внешнему виду корня сделать вывод об архности центрального цилиндра.
4.4.2.3. Вторичное строение
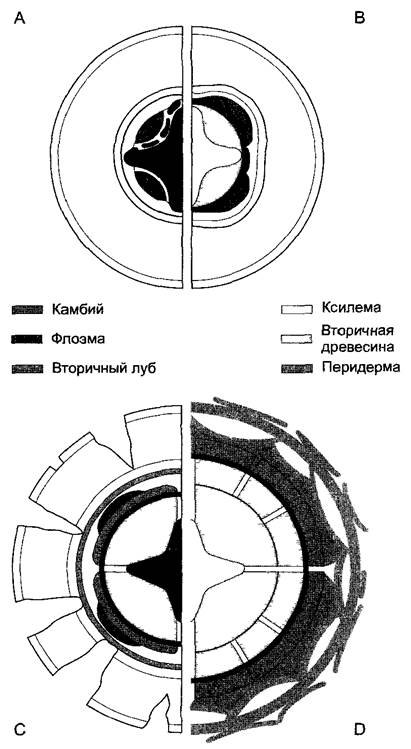
У многолетних древесных растений главный корень подобно стволу мощно утолщается вторично (рис. 4.81). Вначале в вогнутых полосках паренхимы между первичными флоэмой и ксилемой путем реэмбрион изации образуются отдельные участки камбия, которые откладывают внутрь вторичную древесину. Первоначально однослойный перицикл снаружи от ксилемных полюсов становится многослойным, и клетки его самого внутреннего слоя образуют участки камбия. Последние, соединяясь с участками камбия между ксилемой и флоэмой, замыкают камбиальное кольцо. Возникшее кольцо камбия поначалу на поперечных срезах имеет звездчатую форму. Однако вскоре оно округляется благодаря усиленному образованию древесины под тяжами флоэмы. Против ксилемных полюсов закладываются первичные лучи. (Настоящих сердцевинных лучей в корне нет.) Нежная ризодерма чаще всего еще до начала вторичного утолщения отмирает и заменяется гиподермой. Ни она, ни первичная кора корня не участвуют в процессе вторичного утолщения — они разрываются и после отмирания сбрасываются, включая и эндодерму. Поэтому корка, покрывающая сильно утолщенные старые корни, образуется не благодаря формированию перидерм в первичной коре, как в стебле, а за счет перицикла, который сохраняется в виде замкнутого кольца и после начала роста корня в толщину.
Рис. 4.81. Вторичное утолщение корня (поперечные срезы)
А—образование замкнутого камбиального слоя путем реэмбрионизации клеток в паренхиме между ксилемой и флоэмой и над ксилемным полюсом тетрархного центрального цилиндра, В—округление слоя камбия благодаря образованию древесины под тяжами флоэмы, С—начало образования вторичного луба, над ксилемными полюсами возникают лубодревесинные лучи; первичная кора и эндодерма отмирают и разрушаются, в ставшем многослойным перицикле образуется феллоген, откладывающий наружу слои пробки, D—в следующих годичных кольцах закладываются вторичные лубодре веси иные лучи, в лубе благодаря серии феллогенов образуются чешуи корки
Древесина и луб корня гистологически устроены так же, как в стебле. Это относится и к лучам. Поперечный срез корня, который ежегодно рос в толщину, почти не отличается от соответствующего поперечного среза ствола. Только в центре, где сохраняется первичная структура, остаются явные анатомические различия. Но особую переходную зону многолетнего корня в ствол подобно гипокотилю на стадии первичного строения выделить уже не удается.