БОТАНИКА ТОМ 2 - ФИЗИОЛОГИЯ РАСТЕНИЙ - 2007
6. ФИЗИОЛОГИЯ ОБМЕНА ВЕЩЕСТВ
6.12. Мобилизация запасных липидов
Во время прорастания запасающих жиры семян расщепляются накопленные в олеосомах нейтральные жиры, а углерод используется в синтезе углеводов, за счет которых затем будут покрываться издержки ана- и катаболизма проростка, пока он питается гетеротрофным способом. В реакциях метаболизма принимают участие цитоплазма, глиоксисомы и митохондрии. Глиоксисомы, родственные пероксисомам органеллы (см. 6.5.6), в огромных количествах встречающиеся в запасающих клетках во время процесса мобилизации запасных липидов; они исчезают на свету с началом фотосинтеза и замещаются перо- ксисомами, которые с этого времени, помимо всего прочего, задействованы в фотодыхании (см. 6.5.6). Новые исследования показали, что пероксисомы происходят из глиоксисом на свету в результате изменения ферментного состава.
Мобилизация запасных липидов начинается с гидролитического высвобождения жирных кислот из триглицеридов, которое катализируется липазами. Появившийся при этом глицерин сначала превращается под действием глицерин-3-киназы с использованием АТФ в глицерин-3-фосфат, который затем преобразуется глицерин-3-фосфатдегидрогеназой в дигидроксиацетонфосфат (см. рис. 6.101). Этот триозофосфат включается в цитоплазматический обмен сахаров.
Жирные кислоты высвобождаются в цитоплазму, а затем поступают в глиоксисомы (предположительно, посредством диффузии сквозь образованные поринами
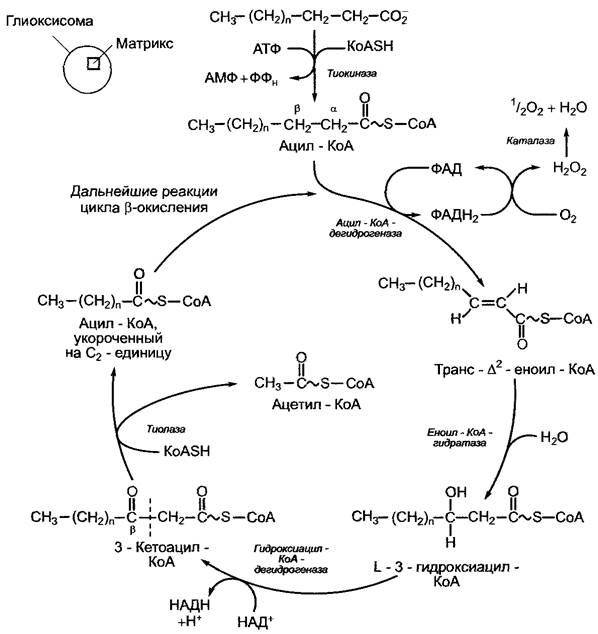
поры, проницаемые для низкомолекулярных метаболитов, см. 6.5.6) и в процессе β-окисления превращаются там в ацетил- КоА. В отличие от животных растения не обладают способностью к митохондриальному β-окислению, оно протекает исключительно в глиоксисомах или пероксисо- мах. Последовательность реакций этого окисления (рис. 6.103) очень похожа на митохондриальную, с той лишь разницей, что в митохондриях связанная с ацил-КоА- дегидрогеназой ФАД, которая в процессе реакции восстанавливается до ФАДН2, окисляется не в дыхательной цепи (см. 6.10.3.3, рис. 6.96), а молекулярным кислородом. Продукт реакции, Н2O2, очень агрессивен химически и вступает в реакцию диспропорционирования под действием каталазы, которая в больших количествах имеется в глиоксисомах и пероксисомах: Н2O + 1/2O2.
Рис. 6.103. Процесс β-окисления насыщенных жирных кислот в глиоксисомах. Полное β-окисление ненасыщенных жирных кислот требует нескольких дополнительных ферментативных реакций, на которых мы здесь останавливаться не будем
Дальнейшая судьба ацетил-КоА представлена на рис. 6.104. В глиоксилатном цикле (рис. 6.105) формально в сукцинат конденсируются две ацетатные единицы из ацетил-КоА. Такая последовательность реакций обнаружена, кроме зеленых растений, у грибов и бактерий (которые, таким образом, могут развиваться на ацетате, используя его в качестве источника углерода), но не у животных. Отметим, что глиоксисомы присутствуют только в клетках эукариот.
Рис. 6.104. Схема реакций преобразования нейтральных жиров (= триглицеридов) в гексозы и принимающие в них участие компартменты. Фосфоенолпируваткарбоксикиназная (ФЕП-карбоксикиназная) реакция подробно представлена на рис. 6.79
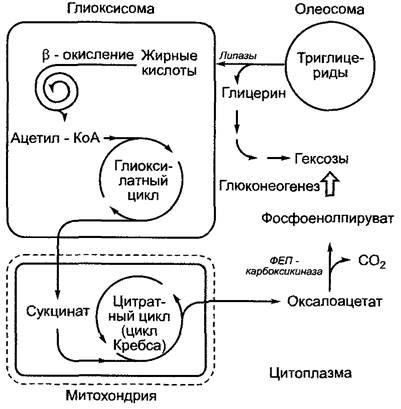
Рис. 6.105. Последовательность реакций глиоксилатного цикла. На сером фоне — основные ферменты глиоксисом
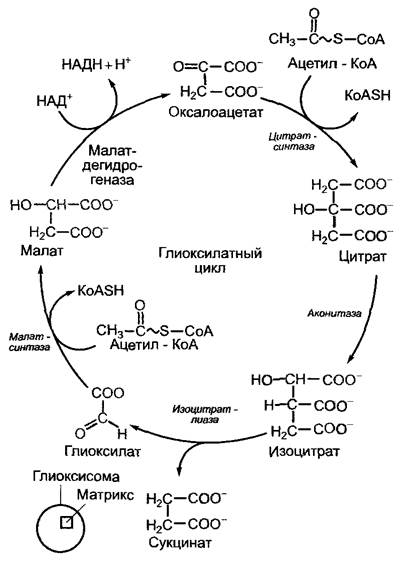
Характерными ферментами глиоксилатного цикла являются изоцитратлиаза и малатсинтаза, остальные ферменты нам знакомы по цитратному циклу (см. рис. 6.94). В процессе изучения культуры клеток огурца обнаружено, что недостаток глюкозы активирует гены ферментов глиоксилатного цикла. С помощью подобного механизма регулируется превращение жиров в углеводы в соответствии с потребностью растения.
Образованный в глиоксилатном цикле сукцинат покидает глиоксисомы через порины и в результате некоторых реакций нитратного цикла преобразуется в митохондриях в оксалоацетат, который в свою очередь в цитоплазме преобразуется фосфоенолпируваткарбоксикиназой в фосфоенолпируват. Из этого метаболита в результате реакций глюконеогенеза (см. 6.10.3.4, рис. 6.98) образуется гексоза. В итоге описанной последовательности реакций 75 % углерода любой жирной кислоты (три из четырех С-атомов) теоретически может преобразоваться в гексозы. остаток (один из четырех С-атомов) теряется в процессе фосфоенолпируваткарбоксикиназной реакции в виде СO2.
С помощью опытов по мечению жирных кислот радиоактивными изотопами 14С было показано, что теоретический высший показатель преобразования жирных кислот в углероды достигается и в эксперименте. Благодаря эффективной компартментации растению почти полностью удается избежать потерь углерода в побочных реакциях.
Помимо того, что реакции преобразования жиров в углеводы обнаружены при прорастании семян, запасающих жиры, аналогичные биохимические процессы наблюдаются при старении листьев (при превращении мембранных липидов в углеводы в целях их оттока в ствол) и весной в стволах деревьев при снабжении углеводами побегов (о преобразовании запасенных осенью липидов в углеводы и их загрузки в ксилему см. 6.3.5).