БОТАНИКА ТОМ 2 - ФИЗИОЛОГИЯ РАСТЕНИЙ - 2007
9. АЛЛЕЛОФИЗИОЛОГИЯ
9.5. Аллелопатия
Аллелопатия — это химическое влияние на одно растение со стороны другого растения. При этом речь может идти о стимулирующем или, чаще, ингибирующем действии. Влияние может происходить, с одной стороны, благодаря летучим соединениям, которые в диффузионной или конвекционной камере достигают концентраций, достаточных для того, чтобы стать активными. Так, этилен обладает характером как феромона (сигнальные вещества, которые имеют координирующие задачи
между индивидуумами одного вида), так и кайромона (сигнальные вещества, которые действуют между индивидуумами различных видов). Однако достигается ли в природе достаточно высокая концентрация этилена в воздушном пространстве, это пока не выяснено. Правда, при транспортировке и хранении плодов (особенно, если совместно содержатся сорта/виды с различной чувствительностью) нужно обращать внимание на ускорение старения благодаря высвобождающемуся этилену (см.
7.6.5.2). Предметом дискуссии может служить также вопрос, оказывает ли наблюдаемое в эксперименте высвобождение алармонов растениями, пораженными насекомыми (см. 9.4.2), сигнальное влияние на соседние, еще не пораженные растения для активации их защиты от травоядных. Кроме того, многие растения выделяют в почву через корневую систему водорастворимые соединения, которые могут ограничить рост конкурентов. Наконец, у некоторых видов активные соединения аллелопатически вымываются с осадками, которые попадают в почву со стоком.
Хотя считается установленным, что аллелопатически действующие ингибиторы часто играют роль в растительных сообществах, характеризующихся наличием конкуренции, однозначное доказательство провести трудно, так как на эти эффекты наслаиваются и другие (например, конкуренция за свет и питательные вещества). К тому же многие соединения, которые рассматривались как аллелопатики, вызывают очень неспецифичное ингибирование и у микроорганизмов и животных (по крайней мере, в эксперименте). Сюда относятся многие простые фенолы (например, коричная кислота и ее производные, такие, как кумарин, см. 6.16.1) и терпеноиды (см. 6.16.2). Этими соединениями заметно ингибируется, в первую очередь, прорастание некоторых видов, поэтому кажется правдоподобным, что при их накоплении в верхних слоях почвы (особенно благодаря вымыванию из листьев или разрушению пленчатых чешуй) сокращается внутри- или, соответственно, межвидовая конкуренция из-за нового нарастания уже сформировавшихся индивидуумов.
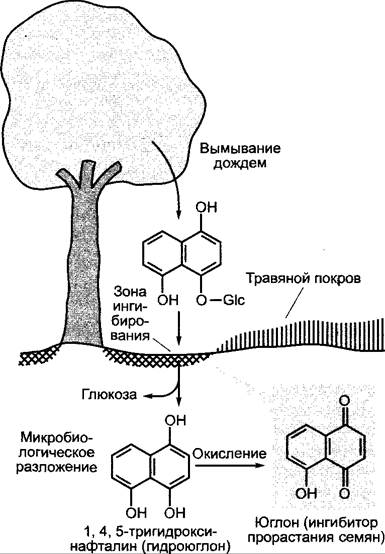
Так, из листьев и плодов грецкого ореха (Juglans regia) вымывается 1,4,5-тригидроксинафтил-4-глюкозид и переводится в почве путем гидролиза и последующего окисления в сильный ингибитор прорастания юглон (рис. 9.23). Тем самым дополнительно к фактору света (затенение кроной) и питательных веществ (истощение почвы в области корней) создаются неблагоприятные условия для прорастания семян, так что на ширине кроны дерева обнаруживают лишь слабый травянистый покров.
Рис. 9.23. Аллелопатическое влияние юглона грецкого ореха (Juglans regia). Юглон блокирует пропилпептидилизомеразы, которые необходимы для приобретения правильной конформации белками клеточного цикла, и прерывает тем самым клеточный цикл в фазе G2. По-видимому, именно этим можно объяснить прямое токсическое действие юглона на сильно растущие проростки
Четкий пример аллелопатии можно увидеть в южно-калифорнийских кустарниковых сообществах в долине Санта-Инес (см. 15.2.7), называемых чапарралем. Растительность здесь представлена густым травостоем Salvia leucophylla и Artemisia califomica, которые все больше оттесняют травянистые растения. В зоне шириной около 1-2 м вокруг кустарников нет никаких других растений, в области 3-8 м — замедленный рост и лишь на большем расстоянии есть неподавленные растущие растения, в первую очередь травы (например, Bromus hordeaceus, Festuca megalura и Avena fatua). Подавление роста трав объясняется исключительно токсичными для травянистых растений монотерпенами, прежде всего камфарой и 1,8-цинеолом (см. рис. 6.123), которые выделяются Salvia leucophylla и Artemisia califomica особенно при высоких температурах воздуха. Эти монотерпены накапливаются в верхних слоях почвы благодаря адсорбции на почвенных коллоидах суглинистых почв. Оттуда очень
липофильные соединения (равновесие распределения через газовую фазу!) переходят в клеточные мембраны прорастающих семян. Аналогично происходит накопление монотерпенов в липофильных кутикулах трав, откуда должен происходить переход в растительные клеточные мембраны. Неясен механизм токсичности камфары и 1,8- цинеола, так же, как и защитный механизм Salvia или Artemisia. Накопление монотерпенов в густых травостоях кустарниковых сообществ повышает при высоких температурах вероятность спонтанного самовозгорания смеси воздух—терпен. Чапарралль подвержен с периодичностью примерно в 25 лет возгораниям, в ходе которых травостои Salvia и Artemisia, а также терпены в почве разрушаются огнем. После этого сначала формируется травянистый покров, за ним появляются и распространяются кустарники.