БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
5. ПРИНЦИПЫ СТРОЕНИЯ ТАЛЛОМНЫХ РАСТЕНИЙ (ТАЛЛОФИТОВ)
Среди листостебельных растений находятся не только самые крупные, но также и самые дифференцированные растения. Цветковые растения могут образовать свыше 70 по-разному устроенных и специализированных к тем или иным функциям типов клеток. Остальные отделы растений не достигают столь сложной организации. Степень дифференцированности, т.е. число типов клеток и тканей, здесь меньше. В этом смысле можно различать растения с низким (более простым) и более высоким (сложным) уровнем организации. Ископаемые находки показывают, что историческое развитие растительного мира, как и мира животных в целом, шло от более простых форм к более сложным. Просто устроенные организмы поэтому обычно противопоставляют как «примитивные» по отношению к сложнее организованным, «продвинутым». Однако с этими понятиями не следует связывать оценки в смысле худшей или лучшей приспособленности. На каждом организационном уровне развились хорошо приспособленные к соответствующим экологическим нишам формы, которые в равной мере прошли испытание отбором.
Все же «примитивные» формы обычно исходные. У них часто сохранились особенности, появившиеся на ранней стадии филетического развития. С другой стороны, растения высокого уровня дифференцированности способны осваивать также биотопы, в которые проще устроенные формы могут внедриться лишь в исключительных случаях.
Например, благодаря тому, что листостебельные растения способны образовывать одревесневшие и кутинизированные или су- беринизированные клеточные стенки, среди них смогли возникнуть крупные наземные формы. Многие из них могут активно выживать и в сухих местообитаниях, так как обладают стабилизированной системой регуляции водного обмена, — это гомогидрические растения. Напротив, большинство мхов, грибов и водорослей «обречены на жизнь» во влажных биотопах или в воде, так как их водный обмен из-за отсутствия снижающих испарение покровных тканей нестабилен в изменчивой влажности воздуха. Они ведут себя как губка, а в сухие периоды впадают в особое состояние, когда все жизненные проявления замирают (латентная жизнь = анабиоз). Такие организмы называют пойкилогидрическими (от греч. роikilos — изменяющийся).
5.1. Одноклеточность и многоклеточность
Очень гетерогенные фототрофные одноклеточные называются протофитами. Такие одноклеточные формы имеются почти во всех классах водорослей.
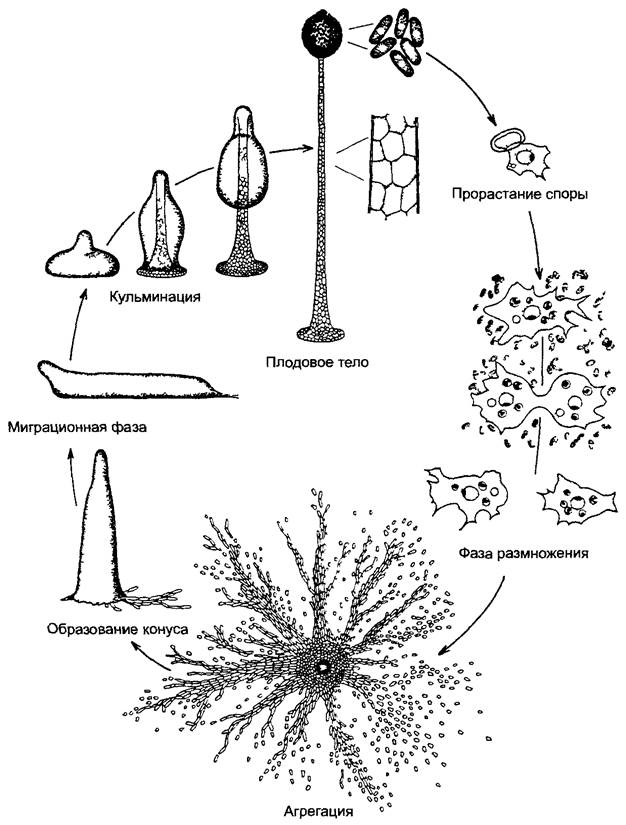
Переход от одноклеточных к многоклеточным характеризуется образованием тканей. Уже у некоторых прокариот имеются межклеточные связи и дифференциация клеток (особенно у цианобактерий, рис. 5 1,11 15). Среди водорослей наряду со слабыми связями клеток, которые удерживаются вместе только за счет общей стенки материнской клетки (ценобии, рис.5.2), имеются и аггрегационные связи, и колонии клеток. Аггрегационные связи образуются путем закономерного взаимного расположения клеток, которые первоначально были независимы и подвижны (см рис. 11 97; 11.98). Крайний случай представляют клеточные слизевики (Acrasiomyceta, например, Dlctyostelшm, рис. 5.3). Аггрегационные связи встречаются, впрочем, и у определенных прокариот, например, у миксобактерий. Колониями клеток называют правильно построенные образования из малого или большого числа клеток, которые происходят от единственной материнской клетки и остаются в постоянной, хотя все еще слабой связи друг с другом. Самый известный пример — Volvox, шаровидным колониям которого свойственны дифференцированность клеток и явный морфогенез (см. рис 11.96).
Рис. 5.1. Нитчатые колонии Nostos, цианобактерии, погруженной в полисахаридный гель, который образуют эти прокариоты
Гетероцисты (стрелки на А, В) выделяются своей величиной. Эти не способные больше к делению клетки с жесткими стенками специализированы на фиксации азота, в то время как фотосинтез идет только в зеленых вегетативных клетках. Гетероцисты непосредственно связаны с соседними клетками через многочисленные плазмодесмообразные каналы. На В и С видно несколько делящихся клеток (разные стадии) (А—170х, В, С—860х)
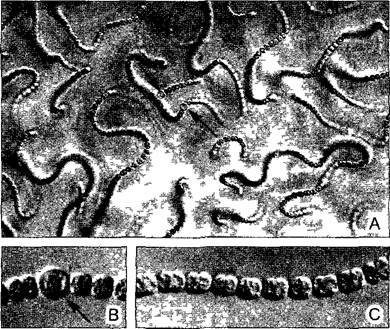
Рис. 5.2. Ценобии (по Е Strasburger)
А—С — образование ценобиев у цианобактерии Gloeocapsa; D—распад ценобия вследствие разрыва самой старой, набухшей клеточной оболочки(500х)
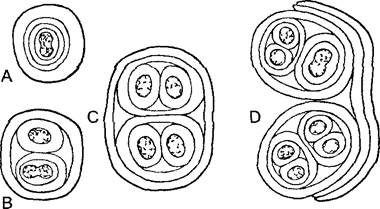
Наконец, макроскопического размера могут достигать многоядерные (полиэнергидные) плазмодии (см. рис. 2.9; 11.16). Плазмодии слизевика Рhysarium polycephalum вырастают до нескольких квадратных сантиметров, содержат свыше миллиарда ядер и ползают, постоянно изменяя форму, по твердым субстратам.
Однако для эволюции крупных морских водорослей, наземных высших грибов и листостебельных растений основополагающим было развитие настоящей многоклеточности.Более 9/10 всех известных видов растений и грибов относятся к многоклеточным Их вегетативные тела состоят из многих или очень многих, преимущественно одноядерных клеток В 1 мм3 ткани находится свыше 1 500 клеток (растительные клетки средних размеров). Из трех этапов, свойственных размножению эукариотических одноклеточных, а именно: деление ядра, деление клетки и расхождение дочерних клеток, — последний остается незавершенным при образовании ценобиев и совсем отсутствует при формировании многоклеточной бластемы — клетки остаются прочно связанными друг с другом Характер бластемы подчеркивает наличие плазмодесм между соседними клетками
Многоклеточность почти без исключений связана с дифференцировкой, которую можно считать прямо-таки типичным признаком многоклеточных Протисты также способны к преобразованию своих клеток, что выражается, например, в образовании покоящихся стадий (цист) или особых форм размножения Однако у них регуляция соответствующей генной активности в общем и целом зависит от внешних факторов, тогда как у типичных многоклеточных наследственно закрепленная дифференцировка происходит благодаря химическим сигнальным веществам, которые образуются внутри бластемы и специфически опознаются компетентными клетками, специфически отвечающими на сигнал Дифференцировка означает специализацию, т е ограничение функций клеток.
Рис. 5.3. Образование упорядоченных агрегатов из амебоидных клеток (по G Gerisch) Цикл развития Dictyostelium discoideum (увеличение справа 100х, слева 8х)
Это имеет очень важное биологическое последствие. Освобожденные от определенных функций соматические клетки больше не служат непосредственно для размножения организма — для этого образуются специальные клетки (половые клетки, споры). Многоклеточный организм освобождает эти клетки и в конце концов погибает. Уже у вольвокса, у которого дочерние колонии образуются из особых клеток, материнская колония отмирает и превращается в труп после того, как заключенные внутри дочерние колонии через разрывы в ней высвобождаются. Смерть по внутренним причинам (физиологическая смерть, в противоположность вызванной внешними причинами катастрофической смерти) и появление трупов жестко коррелирует в живом мире с дифференцировкой и поэтому является неизбежной судьбой всех многоклеточных.
В огромной группе многоклеточных растений, метафитов, можно различить несколько типов организации. В те времена, когда это многообразие типов еще не было четко осознано специалистами, было принято противопоставлять рассмотренные в предыдущей главе листостебельные растения (кормофиты) слоевищным растениям (таллофитам), вегетативные тела которых представлены безлистными осями, не имеющими корней1. Только кормофиты представляют единый тип организации, а в филетическом отношении они также, очевидно, происходят от единого «корня» — группы высокоорганизованных зеленых водорослей. Таллофиты, напротив, развивались независимо друг от друга и многократно. Понятиям таллофиты и протофиты не соответствуют никакие таксоны, это сборные группы, объединяющие многообразные формы. Поэтому и понятие таллом (от греч. thallos — ветвь, лист) можно определить только негативно: оно означает любое многоклеточное или полиэнергид- ное вегетативное тело, которое не имеет типичного для листостебельного растения расчленения. К таллофитам причисляют разнообразные водоросли, грибы и лишайники, а также антоцеротовые и печеночники2. Листостебельные мхи в морфологическом отношении занимают промежуточное положение между талло- и кормофитами.
1 Во многих случаях они имеют не осевое, а пластинчатое тело. — Примеч. ред.
2 Большинство печеночников имеет побеговую организацию. — Примеч. ред.