БОТАНИКА ТОМ 1 - КЛЕТОЧНАЯ БИОЛОГИЯ. АНАТОМИЯ. МОРФОЛОГИЯ - 2007
ВВЕДЕНИЕ
Ботаника как биологическая наука
Ботаника — наука о растениях. Название это было дано Диоскоридом (I столетие н.э.), который понимал под ним науку, изучающую (лекарственные) травы. Фактически греческое слово «botanе» означает злак, причем любое кормовое или полезное растение. Общее же греческое название для растения — «рhyton», поэтому уже многократно предлагалось назвать биологию растений «фитология», противопоставляя ее зоологии как науке о животных.
Понятие «растения» первоначально охватывало все те организмы, клетки которых, наряду с настоящим клеточным ядром, имеющим оболочку и несколько хромосом, содержат еще пластиды. Они или присутствуют в виде хлоропластов или в соответствующих условиях могут превращаться в них. Хлоропласты — это органеллы (клеточные органы), осуществляющие фотосинтез, т.е. превращение световой энергии в химическую и связанный с этим синтез органических соединений (ассимиляция углерода). Зеленые растения фотолитотрофны (фотоавтотрофны). В противоположность животному и всем прочим гетеротрофным (хемоорганогетеротрофным) организмам, зеленые растения обходятся без органического питания.
К растительному царству традиционно относят и грибы, хотя они и не имеют пластид. Грибы гетеротрофны и питаются либо мертвым органическим материалом (сапротрофно), либо живыми организмами (паразитно). (Авторы используют здесь более старый термин «сапрофитно». В научной литературе, в том числе и отечественной, в настоящее время все шире используется термин «сапротрофно». Действительно, слово «сапрофит» означает растение, питающееся мертвым материалом. Поскольку многие из этих организмов сейчас растениями не считаются (в частности, грибы), был предложен более обтекаемый термин «сапротроф», означающий просто «питающийся мертвым». — Примеч. пер.) Грибы эволюционно ближе к животным, но у них есть особенности, общие с растениями: например, вакуоли в клетках, одетых прочной клеточной стенкой, и поглощение питательных веществ в растворенной форме.
В группе одноклеточных организмов (протистов) различение растения и животного проблематично. У жгутиковых иногда в одном и том же роде — следовательно, у близко родственных видов — имеются формы без пластид и формы с хлоропластами: соответственно зоо- и фитофлагелляты (например, Еuglenа, рис. 1). У бактерий и архебактерий (архей) сколько-нибудь целесообразное отнесение к растительному или животному царству вообще невозможно. Эти организмы имеют клетки, которые в целом мельче и организованы принципиально проще, чем клетки всех животных, грибов и растений, даже одноклеточных (рис. 2). Бактерии и архебактерии не имеют настоящего клеточного ядра, здесь нет деления ядра и клетки в том виде, в каком они встречаются у прочих организмов, фототрофные формы не имеют пластид и т.д. Поэтому клетки этих групп иногда называют протоцитами в противоположность эуцитам всех прочих организмов. Бактерии и архебактерии как прокариоты противопоставляют эукариотам (растениям, грибам, животным; всем протистам с настоящим клеточным ядром). Между про- и эукариотами в современном (рецентном) мире живых организмов нет никаких переходов.
Рис. 1. Euglena gracilis (по G. F. Leedaie)
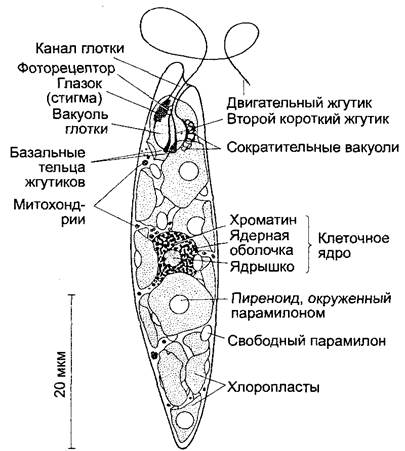
Рис. 2. Сравнение размеров про- и эукариотических клеток: А—бактериальные клетки из настоя сена (накопительная культура сенной палочки); В—клетки листочка мха (Мniumundulatum); можно отметить три характерные особенности растительной клетки: клеточные стенки, хлоропласты и центральные вакуоли. Обе фотографии имеют одинаковое увеличение (380х). На снимках в темном поле видны только те структуры объекта, которые рассеивают и преломляют свет, падающий сбоку

Это эукариотический одноклеточный организм с хлоропластами, которые образуют в качестве запасного вещества близкий к крахмалу парамилон. Двигательный жгутик выходит наружу из «глотки» (передней камеры). Его движения управляются светочувствительным органом (красной стигмой, часто неправильно называемой глазком, плюс фоторецептором на неподвижном коротком жгутике). Клеточная стенка отсутствует. Сократительные вакуоли обеспечивают выделение воды. Эвглены могут не только плавать, но и ползать, сильно изменяя при этом свою форму (2100х).
И все же древнейшие эукариоты развились из прокариот. Исследованию микроскопически малых организмов — как эу-, так и прокариотических — посвящена специальная биологическая наука — микробиология. В изучаемую этой наукой область входят и вирусы — субклеточные системы, которые находятся на границе живого и неживого.
Несмотря на все различия между прокариотическими и эукариотическими клетками, имеется много принципиально общих особенностей между ними, а еще больше — между клетками высших животных и растений, часто имеющими совершенно разные облик и функции. Повсюду мы встречаем сходные классы молекул, а
многие основные функции живых систем у всех организмов сходны. То же можно сказать и о многих генах (факторах наследственности). В этом выражается основополагающее единство всех живых организмов, которое свидетельствует об их общем происхождении; все ныне живущие организмы, очевидно, произошли от одного предка (мопофилетическое происхождение).
Что такое жизнь?
Поскольку мы сами являемся живыми организмами, то имеем непосредственное отношение к основному вопросу биологии: что такое жизнь? Тем не менее ответить на него оказывается довольно сложно. Правда, есть ряд свойств, которыми обладает каждая живая система, однако позволяет отграничить живое от неживых систем или образований только сумма этих признаков. Перечислим классические признаки жизни.
• Химический состав. В сухой массе всех живых организмов доминируют белки, нуклеиновые кислоты, полисахариды и липиды. К тому же имеется большая разнородная масса других органических молекул и ионов. Сложные органические молекулы, особенно макромолекулы, в природе синтезируются только живыми организмами (биосинтез с помощью специфических катализаторов — ферментов).
• Комплексная структура, имеющая системный характер. Жизнь всегда связана с живыми организмами. Даже простые существа отличаются комплексными структурами, которые имеют системный характер. Это означает, что молекулярные и надмолекулярные компоненты функционально взаимосвязаны между собой и согласованы друг с другом. Только за счет правильного взаимодействия — следовательно, совместно — они могут выполнять функции, обеспечивающие живое состояние. Ни один из отдельных компонентов, не говоря уже об отдельных веществах, не был бы способен на это в одиночку. Система представляет собой принципиально нечто большее, чем просто сумма частей, а жизнь — это всегда системная деятельность. Ниже уровня организации клетки самостоятельной жизни нет: клетки — это элементарные организмы. Они всегда содержат структуры, несущие информацию, полный набор необходимых ферментов и за счет селективно проницаемых мембран отграничены от окружающей среды. (Здесь нет противоречия с положением о том, что у большинства многоклеточных растений клетки тканей соединены между собой плазмодесмами — плазматическими каналами в клеточных стенках — в надклеточные симпласты.)
• Питание. Если рассматривать живые организмы в плане изменения энергии и энтропии, их существование может показаться очень неправдоподобным. Они состоят из богатых энергией, нестабильных молекул; высокая структурная и функциональная упорядоченность живых организмов соответствует низкому уровню энтропии. Поддержание лабильного состояния возможно только при притоке энергии, поэтому живые системы — принципиально открытые системы, т.е. они принимают богатые энергией фотоны или же вещества и отдают вещества, бедные энергией (например, СO2, Н2O; аналогия: пламя свечи). С обменом веществ (метаболизмом) неразрывно связан энергетический обмен. Метаболизм не приводит к стационарному равновесию. Наоборот, он всегда поддерживает неравновесное состояние (динамическое равновесие с необратимыми частными процессами). Обмен веществ и энергии позволяет сопрягать требующие энергии процессы биосинтеза специфических для организма (макро-)молекул (анаболизм, ассимиляция) с процессами, дающими энергию, — улавливанием солнечной энергии и/или разложением богатых энергией соединений (катаболизм, диссимиляция). Низкое содержание энтропии живых организмов поддерживается за счет отдачи (диссипации) избытков энтропии в окружающую среду. Только в качестве диссипативных структур живые организмы могут избежать гибельного для них хаоса. Таким образом, «жизнь» в конце концов не состояние, а всегда процесс. Если внешняя форма организмов чаще всего меняется достаточно медленно, то на молекулярном уровне происходит постоянная перестройка путем замены расщепленных веществ образуемыми заново (обмен, круговорот, turnover).
• Движение. Каждый активно живущий организм, и каждая отдельная клетка обладают движением. Правда, многие клетки/организмы могут переходить в покоящееся состояние и при этом образуют, например, семена, споры, цисты. На таких стадиях латентной (скрытой) жизни не только не различимы какие-либо движения, но и все остальные проявления жизни временно приостановлены.
• Восприятие раздражения и ответ на него. Все организмы и клетки для поддержания своей жизнеспособности приспособлены к тому, чтобы с помощью соответствующих рецепторов воспринимать сигналы из внешней среды и реагировать на них адекватными реакциями. Многообразие соответствующих механизмов чрезвычайно велико.
• Развитие. Живые организмы не способны длительно сохранять некую однажды достигнутую структуру. Ни один организм в разных фазах жизни не выглядит одинаково. Новая клетка, возникшая в результате деления, дорастает до размера материнской клетки (рост). Многоклеточные организмы начинают свое индивидуальное развитие в большинстве случаев с одной единственной клетки (оплодотворенной яйцеклетки — зиготы; споры). За счет размножения клеток они дорастают до своего окончательного размера. При этом изменяется и их облик. Развитие до половозрелого многоклеточного организма связано с морфогенетическими процессами. На клеточном уровне это проявляется в том, что первоначально одинаковые клетки зародыша приобретают все больше различий (дифференцировка).
• Воспроизведение. Последовательность поколений состоит из следующих друг за другом жизненных циклов. При этом жизнь всегда продолжается, несмотря на невозможность длительного сохранения определенной стадии развития отдельных особей и несмотря на неизбежную для многих многоклеточных организмов смерть каждого из них. Это составляет последний этап индивидуального развития. «Физиологическая смерть», в противоположность «катастрофической смерти», часто осуществляется исходя из внутренних причин в результате реализации генетически предопределенной программы самоуничтожения. Наоборот, организмы могут возникать только как потомки подобных их предков. Самозарождение (возникновение живых систем из неживой материи) в настоящее время на Земле крайне мало вероятно и к тому же никогда не было зарегистрировано: оmnе vivum е vivo (все живое из живого). Это ныне само собой разумеющееся положение не является столь
уж давним. Так, например, до основополагающих исследований Л. Пастера и Г. Гофмана в середине XIX столетия считалось, что микроорганизмы (а также грибы и черви) в жидкостях, где происходят процессы гниения и брожения, могут возникать сами по себе (что было опровергнуто опытами со стерилизацией питательных растворов).
• Размножение. Размножение обычно связано с увеличением численности особей. Только таким образом возможно воспроизведение вида, несмотря на различные потери в той или иной мере из-за воздействия окружающей среды. Повсюду мы наблюдаем очень высокие скорости размножения (особенно у мелких организмов). Бактериальные клетки часто при оптимальных условиях делятся каждые 20 мин. Это означает, что при размножении, которому ничто не препятствует, одна-единственная клетка уже в пределах двух дней может дать такое потомство, которое за два дня достигает объема земного шара. У более крупных организмов скорость размножения обычно значительно меньше, поэтому жизнь каждого индивидуума лучше защищена разного рода приспособлениями.
• Наследственность. Индивидуальное развитие протекает в ряду следующих друг за другом поколений более или менее одинаково. Генетическая информация всех клеточных организмов (про- и эукариот) содержится в последовательности оснований (нуклеотидов) молекул дезоксирибонуклеиновой кислоты (ДНК, в международной литературе — DNA — от англ, deoxyribonucleic acid). Здесь речь идет о линейных или циклических макромолекулах, состоящих из двух нитей (двухцепочечная ДНК, dsDNA). У вирусов генетическая информация может также передаваться через молекулу ДНК, состоящую из одной цепи, а также через рибонуклеиновые кислоты (РНК = RNA, состоящие из одной или двух цепей).
• Эволюция. Копирование (репликация) и дальнейшая передача генетической информации осуществляются с высокой точностью, однако в последовательном ряду нескольких поколений могут появиться изменения, которые наследуются (мутации). За длительный период времени в популяциях проявляются значительные различия между особями, которые в дальнейшем имеют те или иные шансы оставить потомство. Согласно теории естественного отбора, обоснованной в 1859 г. Ч. Дарвином и независимо от него Р. Уоллесом, в природе постоянно накапливаются формы, все более приспособленные для продолжения вида. Это приводит к изменению особенностей представителей видов и в конечном счете к возникновению новых видов: эволюция, филогенетическое развитие (филогенез).
Как важнейший критерий жизни, по- видимому, можно рассматривать имеющуюся у всех организмов способность к воспроизведению. Все другие характеристики представляют собой либо предпосылки, либо следствия этого центрального свойства. У всех организмов генетическая информация содержит план развития для очень сложного молекулярного механизма, основная функция которого — самовоспроизведение. Жизнь (по крайней мере на современной Земле) может быть доказуема и мыслима только как континуум. Это подчеркивается необратимостью индивидуальной смерти и вымиранием видов.
Математиком Дж. фон Нойманом была разработана общая теория самовоспроизводящихся систем. «Ноймановский автомат» включает четыре компонента: (1) устройства для производства всех системных элементов; (2) необходимая для этого информация; (3) множительное устройство для точного размножения (2); (4) регулятор для соответствующей программы последовательности всех единичных процессов. Совместное функционирование четырех компонентов приводит к циклическим процессам, в которых система сама себя воспроизводит. Живая клетка — это ноймановский автомат1.
1 В нашей стране под руководством акад. В. Б. Кудрявцева разработана теория клеточных автоматов, базирующаяся на тех же принципах. — Примеч. ред.
Происхождение и эволюция жизни
Современный (рецентный) мир живого — результат невообразимо длительной эволюции. Исходя из природной радиоактивности и состава наиболее древних скальных формаций возраст Земли можно оценить в 4,6 млрд лет. Изучение остатков различных организмов (ископаемых: палеонтология) в отложениях разного возраста показывает, что в более ранние эпохи истории Земли на ней жили растения и животные, отличные от современных. Филетическая непрерывность проявляется в том, что флоры и фауны прошлых эпох тем менее сходны с современным миром организмов, чем дальше они отстоят во времени от нас. Более крупные многоклеточные организмы обнаруживаются только к концу докембрия (ок. 570 млн лет назад). До этого доминировали одноклеточные организмы, а среди них прежде всего прокариоты. Указания на обширные колонии цианобактерий имеются уже из архея (>3 млрд лет назад): отложения такого возраста в Австралии и Южной Африке содержат слоистые строматолиты размером до 30 см и более. При этом речь идет о характерных биогенных отложениях, которые еще и сейчас образуются в теплых водоемах плотными образованиями фототрофных цианобактерий и других организмов (цианобактериальные маты).
Как могла возникнуть жизнь? К ответу на этот фундаментальный вопрос биологии пытаются подойти с помощью экспериментов, воспроизводящих (моделирование) те условия, которые предположительно господствовали в древнейшие времена на Земле1. Предпосылкой для образования простейших самовоспроизводящихся систем было наличие органических (макро-)молекул. В противоположность нашему времени на еще горячей древней Земле органические соединения могли возникать абиогенно. Древняя атмосфера наряду с водяными парами содержала прежде всего диоксид углерода и азот, предположительно также незначительные доли восстанавливающих газов, однако практически не содержала свободного кислорода. Поэтому не было озонового слоя, который бы поглощал богатое энергией ультрафиолетовое излучение Солнца. В таких условиях могли спонтанно образовываться различные органические соединения. Даже в водных смесях монооксида углерода, сероводорода и сульфидов металлов, которые, например, выбрасываются горячими источниками из водных глубин, могут возникать абиогенным путем уксусная кислота и богатые энергией тиоэфирные связи между молекулами. Такие соединения, по-видимому, накапливались в некоторых местах древней Земли, тем более что живые организмы, использующие их для своего питания, еще не существовали, и не могло происходить их разрушение за счет окисления.
1 Эксперименты по имитации условий, в которых возникали первые органические молекулы, были впервые поставлены в нашей стране академиком А. И.Опариным. Ему принадлежит описываемая здесь теория. — Примеч. ред.
Чисто умозрительно простейшие клетки — примерно такие, как у ныне живущих сапротрофных микоплазм (см. ниже), уже настолько сложно устроены, что их возникновение из хаотической смеси молекулярных компонентов в результате одного единственного случайного события представляется совершенно невероятным. Однако возникновение простейших самовоспроизводящихся систем можно объяснить хотя бы спекулятивно как последовательность гипотетических промежуточных этапов: гипотеза многих этапов. Если необходимые отдельные этапы этой пребиотической эволюции были достаточно малыми, вероятность того, что они могли случиться на протяжении очень длительных периодов времени, становится достаточно большой. Некоторые молекулы, которые могли возникнуть абиогенно, обладают ферментативой активностью, т.е. действуют как биокатализаторы. Определенные молекулы РНК (рибозимы) могут при этом сами по себе катализировать некоторые химические реакции, а вместе с ионами тяжелых металлов даже управляют собственным размножением, хотя и очень несовершенным образом («мир РНК»). Решающий шаг к собственно жизни был сделан, когда стала возможной эффективная и точная репликация нуклеиновых кислот, происходящая при участии белков-катализаторов, а синтез этих белков-ферментов стал осуществляться на основе информации, заключенной в нуклеиновых кислотах. Благодаря этому двойному прогрессу, который предположительно явился результатом многих отдельных мелких этапов, возникла взаимосвязь между белками и нуклеиновыми кислотами, составляющая основу всякой жизни в ее современной форме. Теперь имелся генетический код для перевода последовательностей нуклеиновых кислот в полипептидные последовательности белков, и произошло разделение гена (наследственного фактора) и фена (признака, возникающего на основе наследственной информации).
Пока происходило абиотическое образование органических молекул, первые способные к размножению системы — гипотетические прогеноты — и развившиеся наконец из них прокариоты могли жить органотрофно. Однако по мере все большего потребления и исчерпания органических источников питания на передний план стали выходить фототрофные формы, в том числе и такие, которые при фотосинтезе расщепляли воду и высвобождали кислород. За счет этого атмосфера становилась все более окисляющей, что создало предпосылки для значительно более эффективного получения энергии из органических веществ путем клеточного дыхания. Одновременно в стратосфере возник озоновый слой1, который стал поглощать сильное мутагенное ультрафиолетовое излучение Солнца и этим сделал возможным заселение суши.
1 Возникновение озонового слоя — более позднее событие, чем возникновение дыхания. Расчеты показывают, что для дыхания достаточно 0,2 % О2 в атмосфере (точка Пастера), а для возникновения озонового слоя — 1—2% О2. — Примеч. ред.
Ископаемые находки времен длительной докембрийской эволюции, разумеется, редки и соответственно фрагментарны. Однако с помощью сравнения последовательностей белков и нуклеиновых кислот можно устанавливать степень родства современных организмов и реконструировать ход эволюции. Чем больше различаются последовательности соответствующих белков, ДНК и РНК, тем раньше должны были жить последние общие предки сравниваемых организмов. Изменения в ходе эволюции могли происходить у различных (частных) последовательностей с разной скоростью, поэтому для реконструкции раннего филогенеза выбирают такие последовательности (или участки последовательностей), которые изменяются лишь очень медленно и даже у совершенно непохожих современных организмов еще в значительной мере сходны. Из сравнения таких очень консервативных последовательностей можно заключить, что разделение архей и бактерий произошло уже более 3 млрд лет назад1. У современных эукариотических клеток пластиды и митохондрии (соответственно органеллы фотосинтеза и клеточного дыхания) обладают собственной генетической информацией и сами синтезируют часть своих белков. Они могут возникать только из себе подобных и занимают, таким образом, в эукариотических клетках (полу-)автономное положение. Кроме того, они обладают многочисленными свойствами прокариот, например, в характере деления и деталях своего химического состава. По крайней мере, пластиды, очевидно, представляют собой потомков некогда свободно живших бактерий, которые более миллиарда лет назад внедрились в качестве внутриклеточных симбионтов в клетки примитивных эукариот и постепенно развились в клеточные органеллы (теория эндосимбиоза).
1 Эти количественные оценки получены исходя из предположения, что мутации в генах происходят равномерно во времени. Теоретические основы сравнения последовательностей ДНК заложил Кимура (Япония): теория нейтральности. — Примеч. ред.
Остатки многоклеточных макроорганизмов встречаются только в отложениях, которые не старше миллиарда лет. Такие организмы — исключительно эукариоты. Их эволюция, которая благодаря данным палеонтологии становится все более реконструируемой, также происходила прежде всего при взаимодействии случайных мутаций и задающего направление отбора (теория отбора, дарвинизм). При этом полагают, что эволюция является результатом суммирования бесчисленных мелких изменений (градуализм). Постоянно происходят и макроэволюционные ароморфозы(major evolutionary transitions), однако отличаются они не по виду своего осуществления, а скорее по эффекту от постепенных эволюционных изменений. Ароморфозы, хотя они и происходили значительно реже, чем прочие постепенные эволюционные шаги, но имели далеко идущие последствия. Очевидно, репродуктивные единицы, которые до определенного момента развивались самостоятельно, постепенно становились заключенными во все более крупные и значительно более сложные структуры. За счет этого возникали полностью новые системы, которые могли становиться исходными пунктами эволюционных линий другого, типа.
Наглядный пример такого крупного перехода — возникновение в ходе эволюции многоклеточных организмов из одноклеточных. Отдельная клетка, которая до этого перехода представляла собой целый организм, у многоклеточных организмов — только один из многих элементов целого тела. Отбору подвергаются уже не отдельные клетки, как раньше, а общая надклеточная система. Если она погибает, погибают и все ее клетки. Встраиваясь в более крупную систему, обеспечивающую стабильные условия жизни, клетки в значительной мере утрачивают самостоятельность. Система в целом определяет, например, деление отдельных клеток и их особые функции и продолжительность жизни при выполнении самых разных функций. При этом реализуется важная функция таких систем: здесь имеет место «разделение труда». В многоклеточных организмах отдельные клетки могут специализироваться для выполнения определенных частных задач, тогда как остальные клетки в этом не участвуют. За счет этого не только частные процессы важных путей обмена веществ могут выполняться с большей эффективностью, но и синергетические потенциалы будут использоваться полнее. В ходе дальнейшего филогенеза дифференцированные клетки или целые ткани в количественном отношении увеличивались или уменьшались, и они могли, как модули (кубики «конструктора»), перемещаться в общей системе или заново комбинироваться. Такая комбинаторика при наличии относительно небольшого числа различающихся элементов способствует развитию почти бесконечно большого числа различных систем. С этим связано необычайно большое разнообразие видов и форм макроорганизмов, которые нас окружают.
Другие крупные эволюционные изменения связаны с симбиозом. За счет тесной совместной жизни неродственных организмов могут возникать принципиально новые системы с более высоким уровнем организации. Вышеупомянутая теория эндосимбиоза — хороший пример этому. Эукариотические клетки в свете теории симбиогенеза собственно не являются клетками, их нужно рассматривать как мозаичные образования, возникшие в результате внутриклеточного симбиоза клеток древнейших эукариот («уркариот») с бактериальными клетками. Обсуждается даже такое предположение, что первые эукариотические клетки, в свою очередь, были результатом симбиоза архебактерий и эубактерий.
Границы жизни
Вопрос о границах жизни имеет двоякий смысл: с одной стороны, как вопрос о границах распространения живых организмов, а с другой стороны, как вопрос о мельчайших или же наиболее крупных живых организмах. Первый аспект составляет предмет экологии; можно сказать, что, несмотря на исключительно большой
возраст механизмов адаптации, общие потребности живых организмов остаются в весьма узких границах. Они определяются прежде всего максимумами и минимумами содержания воды, температурой и светом. Оптимум для большинства организмов находится в области средних температур (10 —40°С) и большого содержания воды. Соответственно продукты питания можно сохранить от разложения разными организмами в пригодном к употреблению виде в холодильнике (в частности, в морозильной камере) или путем высушивания (бобовые, злаки, мука, хлеб, макаронные издели..; сено), или путем стерилизации нагреванием (молоко...). В дикой природе особенно холодные и сухие области заселены лишь скудно или вообще безжизненны. Вместе с тем, покоящиеся стадии многих организмов могут выживать при температурах, близких к абсолютному нулю, однако жизненные функции становятся невозможными при температурах в пределах от 0 до -10 °С. Температуры до 100 °С и выше, которые на Земле встречаются лишь в немногих местах (горячие источники, вулканы), могут, напротив, быть пригодными для обитания термофильных организмов. Некоторые архебактерии имеют температурный оптимум порядка 100 °С; возможно, эта способность сохранилась как реликтовая с древнейших времен истории Земли. Поскольку в качестве продуцентов органического материала (биомассы) в значительной мере выступают только фототрофные организмы1, жизнь преимущественно ограничена хорошо освещаемыми областями поверхности суши и океанов. Земля покрыта относительно тонким слоем биосферы. Последняя не составляет и сотой доли процента объема Земли.
1 Имеются сведения, что значительную — и даже, может быть, большую, чем фотоавтотрофы, роль как продуценты органического материала имеют хемоавтотрофные организмы, особенно распространенные на больших глубинах мирового океана. — Примеч. пер.
Особенно крупные организмы встречаются среди ископаемых и ныне живущих форм позвоночных животных (динозавры, беззубые киты), хвойных и лиственных деревьев (даже в значительно большем числе видов и особей), хотя это не столь заметно на первый взгляд — у взаимосвязанных клонов некоторых растений (например, пирамидальных тополей, камыша, папоротника-орляка) и грибов. Гиганты среди деревьев (секвойи, криптомерии, некоторые эвкалипты1) — одновременно имеют и наибольшую массу. (Гигантские киты имеют, несмотря на огромную массу, не такую большую плотность, их вес компенсируется подъемной силой.)
1 Имеются сведения, что наибольшей высоты — до 145 — 150 м — достигают некоторые экземпляры лжетсуги Мензиса (Psedotsuga menziesil). — Примеч. пер.
Для теоретической биологии более важен вопрос, как могут существовать мелкие живые организмы: где находится нижняя граница сложных самовоспроизводящихся биосистем? Мельчайшие клетки имеют прокариотическую организацию. Они встречаются у микоплазм. Диаметр этих прокариотических клеток, лишенных клеточной стенки, составляет порядка 0,3 мкм; их ДНК может кодировать всего лишь около 500 разных белков. Это близко к минимуму того, что необходимо для воспроизведения ДНК, реализации содержащейся в ней генетической информации, поддержания гетеротрофного обмена веществ и энергии и простой клеточной структуры (приблизительно 350 генов). Для сравнения: клетки типичных бактерий имеют диаметр порядка 2 мкм и содержат свыше 3 000 различных белков; диаметр же большинства эукариотических клеток находится в пределах 10 и 100 мкм, они могут образовывать до 30 000 и более различных белков. Полностью секвенированный геном резушховидки Таля (Агаbidopsis thaliana) содержит около 25 000 генов, что на 11 000 больше, чем у дрозофилы.
Вирусы имеют значительно более простую организацию, чем клетки микоплазм, большинство из них также значительно мельче. Однако вирион (вирусная частица) не является клеткой. Если, например, даже простейшая клетка содержит как ДНК (как носитель информации), так и РНК (для реализации генетической информации), то у вириона лишь один тип нуклеиновых кислот: либо ДНК, либо РНК. Нуклеиновая кислота часто ассоциирована только с одним-единственным видом белка, как у вируса табачной мозаики (ВТМ, рис. 3), или она окружена белковой оболочкой (капсидом), которая состоит из одного-единственного белка или немногих разных белков. Образованный таким образом капсид вируса часто обладает кристаллической симметрией. Вирусы, в том числе (бактерио-)фаги (вирусы, которые поражают прокариотические клетки), лишь частично соответствуют критериям жизни. Они не осуществляют обмен веществ и энергии, не обладают сами способностью к репликации и синтезу белка и поэтому не могут размножаться самостоятельно. Эту функцию они реализуют только, используя обмен веществ и энергию живых клеток, т.е. являются облигатными внутриклеточными паразитами («жизнь взаймы»). Вне живых клеток вирионы,представляющие собой форму для распространения, — это безжизненные органические системы.
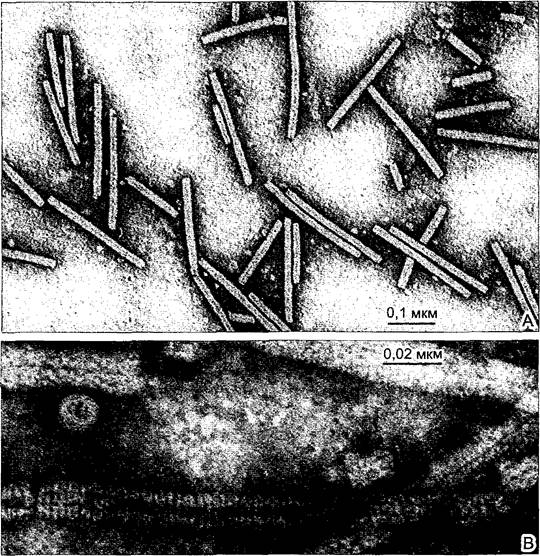
Рис. 3. Вирус табачной мозаики (ВТМ) (электронно-микроскопические снимки А — F. Amelunxen; В — С. Weichan)
При электронной микроскопии (ЭМ) частицы вируса выглядят как палочки. Вирион содержит спиральную молекулу РНК, на которой у неповрежденных вирионов расположены в ряд 2 130 идентичных молекул белка, каждая состоит из 158 аминокислотных остатков. Образуемый спиралью РНК центральный осевой канал четко виден в этих негативно-контрастных препаратах
Простейшие уровни организации достигаются вироидами — инфекционными нуклеиновыми кислотами (РНК) без сопровождающих белков. Очень короткие кольцевые молекулы РНК не кодируют ни одного белка. Среди вироидов известны опасные паразиты растений.
Несмотря на особенно простую организацию, вирусы и вироиды не могут рассматриваться как исходные формы жизни, поскольку их размножение предполагает наличие живых клеток. Скорее здесь речь идет о генетических элементах, которые могли частично обособиться от своих клеток-носителей (эгоистичные генетические элементы). Фактически у многих (вероятно, что даже у всех) эу- и прокариот имеются сегменты генетической информации, которые либо наследуются независимо от основных несущих гены структур (хромосом, генофоров), либо могут, хотя бы временно, отделяться от них. К этой гетерогенной группе относятся, с одной стороны, плазмиды многих бактерий и некоторых эукариот, а с другой стороны — так называемые инсерционные последовательности и транспозоны («прыгающие гены»).
Биология как естественная наука
Живая природа впечатляет прежде всего огромным разнообразием организмов. Изучение, описание и систематическое упорядочение всех ныне живущих и вымерших видов организмов — очень большая, всегда до конца не выполнимая задача биологии, главным образом, систематики. Однако задача биологии не исчерпывается констатацией имеющегося; скорее исследователи пытаются вскрыть закономерности, лежащие в основе этого многообразия. Наряду с обычным наблюдением и сравнением здесь участвует эксперимент — наблюдение за процессами при искусственно задаваемых или же варьирующих условиях. Правда, результаты экспериментов или наблюдений дают лишь сырой материал для построения гипотез и теорий, т.е. для выяснения причинных взаимосвязей. (А. Пуанкаре: «Накопление фактов столь же мало представляет собой науку, сколь и куча камней — уже готовый дом».) За счет обнаружения закономерных взаимосвязей и их окончательной формулировки как законов природы сколь угодно многочисленные данные специальных наблюдений могут быть обобщены и осмыслены в короткой, наглядной форме. Без этой абстракции возможность интеллектуального проникновения в реальный мир с его принципиально необъятным обилием структур и событий была бы исключена. Только открытие законов природы делает возможными понимание естественных состояний и процессов (их объяснение), предсказание событий и, наконец, разумное применение достигнутых наукой результатов. На этом основывается колоссальное значение естественных наук в наше время. Особое место в этом плане занимает и современная биология (ключевые слова: биотехнология, генная инженерия).
Сумма ставших известными законов природы (не отдельные данные!) и их интерпретаций создает естественно-научную картину мира — упрощенное отражение природы в понятиях, символах и представлениях. Эта картина мира — высшее выражение нашего знания о природе. Она позволяет производить умственные операции (мысленные эксперименты), которые в реальном мире были бы дорогостоящими, опасными или вообще невыполнимыми. Естественно-научная картина мира в основном открытая (динамичная), т.е. с прогрессом исследований и новыми интерпретациями постоянно расширяющаяся и изменяющаяся, поэтому она неизбежно имеет временный и фрагментарный характер и никогда не может рассматриваться как окончательная. И тем не менее она одновременно является лучшей, чем человечество может располагать в этой области. Фрагментарный характер естественно-научной картины мира связан не только с произвольными, хотя и не всегда осознанными и принимаемыми во внимание границами установок естественно-научных целей (например, отсутствуют эстетические, этические и трансцендентальные параметры), но и также с методическими ограничениями, прежде всего со способом поиска новых знаний. Такой поиск в исследовании основ мироздания, направленный на познание мира, а не на его изменение и господство над ним, может быть не прямым, так как цель (окончательный результат) заранее не известна. При косвенных поисках цели перепроверяемые попытки объяснения делаются в форме гипотез (греч. hypothesis — подтасовка). Однако гипотеза, вообще научная концепция, может оказаться при этом не полностью соответствующей столь многим согласующимся данным, что число подтверждающих ее фактов будет слишком мало на фоне неограниченно большого числа мыслимых противоречий. Напротив, общее высказывание (обобщение) может быть опровергнуто одним-единственным противоречащим результатом (асимметрия верификации и фальсификации: К. Р. Поппер). Утверждение «Все розы имеют красные цветки» не может быть доказано даже наличием тысячи красных роз, но опровергается наличием одной- единственной желтой или белой розы.
Корреляции выражают закономерные отношения на уровне доступных наблюдению явлений (например, курение сигарет/рак легких; частота встречаемости аистов/рождаемость в определенных местностях). Корреляции могут, но не обязаны означать причинную связь. Если величины В и С коррелируют, то В может быть причиной С или наоборот. Однако В и С могут иметь и третью, пока не принимавшуюся во внимание величину А в качестве общей причины, тогда они, хотя и коррелируют, но каждая со своей стороны не состоит с другой в причинной связи. Таким образом, если отсутствующая корреляция указывает на отсутствие причинной связи, даже достоверная корреляция еще не служит ее доказательством, т. е. не может быть верификацией соответствующего предположения.
Из-за асимметрии верификации и фальсификации прогресс познания достигается не прямым, а косвенным путем опровержения неподходящих гипотез (метод проб и ошибок — англ, trial and error). Цель — соответствующее действительности знание и объясняющее понимание — может быть достигнута только путем разочарования и окольными путями (греч. слово «methodos» означает не только «основательное исследование», но и также «окольный путь»).
Конечно, с каждой неудавшейся попыткой фальсификации повышается вероятность того, что гипотеза оправдается, особенно если гипотезу удается успешно применить к опытам из других областей науки, произведенным независимо от нее. Обобщающие гипотезы, которые, несмотря на многие попытки, не были фальсифицированы, трактуются как теории.Теории — это элементы естественно-научной картины мира. Исходя из какой- либо теории (например, центральной для биологии теории эволюции), можно объяснить множество данных. На основе научной теории можно сформулировать многочисленные экспериментально проверяемые утверждения. В научно-теоретическом плане теория представляет собой дисциплинарный матрикс — парадигму, которая создает интеллектуальные рамки для дальнейшей экспериментальной работы в соответствующей области. Поскольку целенаправленные наблюдения и обдуманные эксперименты могут проводиться только на основе гипотез или же теорий, большая часть исследований удивительным образом совершенно не индуктивная (т.е. не исходит из опыта и не ведет к соответствующему действительности пониманию), а дедуктивная. Прежде всего эти опыты не направлены на открытие неожиданного и нового, а служат для дополнения или же перепроверки имевшейся ранее парадигмы. Конечно, даже считающиеся «достоверными», всеобще признанными теории в некоторых случаях оказываются фальсифицированными. Тогда должна быть найдена новая, всеобъемлющая теория. Однако такие научные революции (Л. Флек, Т. С. Кун) удаются только тогда, когда новая теория в состоянии объяснить, почему предшествующая ей теория соответствует столь многим фактам. Часто оказывается, что старая теория в рамках определенных, сначала не осознаваемых границ остается действенной. В истории научной биологии встречаются многие примеры подобных революций, в частности развитие учения о клетке и генетики.
Представленные здесь рассуждения — это часть эпистемологии (учения о возможностях и границах человеческого познания), которой не только в теоретических естественных науках, но и в философии отводится центральное положение (например, у И. Канта). При этом долгое время оставалось загадкой, почему имеются независимая от опыта логика, далее математика и др., которые все-таки везде могут быть приложимы к живой природе. (А. Эйнштейн: «Самое непостижимое в мире — это его постижимость».) Эта загадка была принципиально решена благодаря исходящей из биологии эволюционной теории познания (К. Лоренц, Г. Фольмер и др.): даже независимые от индивидуального опыта (априори) высказывания логики, математики основываются в конечном счете на опыте, который был, правда, накоплен уже во время эволюции гоминид во многих поколениях и при постоянном подтверждении, наконец, зафиксирован гентически1, поскольку давал мощное селективное преимущество.
1 Здесь можно поспорить с немецким автором, поскольку логика заключена в структуре языка. Язык — система негенетической передачи информации из поколения в поколение. Способность к логике и математике развивается тренировкой. — Примеч. ред.
Особое положение биологии
Особому положению живых организмов в природе соответствует особое положение биологии среди естественных наук. Постоянно ставится вопрос, не подчиняются ли живые системы закономерностям, отличным от имеющихся в неживой природе, и часто постулируются особые жизненные силы (витализм). Вместе с тем до нынешнего времени не известно такого случая, когда физические и химические закономерности не действовали бы среди живых организмов. Однако, с другой стороны, в мир живых организмов привносятся совершенно невероятная сложность и системный характер, так что в биологии проявляются закономерности, которые нигде больше не наблюдаются. Речь идет об эмергентных свойствах. Важное следствие сложности живых систем состоит в том, что суть биологии в логическом плане или же с помощью математических методов не может быть постигнута так, как объекты физики и химии 2. Правда, биология является по своей сути точной номотетической (ориентированной на познание закономерностей) естественной наукой, но наблюдения, описания и сравнения играют в ней значительно большую роль, чем, скажем, в физике. Полное сведение всех биологических явлений к закономерностям, известным из области химии и физики, как это требовалось бы с позиций последовательного редукционизма, во всяком случае, является иллюзорным.
2 Такое противопоставление биологии и физики/химии нам представляется спорным. С одной стороны, в полном математическом и логическом описании объектов в химии и физике остается большая «недосказанность» (иначе теоретические исследования в этих науках прекратились бы). С другой стороны, биологи все еще недостаточно стремятся к познанию живого с помощью логики и математики, поэтому соответствующие разделы «Математики живых систем» все еще не разработаны. — Примеч. ред.
Характеризуя живые организмы как самовоспроизводяшиеся системы, мы подходим к дальнейшему пункту, который поясняет особое положение организмов: к биологической телеономии. Живые организмы ведут себя целенаправленно, они целесообразно реагируют и выглядят разумно сконструированными. Наряду с вопросом «почему?» (каузальность), в биологии (и среди естественных наук только в биологии) целесообразен и правомерен также вопрос «для чего?» (фннальность). Это основывается в конечном счете на циклическом развитии живых организмов (ср. понятия «цикл развития, размножения или поколений»). От некой данной исходной ситуации эти циклы ведут по генетически установленным путям развития опять к подобным же исходным ситуациям (например, одноклеточным формам, спорам). Благодаря этому возникают квазициклические цепи событий и причинные цепи. Например, определенное состояние развития В выходит не только как следствие предшествующего А, но и через последующие состояния С, D... и одновременно опять как причина для повторных (даже если и не упорядоченных во времени) проявлений А. Финальный вид рассмотрения выступает поэтому в биологии почти равноправно наряду с каузальным. В неживой природе циклические процессы (например, колебания) не обладают механизмами, которые бы компенсировали потери из-за процессов затухания, и приходят в конечном счете к состоянию покоя1. Напротив, живые организмы могут при размножении еще и увеличивать свою численность.
1 На макроуровне неживая природа Земли до сих пор не пришла к состоянию покоя: круговорот воды (течение рек и ледников), движение литосферных плит, атмосферные потоки, по-видимому, служили посылкой для возникновения жизни на нашей планете. — Примеч. ред.
Также и при изучении эволюции и происхождения жизни биология занимает среди естественных наук необычное положение. Если там прежде всего выявляют закономерности, которые отличаются регулярными повторениями структур или процессов, то здесь решающую роль часто играет единичное, случайное событие. Это связано с размножением и отбором организмов. Природные мутации — случайные события, единичные и непредсказуемые. Если мутация ведет к благоприятным последствиям для ее носителя, то, согласно теории отбора, она вновь и вновь дает преимущества в последующих поколениях. Живые организмы «работают» в этом плане как чрезвычайно действенные усилители: многие (все?) из наблюдаемых у них наследуемых свойств происходят от крайне маловероятных и соответственно редких случайных событий (единичных случаев), которые, однако, в дальнейшем путем процессов размножения необычайно распространились и усилились. Не исключено, что, например, возникновение жизни или «открытие» генетического кода, который у всех организмов почти не имеет отклонений, имели в основе единичные события, зафиксированные в ходе земной жизни и размноженные при воспроизведении организмов в гигантском масштабе. Возникновение и эволюция жизни — хорошие примеры детерминистического хаоса, хотя и находящегося под господством законов и не лишенного причинности, однако из-за детерминирующих случайных событий не являющегося ни предсказуемым, ни полностью понимаемым.
Животное и растение
После преодоления (более исторически, чем объективно) обоснованной специализации науки на зоологию и ботанику в современной биологии доминирует междисциплинарный общий подход: генетические, биофизические и биохимические, а также физиологические данные образуют широкий фундамент общей биологии; так же эволюционная биология и биология развития, как и молекулярная и клеточная биология, оказались выше границ «классических» предметов ботаники и зоологии. Однако при таком подразделении не следует забывать, что типичное животное и типичное растение (оба термина понимаются в обиходном смысле) показывают многочисленные существенные различия.
Типичное животное способно к перемене места, поэтому его тело построено компактно, а все органы, за исключением органов чувств, предназначенных для восприятия сигналов из внешней среды, находятся внутри организма. Чтобы увидеть строение тела животного, тело должно быть вскрыто (анатомия — от греч. слова, означающего «вскрытие»). Обширные поверхности, необходимые для дыхания, питания и выделения, развиваются как впячивания внутрь тела. Внешняя поверхность сводится к минимуму: животное — «закрытый» организм. Компактное строение тела обеспечивает центральное расположение органов для кровообращения и выделения. Также и нервная система, которая делает возможной быструю координацию, в ходе филогении показывает тенденцию к централизации. Большинство органов формируется в ограниченном числе. Симметрия тела преимущественно билатеральная и дорсовентральная, соответственно двум ориентированным перпендикулярно друг другу векторам силы тяжести и движения. В узком смысле радиально-симметричные формы встречаются почти только у прикрепленных или парящих в толще воды форм. Специализация тканей и органов идет очень далеко. Уже образовательные ткани часто специализированы для последующего формирования совершенно определенных типов клеток (стволовые клетки крови и иммунной системы, кожи, эпителия кишечника и т.д.). Продолжительность жизни даже крупных животных ограничена. Функции регенерации у высокоразвитых животных незначительны. У них некоторые высокодифференцированные клетки остаются активными на протяжении всей жизни и обычно у взрослого животного больше вновь не образуются (крупные нейроны; поперечнополосатые мышечные волокна; клетки хрусталика глаза).
Типичное растение, напротив, ведет прикрепленный образ жизни. Оно развивает многие свои органы (корни, листья, цветки) в большом числе и свободно наружу. Поверхность тела за счет выпячивания и ветвления увеличивается до максимума. Растение — «открытый» организм. Многолетние растения продолжают расти в каждый период вегетации, имея многочисленные точки роста (у деревьев: ежегодный прирост всех побегов, годичные кольца древесины и т.д.). Открытая организация тела растения ограничивает развитие центральных органов, у растений нет ни сердца, ни почек, ни органов, аналогичных нервной системе. Отходы обмена веществ должны удаляться каждой отдельной клеткой самостоятельно; вместо центрального выделения здесь имеется локальное, клеточное. Тело чаще всего радиально-симметричное; билатеральные органы в основном образуются только тогда, когда векторы силы тяжести и роста направлены перпендикулярно друг другу (имеющие боковое расположение листья, многие цветки). Способность к регенерации огромна: каждая точка роста может, в принципе, вырасти в полное новое растение, на чем основано часто применяемое в садоводстве и вообще в сельском хозяйстве вегетативное размножение черенками, отводками, выводковыми почками и т.д. К тому же в хаотически растущих клетках (каллусная ткань), которые обычно образуются сначала после повреждения, могут заново возникать точки роста. Благодаря этому из культур клеток растений могут снова регенерировать целые растения, что у животных в культуре клеток и тканей невозможно. Растения, возраст которых насчитывает несколько столетий и даже тысячелетий, нередко встречаются не только среди деревьев и кустарников, но и среди травянистых многолетников.
Животные и растения значительно различаются также по строению и функции своих клеток. Общее, основанное на типичных чертах противопоставление делает ясным, что растительные клетки (фитоциты) отличаются не только наличием пластид. Они не только фототрофны, но и также осмотрофны, т.е. воспринимают вещества лишь в растворенной форме, тогда как животные клетки (зооциты) фаготрофны, т.е. могут воспринимать питательные вещества в форме частиц. Характерно, что у жгутиковых имеется так называемое миксотрофное питание, т.е. развиты обе формы клеточного питания (рис. 4). Растительная клетка обладает в зрелом состоянии центральной вакуолью, которая часто составляет свыше 90% объема клетки, и прочной клеточной стенкой. Клеточная стенка механически сопротивляется гидростатическому давлению вакуоли (тургор), которое иначе разорвало бы клетку на куски. Тургор — следствие осмотических явлений; общая молярная концентрация клеточного сока в вакуоли значительно выше, чем водных растворов в клеточных стенках. Клетки тканей животных не имеют ни крупных вакуолей (и поэтому обычно значительно мельче, чем клетки тканей растений), ни жестких клеточных стенок, которые служат для стабилизации отдельных клеток. Их тургор незначителен, так как они заполнены изотоническими жидкостями тела и ткани. Массивные межклеточные вещества соединительной и опорной ткани у животных служат для упрочения не клеток, а надклеточных структур. При клеточном делении первый зачаток стенки у растительных и грибных клеток возникает между дочерними клетками путем «внутренней» секреции веществ клеточной стенки. Напротив, у животных типичная форма клеточного деления — перешнуровка материнской клетки (дробление). И если клетки тела растения почти без исключения остаются фиксированными на месте своего образования, при развитии зародышей животных бывает перемещение и миграция клеток.
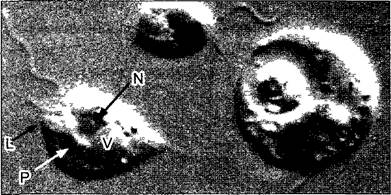
Рис. 4. Potenoochromonas таИпатепая (интерференционный контраст, микрофотоснимок w. Негth)
Это миксотрофный жгутиковый организм из порядка Сhrysomonadales (ср рис 11 74) с двумя жгутиками неравной длины и питающими ложноножками на переднем конце (лобоподий L), а также с задним придатком для прикрепления (1160х). В клетке слева можно различить ядро (N) с ядрышком, пластиду Р и запасающую вакуоль V. У клетки справа видна крупная пищеварительная вакуоль с наполовину переваренной водорослевой клеткой внутри
Клетки грибов — если не считать отсутствия пластид и неспособности к фототрофности — близки к типичным растительным клеткам: здесь мы имеем вакуолизированные, осмотрофные клетки с жесткими клеточными стенками, которые обычно делятся не путем дробления, а путем внутренней секреции клеточных стенок.
Подразделение и значение ботаники
Изучение мира растений, грибов и протистов может (как и вообще в области всех организмов) рассматриваться с совершенно разных точек зрения. Можно, например, весь спектр работ в области биологии рассматривать с позиций иерархии исследуемых структур (табл. 1). Фундаментальные исследования направлены при этом на понимание формы и функции в их обоюдной зависимости, осуществлении и многообразии. При этом на переднем плане находится объект исследования. В прикладных исследованиях, напротив, имеют дело с применением растений, грибов, микроорганизмов для питания человека и полезных животных; с лекарственными, ядовитыми и наркотическими растениями — основа фармакологии; с выращиванием, генетическими манипуляциями и биотехнологией; с сельским хозяйством в самом широком смысле и лесоводством; с фитопатологией, борьбой с вредителями, патогенами и сорной растительностью; с уходом за ландшафтами, охраной природы и отдельных видов и с экологией в современном понимании. Фундаментальные исследования дают базовые знания для прикладного исследования любого рода.
Таблица 1. Направления биологических исследований и уровень организации объектов (по J. Lengeler)
Структуры |
Направления исследования |
Атомы |
Биофизика |
Молекулы |
Биохимия |
Носители информации (семантические макромолекулы) |
Молекулярная биология |
Гены, хромосомы |
Генетика |
Клетки |
Биология клетки |
Ткани |
Гистология |
Органы |
Анатомия, физиология |
Организмы |
Морфология, физиология развития, систематика, филогения, аутэкология |
Популяции |
Геоботаника, синэкология |
В данном учебнике вначале дается описание общих закономерностей структурной организации. При этом затрагивается огромная область от атомарного до макроскопического уровня: после обзора молекулярных основ рассматриваются строение и ультраструктура клетки (цитология), далее ткани растений (гистология) и, наконец, их внешнее, различимое невооруженным глазом строение (морфология). При этом мы исходим прежде всего не из разнообразия форм, а из общих исходных положений, причем на переднем плане стоит описание.
Представление о структурах дается в учебнике в каждом случае с освещением общих функций в обмене веществ и энергии, смен форм и движений. В области физиологии на переднем плане стоит динамика жизненных процессов. За подробным рассмотрением физиологии обмена веществ следуют физиология развития и, наконец, физиология движения. Заключительный, во многих отношениях особенно актуальный раздел посвящен аллелофизиологии, т. е. всевозможным физиологическим отношениям между разнообразными организмами в природе.
При делении учебника «Ботаника» на части и разделы авторы не оставили без внимания того факта, что современная биология отличается междисциплинарным характером. Некогда отдельные области знания сливаются между собой и дают новые, особенно плодотворные области исследования. Так, например, из срастания описательного учения о клетке (цитологии), биохимии и молекулярной биологии возникла в результате современная клеточная биология.
В материале учебника, где рассматриваются эволюционное учение, закономерности и причины видообразования и их генетические основы, значительное место занимает ботаническая систематика. Как наука о родственных связях организмов она основывается на данных всех других дисциплин и имеет дело с описанием, названиями и упорядочением свыше 500 000 видов ныне известных растений. Целенаправленное упорядочение ориентировано на реконструированное эволюционное развитие (филогению) растительного царства. При этом сравнения последовательностей нуклеиновых кислот и белков (молекулярная филогения) и исследования ископаемых растительных остатков (палеоботаника) играют доминирующую роль. Систематическая часть содержит сведения о направлениях, в области которых ведутся интенсивные исследования с отдельными группами организмов (микробиология и бактериология, микология и т.д.), а также о прикладных дисциплинах, которые исследуют практическое значение растений для человека.
Экология растений, наконец, занимается взаимоотношениями между растениями и целыми растительными сообществами с окружающей их биотической и абиотической средой. Экологическая ботаника пытается понять факторы, закономерности и причины распространения и совместного существования растений на Земле в пространстве и времени. Из-за особого значения, которое придается природным экосистемам в современном, перенаселенном человеком, мире, этой важной области исследований и соприкасающимся с ней областям знаний (таких как изучение биотических взаимодействий, учение о растительности, изучение глобальных изменений) посвящена последняя часть учебника.
С экологией растений сопряжен аспект, убедительно показывающий особое значение ботаники непосредственно в современном мире. В энергетическом плане вся жизнь на Земле зависит от фототрофных организмов и при этом практически полностью от растений: они находятся как единственные реальные продуценты в начале практически всех пищевых цепей и составляют фундамент всех пищевых пирамид. Это имеет место, по крайней мере, в течение миллиарда лет. Не в последнюю очередь благодаря большому разнообразию (биоразнообразию) растения обеспечивают поддержание структуры и функции крупных экосистем. В настоящее время это многообразие, как и функции отдельных особей, находятся с самых разных сторон под угрозой из-за слишком ощутимого влияния на биосферу более 6 млрд человек. При этом именно человек относится к числу организмов, которые для своего выживания как особи и вида особенно нуждаются в стабильной окружающей среде. Защита окружающей среды, основанная на максимально возможном научном понимании, в таких условиях важнее, тем более что диктуется она не только соображениями этики ответственности за все живое на Земле, но и по крайней мере с позиций эгоизма — в противном случае мы подрываем основы нашей собственной жизни. Очевидно, что эта сложная тема легче может быть узурпирована чуждыми реальности утопистами и эмоциональными демагогами, если серьезная наука меньше сможет вносить свой вклад в решение экологических проблем.
В развитии биологических наук ботаника, как и раньше, играет важную роль. Многие фундаментальные биологические положения были обоснованы прежде всего при изучении растений. Так, на них были сделаны открытия клетки и клеточного ядра, хромосом, митоза и мейоза, осмоса и законов наследственности. И если даже в настоящее время для решения многочисленных проблем современной биологии наиболее подходящие системы можно найти среди микроорганизмов и определенных представителей животного мира, а многие важные вопросы рака, иммунной системы, памяти и сознания, естественно, могут разрабатываться только на (высших) животных, ботаника тем не менее остается плодотворной областью изучения основ биологии. Снова и снова чрезвычайно важны прикладные исследования в этой области. В биотехнологии растения и грибы неуклонно играют центральную роль. Неудивительно, что и генная инженерия в сельском хозяйстве получает быстро возрастающее значение («зеленая генная инженерия»). При этом, как и повсюду в современной биологии, расшифровка генома (геномика) все больше пополняется исследованием различных белковых структур неодинаковых клеток одного и того же организма (протеомика).
Наконец, и в настоящее время не надо забывать о том, что растения, эти тихие и часто особенно красивые объекты ботаники, вновь и вновь оказывают на человека сильное эмоциональное воздействие. Если ботаника в прежние времена носила прозвище «scientia amabilis» (любительская наука), то этим она обязана эстетической привлекательности деревьев и цветов, что находится уже за гранью естественной науки, однако дорого для человека.
Учение об облике в биологии — морфология — занимается всеми характерными для живых организмов структурами — от биомолекул до крупнейших организмов. Как наиболее хорошо известный пример здесь — клеточное строение листочка листостебельного мха (Plagiochila asplenioides, внизу); во всех клетках многочисленные хлоропласты и светлые масляные тельца в виде виноградных гроздей, содержащие различные терпены. Вверху — внешний вид «розы» цветной капусты (Brassica olerácea var. botrytis). Речь идет о структуре, представляющей собой обширное соцветие, у которого вытягивание элементов осей сначала подавляется (впрочем, у изображенного здесь сорта «Minaret» не совсем). Эта совокупность почек образует «фрактальную» структуру, у которой формообразование основного побега повторяется у всех боковых побегов, у их боковых побегов снова и т. д. Только на этом фрагменте иллюстрации при соответствующем увеличении было бы видно почти 500 тыс. точек роста. Оба рисунка позволяют различить образования структур, как они снова и снова встречаются в бесконечных вариациях у самых различных живых организмов
