БОТАНИКА ТОМ 2 - ФИЗИОЛОГИЯ РАСТЕНИЙ - 2007
7. ФИЗИОЛОГИЯ РАЗВИТИЯ
Под развитием понимают совокупность всех процессов изменения формы и функций в жизненном цикле одно- или многоклеточного организма. Процессы развития могут происходить на уровне молекул, ком- партментов, клеток, тканей или органов, они неразрывно связаны с наличием рассмотренного в гл. 6 обмена веществ. Физиология развития занимается анализом механизмов, управляющих развитием. Ее цель — понять молекулярные процессы, благодаря которым при взаимодействии нуклеиновых кислот, белков и низкомолекулярных соединений, а также окружающей среды генетическая информация (генотип) реализуется при развитии организма с определенным набором признаков (фенотип). Это приводит в потомстве как к точному воспроизведению характерных свойств вида родительского поколения, так и к индивидуальной вариабельности в выражении фенотипа в рамках определенной генотипом нормы реакции. Такая пластичность развития служит у растений прежде всего для адаптации организма к различным условиям окружающей среды. Примеры циклов развития растений из различных систематических групп представлены в разделе 11.2.
7.1. Основные принципы физиологии развития
Развитие организма включает процессы роста и дифференцировки. Под ростом понимают необратимое увеличение объема (размеров), под дифференцировкой — качественное изменение формы или соответственно функции клетки, ткани или органа.
Преимущественно как процесс роста будем обозначать, например, развитие картофельного клубня от набухания апикальной части столона до достижения клубнем окончательной величины, увеличение длины колеоптиля, которое происходит при постоянном числе клеток исключительно благодаря их растяжению, или же размножение ткани в клеточной культуре in vitro. Преимущественно дифференцировка — это, например, превращение клетки эпидермиса в замыкающую клетку (см. рис. 3.13) или превращение прокамбиальных тяжей в различные элементы проводящего пучка (см. 3.2.4, рис. 3.22, F — L).
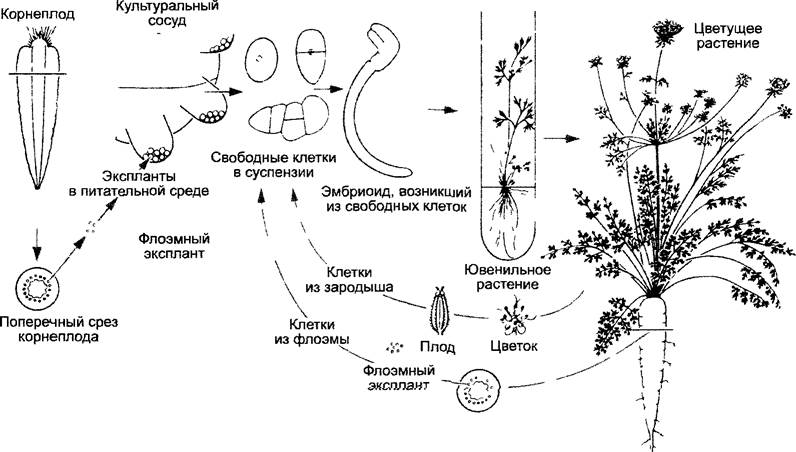
Регенерация растений из культуры клеток (рис. 7.1) дает возможность экспериментально разделить в значительной мере процессы роста и дифференцировки. Однако рост и дифференцировка, как правило, протекают друг за другом. Раздельное рассмотрение имеет прежде всего дидактические основания. В то же время возможность регенерировать из дифференцированной клетки целое растение (например, из клетки мезофилла или клетки сердцевины побега) доказывает тотипотентпость живых растительных клеток, т.е. наличие в них полной генетической информации и после окончания дифференцировки клеток, поскольку присутствуют все органеллы, содержащие наследственную информацию (см. 7.2.1). У ситовидных клеток в ходе дифференцировки теряется клеточное ядро, поэтому регенерация из таких клеток в принципе исключена.
Рис. 7.1. Развитие полноценного растительного организма из изолированных клеток на примере Daucus carota. Отдельные клетки из флоэмных эксплантатов, а также из незрелых зародышей развиваются через стадию эмбриоидов (называемых также соматическими зародышами) до молодых растений, которые вырастают высокими, цветущими и плодоносящими. Рост в виде клеточной суспензии и регенерация целых растений из отдельных клеток в культуре in vitro регулируются составом фитогормонов в питательной среде (см. 7.6.2.3). Аналогичным способом можно вегетативно размножать многие виды
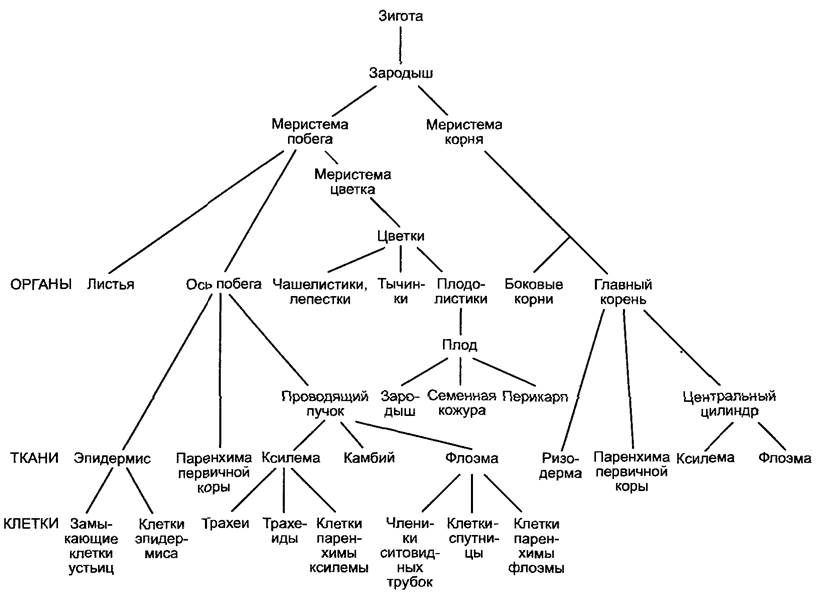
В ходе онтогенеза многоклеточный организм проходит различные уровни дифференцировки, представленные на рис. 7.2 на примере развития цветкового растения.
Во время эмбриогенеза кроме оси зародыша (гипокотиль и зародышевый корешок) и 1 — 2 зародышевых листьев закладываются обе первичные меристемы (побеговая и корневая), из которых развиваются все другие органы растения. Это событие в развитии организма можно воспринимать как иерархический процесс, в ходе которого сначала определяются тип и положение меристемы1, затем внутри меристемы определяются тип, число и положение органов и, наконец, внутри размеченных областей органообразования (например, в зачатке листа) — число, расположение и дифференцировка клеток, ведущие к образованию ткани и определяющие форму и величину органа.
1 В иностранной литературе пользуются термином «идентичность», который подразумевает определение типа органа, его места и анатомической структуры. В переводе мы даем его расшифровку, поскольку в русском языке «идентичность» имеет другой смысловой оттенок. — Примеч. ред.
Рис. 7.2. Стадии дифференцировки во время развития высшего растения
Уже первое деление зиготы неравное и дает две клетки: базальную клетку, из которой образуются суспензор, покоящийся центр и часть корневого чехлика, и апикальную клетку, из которой образуется остальной зародыш. Полярность зиготы встречается уже у низших растений, однако клеточную полярность можно найти и у других типов клеток (см. 7.3.3). В качестве причин, вызывающих поляризацию клеток, рассматривают градиенты веществ. Они в то же время важны для возникновения позиционной информации в многоклеточной системе (см. 7.4). Определение судьбы при дифференцировке (детерминация) растительной клетки лишь отчасти регулируется автономными внутриклеточными процессами (исключительно определяемыми геномом), например, меристемные инициали в заметной мере детерминируют дальнейший путь дифференцировки произведенных ими дочерних клеток (в соответствии с моделью дифференцировки клеток: англ. cell-lineage model)1. Однако позиция клетки в многоклеточном организме также оказывает значительное влияние на ее дальнейшую судьбу, т.е. дифференцировка клетки в значительной степени контролируется ее окружением. Таким образом, многоклеточный организм развивается не только благодаря сочетанию множества отдельных автономных внутриклеточных процессов (из-за малых возможностей исправлять ошибки, это легко бы привело к нестабильности программы развития в целом), а в виде комплекса взаимодействующих друг с другом клеток (чаще объединенных в симпластические системы), которые взаимно координируют и контролируют свою активность.
1 Речь идет о детерминированном развитии организма, когда судьба клеток строго определена генетической программой развития. Этот тип развития характерен для круглых червей (например, Саenorhabditis). Другим организмам, в том числе растениям, больше присуще регулятивное развитие, позволяющее исправлять случайные ошибки. — Примеч. ред.
Градиенты веществ оказывают влияние не только на ход дифференцировки отдельных клеток или соответственно клеток в клеточном комплексе, они также ответственны за разметку1. Под этим понимают процессы, которые определяют число, положение дифференцирующихся клеток и дальнейшего развития. Наилучшим русским эквивалентом можно считать термин «разметка». Химическая природа действующих веществ, принимающих участие в образовании градиентов, важных для развития, известна лишь в редких случаях (см. 7.4.2). Однако здесь могли бы участвовать фитогормоны (см. 7.6), особенно ауксин индолил-3-уксусная кислота (см. 7.6.1).
1 Здесь использован термин Musterbildung (калька с англ. pattern formation), что дословно можно перевести как «формирование образа», «создание шаблона» для
временной интервал событий дифференцировки. Так, размещение устьиц в эпидермисе, образование трихомов, корневых волосков и боковых корней, число и положение листьев на оси побега контролируются процессом разметки.
В отличие от большинства животных высокоразвитые растения построены по модульному принципу. Модули называются также фитомерами. Они образуются апикальными меристемами, т. е. меристемами побега и меристемами корня. Единица организации побега состоит из узла, междоузлия, пазушной почки и листа; единица организации корня — из участка оси корня и зачатков боковых корней. Развивающиеся боковые корни и пазушные почки снова образуют в свою очередь фитомеры и т.д. Хотя строение побега показывает, что термин индивидуум (лат. — неделимое) для растений, строго говоря, не подходит, фитомеры все-таки не полностью самостоятельные единицы развития растения, а сами находятся под строгим контролем целого организма. Этот процесс системного контроля развития называется корреляцией (см. 7.5). Так, верхушечная почка в апикальной области побега ингибирует прорастание пазушных почек (апикальное доминирование — см. 7.5; 7.6.1.4). Системный контроль можно обнаружить также при индукции образования цветка, развитии плодов и образовании запасающих органов.
Даже у отдельной клетки рост — сложный процесс; в многоклеточном организме, кроме того, рост каждой отдельной клетки должен быть согласован с ростом соседних и всех остальных клеток организма во времени и пространстве, что делает процесс еще сложнее.
Клеточный рост охватывает, с одной стороны, увеличение количества составных частей клетки (рост цитоплазмы), что может происходить без значительного увеличения объема клетки (например, при росте меристемных клеток между клеточными делениями — см. рис. 3.5), и, с другой стороны, рост растяжением, идущий часто со значительным увеличением объема клетки, уже воспринимаемый как процесс дифференцировки, при котором клетка либо растягивается во всех направлениях более или менее равномерно (изодиаметрический рост, например, у многих паренхимных клеток — см. рис. 3.7, А), либо расширяется в определенных предпочтительных направлениях (прозенхиматический рост). Например, в колеоптилях трав в ситовидных клетках или склеренхимных волокнах прозенхиматический рост приводит к образованию сильно удлиненных клеток (см. рис. 3.20). Увеличение объема при растяжении клеток вызывается преимущественно поглощением воды. Поэтому рост растяжением всегда сопряжен с увеличением вакуолей и образованием центральной вакуоли; общее количество белков при растяжении клетки не должно увеличиваться. При растяжении клетки материал клеточной стенки синтезируется умеренно: у стебелька коробочки печеночного мха Lophocolеа (лофоколея), например, в течение 3 — 4 дней увеличение массы клеточной стенки происходит лишь в 1,8 раза на фоне 48-кратного удлинения клетки.
Рост растяжением может охватывать всю поверхность клетки более или менее равномерно, или быть ограниченным определенным участком клеточной стенки. Выраженный верхушечный рост демонстрируют, например, апикальные клетки некоторых водорослей, а также грибных гиф, корневых волосков, пыльцевых трубок и некоторых растянутых, прозенхиматических клеток в тяжах прокамбия. Неравномерный сильный рост в нескольких местах поверхности клетки — основа для образования более сложных клеточных форм (например, в случае клеток губчатой и звездчатой паренхимы, некоторых идиобластов и волосков, отдельных клеток у водоросли Мicrasterias, см. рис. 3.7, В, С).
Рост многоклеточных включает наряду с клеточным ростом также размножение клеток (рост делением). В некоторых органах, например, в корнях, зона деления (ме- ристематическая, эмбриональная зона) четко ограничена от зоны роста растяжением, в то время как в апексе побега они постепенно переходят друг в друга. За окончанием растяжения клетки обычно следуют дальнейшие процессы дифференцировки.
Таблица 7.1. Продолжительность и скорость роста некоторых органов растений путем растяжения
Орган |
Продолжительность растяжения |
Скорость растяжения |
Зародышевый корень бобов |
3 сут |
0,012 мм мин 1 = 1,7 см сут-1 |
Колеоптиль овса |
2 сут |
0,025 мм мин 1 = 3,6 см сут-1 |
Побег бамбука |
Несколько суток |
0,4 мм мин 5 = 58 см сут-1 |
Тычинки ржи |
10 мин |
2,5 мм мин 5 |
Плодовое тело гриба (Dictyophora) |
15 мин |
5 мм мин 1 |
Часто рост частей растений объясняется исключительно растяжением клетки без участия клеточных делений. Это справедливо, например, для
✵ роста колеоптилей злаков,
✵ раскрывания почек и цветения многих деревьев в течение нескольких дней весной;
✵ первой фазы роста зародышевых корешков;
✵ быстрого растяжения некоторых побегов (например, бамбук);
✵ удлинения тычинок (например, у злаков);
✵ вытягивания стебелька коробочки (seta) у спорогониев мха;
✵ ножек плодовых тел базидиомицетов.
Скорость растяжения органов при этом часто значительна (табл 7 1). У почвенных корней зона роста растяжением находится прямо за кончиком и насчитывает в длину лишь несколько миллиметров (рис 7 3). Верхушечная меристема корня у кукурузы образует в день около 10 000 клеток корневого чехлика и тем самым ежедневно полностью обновляет корневой чехлик, а также около 170000 клеток для роста корня в длину. В зоне образования корневых волосков клетки чаще всего уже достигают максимального размера и начинают окончательную дифференцировку. У воздушных корней зона роста растяжением длиннее, и еще значительно более длинная она у побега. Например, у спаржи Asparagus officinalis она превышает 50 см. У осей побега, которые подразделяются на узлы и междоузлия, дольше всего способным к росту остается основание междоузлия.
У злаков этот интеркалярный рост сохраняется долгое время, причем в участках междоузлий над узлами наряду с ростом растяжением обнаружен рост цитоплазмы и рост делением. Подобные базальные интеркалярные зоны роста образуются также на листьях (особенно четко у хвойных и однодольных, но также у двудольных растений). Так, например, черешок листа интеркалярно растет между листовой пластинкой и основанием листа.
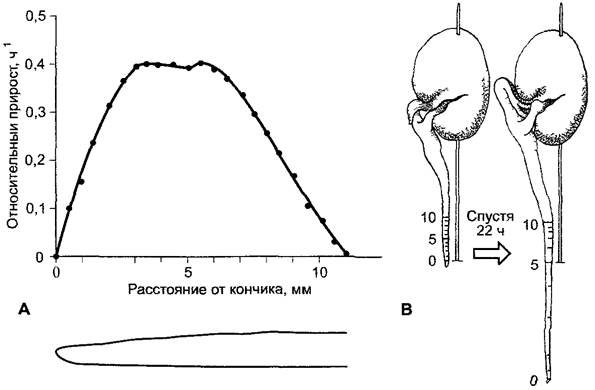
Если наблюдать за локальной скоростью роста вдоль зоны роста, примерно в зоне растяжения корня (рис. 7.3, А), то можно заметить постепенное увеличение скорости роста до максимума и далее спад до остановки («большой период роста»). Такого рода увеличение и спад скорости роста демонстрирует каждая отдельная клетка, которая «проходит» зону растяжения. Подача клеток из меристемы и начало их роста растяжением так гармонично связаны со спадом интенсивности роста в более старых частях органа, что корни в целом растут дальше равномерно. У побегов часто встречается циркадное1 нарастание и спад скорости роста: в темноте скорость роста немного выше, чем в течение дня. Этот процесс управляется светом (см. 7.7.2). Периодичный рост может (например, у Роасеае) осуществляться, если молодое междоузлие только начинает рост растяжением, тогда как более старое в значительной мере его заканчивает. У ячменя было выявлено периодическое нарастание концентрации фитогормона гиббереллина GA1, стимулирующего растяжение междоузлий (см. 7.6.3): его концентрация возрастает каждый раз перед повышением скорости роста междоузлия (о механизмах роста цитоплазмы см. 7.3.1, роста делением см. 7.3.2, растяжения клеток см. 7.3.3; 7.6.1.4).
1 С периодом 24 ч — Примеч. ред.
Рис. 7.3. Рост корня. А — распределение скоростей роста вдоль первичного корня проростка кукурузы; был установлен относительный прирост в длину в 1 ч (0,1 = 10 %) из кратковременных измерений в различных местах корня, В — распределение прироста в кончике корня Vicia faba; штрихи обозначают положение отметок тушью, которые были нанесены на корне в начале эксперимента на расстоянии 1 мм друг от друга (слева), спустя 22 ч (справа) Штрихи тушью из-за неравномерного роста отдельных зон оказываются на разных расстояниях друг от друга
Дифференцировка и коррелятивный контроль при развитии рассмотрены далее (см. 7.3 — 7.5), а сейчас перейдем к генетическим основам развития.