БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
5. ПРИНЦИПЫ СТРОЕНИЯ ТАЛЛОМНЫХ РАСТЕНИЙ (ТАЛЛОФИТОВ)
5.2. Клеточный и сифональный талломы
Общая филогенетическая тенденция развития все более крупных организмов проявлялась у таллофитов совершенно различным образом.
Кажущийся простейшим путь увеличения единственной одноядерной клетки, т. е. развитие крупноклеточности, — очевидный эволюционный тупик. Растительные организмы такого строения находятся только в одной из групп зеленых водорослей — Dasycladасеае, к которым относят хорошо известную, достигающую более 5 см Аcetabularia. В их сложно устроенных вегетативных телах (см. рис. 11.92) содержится сначала только единственное, необычно крупное «первичное» ядро диаметром более 70 мкм, из которого при дальнейшем развитии возникает свыше 10 000 мелких «вторичных» ядер.
Увеличение одноядерной клетки ограничено ядерно-плазменным отношением. При особенно крупных ядрах отношение поверхности ядра к его объему неблагоприятно. Число ядерных пор, чья плотность на поверхности ядра не может превышать 80 на 1 мкм2, слишком мало относительно объема ядра. Кроме того, слишком велики расстояния для внутриклеточного транспорта генных продуктов в далеко расположенные части плазмы и органеллы. Вторую трудность можно обойти образованием многоядерных гигантских клеток — ценобластов. Многоядерные плазмодиальные крупные клетки широко распространены в различных группах водорослей и у грибов, они представляют сифональный тин организации (от греч. siphon — бурдюк). Так, есть сифональные зеленые водоросли (например, Cauierpa, см. рис. 11.91) и сифональные Xanthophyceae (Botrydium, Vaucheria, см. рис. 11.73). Среди грибов многоядерные талломы особенно свойственны Oomycota, Chytndiomycota и Zygomycota.
Но и этот тип увеличения таллома имеет предел, с одной стороны, потому, что дальнейшее дифференцирование путем модуляции генной активности в многоядерных клетках вряд ли возможно, с другой стороны — сифонобластема, единственная клеточная стенка которой не может быть достаточно толстой, легко уязвима и недостаточно прочна. Характерно, что у превышающего дециметр таллома Cauierpa (см. рис. 11.91, А) противоположные части клеточной стенки укреплены «балками», которые тянутся поперек внутри клетки. У других водорослей (Cladophora, см. рис. 11.90) сифональный таллом поделен на камеры поперечными стенками, но такие клетки все еще многоядерные — сифонокладальная организация.
5.3. Многоклеточные талломы водорослей
5.3.1. Нитчатый таллом
Самой простой формой трихальной организации можно считать нитчатый таллом — ганлонему, равновеликие следующие в ряд друг за другом одноядерные клетки (рис 5.4, А; от греч. trichoos — волосистый, соответственно thrix — волос, nеmа — нить). Известный пример — водоросль спирогира (Spirogiга) (см. рис. 2.87; 11.104, А). У нее все клетки вдоль нити равноценные, дифференцировки нет. В других случаях нитчатый таллом на одном конце прочно соединен с особо устроенной клеткой без хлорофилла — ризоидом, как, например, у зеленой водоросли Ulothrix (см. рис. 11.89, А). В этом случае гаплонема нолярна, т.е. имеется ризоидный конец и верхушка. Полярность нередко усилена тем, что клеточные деления ограничены апикальной клеткой (рис. 5.4, В).
Рис. 5.4. Рост и ветвление нитчатого или соответственно плоского таллома водорослей, отмечены оси веретен деления (по D. von Denffer)
А—нитчатый таллом с интеркалярным ростом; В—рост посредством верхушечной клетки, С—то же с апикально-полярным ветвлением, D—вильчатое деление верхушечной клетки путем делений, периодически происходящих перпендикулярно направлению предыдущих делений, Е—субапикальное боковое ветвление путем неравного деления верхушечной клетки; F—боковое ветвление из сегментов, расположенных позади верхушечной клетки, G—благодаря срастанию боковых ветвей возникает плоский тканевый таллом
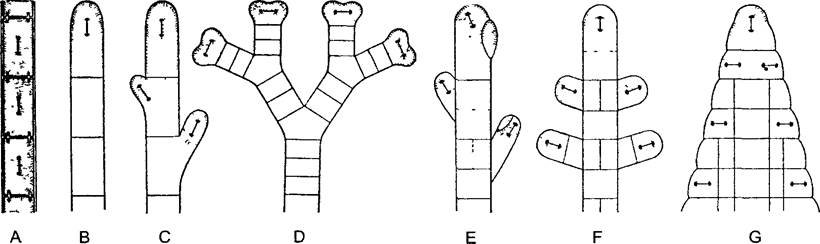
Когда в нитчатом талломе совершаются не только поперечные деления (веретено деления ориентировано вдоль), происходит ветвление (рис. 5.4, С —F). Ветви могут возникать из апикальной клетки, а также и из других клеток. Обычно соответствующие клеточные деления неравные, что ведет к иерархической системе ветвления: можно различить главную и боковые ветви, а при дальнейшем ветвлении — боковые ветви первого, второго и т.д. порядка (рис. 5.5). В результате клеточных делений, следующих, хотя и в одной плоскости, но в разных направлениях, может в конце концов образоваться плоский таллом. Из-за внешнего сходства с листом его называют филлоидом (см. рис. 5.4, G; 5.8, А).
Рис. 5.5. Длинный побег бурой водоросли Наlopteris filicna (по К.Goebel).
Верхушечная клетка в ходе неравных делений отчленяет сегменты, которые далее делятся поперечными и продольными стенками. Чередуясь с образованием сегментов, от верхушечной клетки косыми изогнутыми стенками отделяются инициали. формирующие боковые ветви (40х)
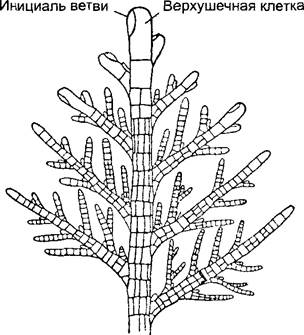
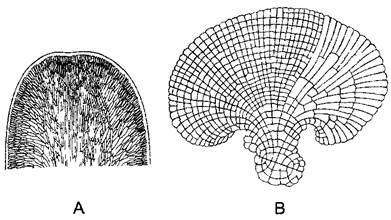
Рис. 5.6. Таллом красной водоросли, центрально-нитчатый тип, пример Сhondria tenuissima (по Falkenberg): А—продольный срез; В—поперечный срез
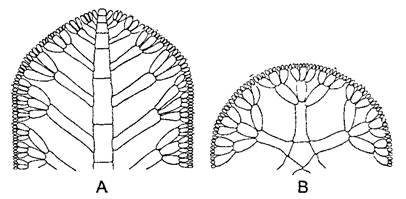
Рис. 5.7. Пластинчатые талломы красных водорослей (А—F.Oltmanns, В—по Rosanoff): А—верхушка ветви таллома Furcetlaria fästigiata (35x); В—однослойный таллом Metobesiaрастет веерообразно в одной плоскости благодаря нерегулярным продольным делениям краевых клеток(45х)
Самый известный пример талломов такого типа — родственник улотрикса морской салат Ulva lactuca (см. рис. 11.89, L), чьи лопастные талломы размером примерно с ладонь часто удается найти в приливной и прибойной зонах европейских морских побережий. Филлоид Ulva состоит из двух слоев клеток. Сравнение с родственными видами показывает, что они соответствуют однослойной стенке трубчатого таллома, который у Ulva, однако, сплющен. Необходимая механическая прочность достигается благодаря прочным на разрыв нитевидным («гифальным») клеткам, которые врастают между обоими клеточными слоями от ризоидного полюса филлоида. Это — простая механическая ткань1.
1 В большей мере они соответствуют идиобластам, а не ткани. — Примеч. ред.
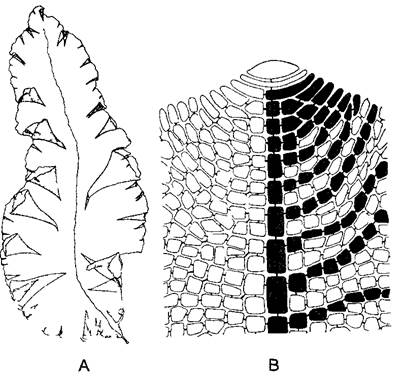
Талломы многих красных водорослей внешне сложно расчленены. Тем не менее настоящих тканей у них нет. Напротив, в основе расположения клеток, как проявления онтогенетического развития, лежит тип организации нитчатого таллома. Вегетативные тела большинства красных водорослей образованы множеством нитей, растущих согласованно, так что возникают вполне определенные макроструктуры. Отдельные нити в основном растут апикально. Они ветвятся или субапикально, благодаря чему возникает мутовка боковых нитей (центрально-нитчатый тип. рис. 5.6), или апикально (пластинчатый тип, рис 5.7) При этом могут возникать и плоские талломы (рис 5.8) В обоих случаях нити или срастаются (либо склеиваются) между собой в ложную ткань — псевдопаренхиму, которую в зрелом состоянии вряд ли можно отличить от типичной паренхимы, или же нити образуют плотное сплетение — плектенхиму (греч plectos — сплетенный) При этом только клетки одной нити связаны между собой порообразными структурами.
Рис. 5.8. Листовидный таллом красной водоросли Gnnellia americana (0,5х) (А) и передний конец однослойного таллома (В) с крупной верхушечной клеткой и образованной ею центральной нитью, последняя и некоторые отходящие от нее линии клеток зачернены (300х) (А—по R L Smith, В—по J Tilden)
5.3.2. Тканевые талломы
Высшей степени дифференцирования таллома среди водорослей достигают, с одной стороны, харовые (Сharophyсеае, рис 5.9, 11 106), с другой — бурые водоросли. Среди морских бурых водорослей есть самые крупные и долговечные водные растения Длина талломов некоторых ламинарий может быть более 100 м1. Они дифференцированы на органы прикрепления (несмотря на многоклеточность их и здесь называют ризоидами), аналоги стеблей — каулоиды (греч kaulos — стебель) и листовидные ассимиляторы — филлоиды. Этому соответствует сравнительно богатая дифференцировка клеток, позволяющая различать покровную, коровую и сердцевинную ткани.
1 Столь крупные размеры имеют бурые водоросли нескольких родов, но собственно ламинарии значительно мельче — Примеч. ред.
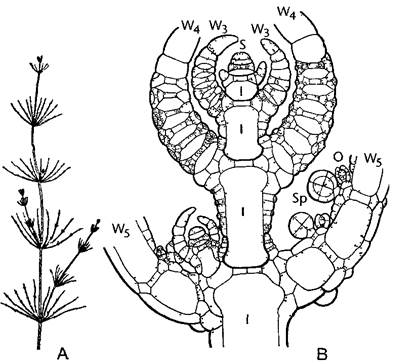
Рис. 5.9. Строение таллома харовой водоросли Chara fragdis (А—по A W Haupt, В—по J Sachs)
А—расчленение на узлы с мутовками ветвей и соединяющие их междоузлия На каждом узле может образоваться боковой побег (0,5х), В—продольный срез через верхушку таллома с апикальной клеткой Отчленяющиеся от нее клетки делятся вновь неравно на дистальную узловую клетку и базальную клетку междоузлия, которая обрастает корой из производных узловой клетки Клетки междоузлий во взрослом состоянии необычно крупные, в них наблюдается быстрое движение цитоплазмы Из наружных узловых клеток вырастают ветви мутовки (W3—W5), которые также расчленяются на узлы и междоузлия, на их узлах возникают оогонии и антеридии (30х), 1— клетки междоузлий, О—оогоний, S—верхушечная клетка, Sp—сперматогоний
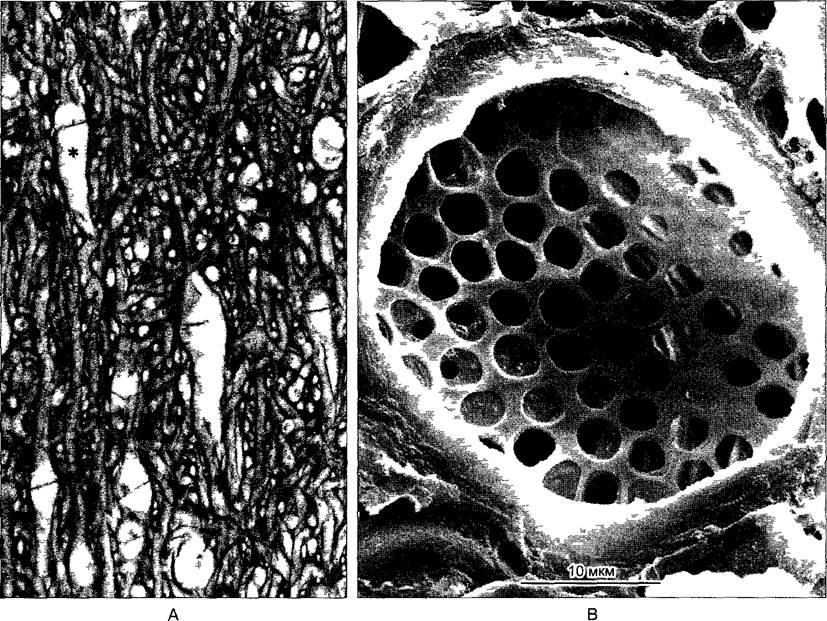
В каулоидах находятся так называемые ситовидные гифы, структурно и функционально соответствующие членикам ситовидных трубок покрытосеменных Диаметр пор их поперечных стенок, напоминающих ситовидные пластинки, достигают 6 мкм (рис 5.10) Сходство ситовидных гиф бурых водорослей и члеников ситовидных трубок высших растений (см рис 3 22) представляет собой аналогию, это — впечатляющий пример конвергенции.
Рис. 5.10. А—переплетение клеточных нитей в каулодии бурой водоросли Laminaria, видны широкопросветные ситовидные гифы (одна из них обозначена звездочкой) с поперечными ситовидными пластинками (150х) В—ситовидная пластинка в плектенхиме бурой водоросли Macrocystis integrifolia (СЭМ-фотография К Schmitz)