БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
Третье подцарство: Heterokontobionta
Организмы, относящиеся к этой большой группе, отличаются гегероконтными подвижными стадиями, т. е. они снабжены одним направленным вперед перистым жгутиком и одним направленным назад гладким жгутиком, который служит рулем (см. рис. 11.20, А, С; 11.72, F). Перистый жгутик имеет два ряда жестких волосков. Последние состоят из основания, трубчатого ствола и терминальных волосков (одного или нескольких). Волоски, встречающиеся иногда на жгутиках среди представителей других эволюционных групп (например, Glaucobionta, Chlorobionta), никогда не бывают ни жесткими, ни трубчатыми.
В этом подцарстве от одних предков возникли как гетеротрофные грибоподобные организмы (Labyrinthulomycota; Ооmycota), так и автотрофные водоросли (Heterokontophyta). Сначала рассмотрим гетеротрофные группы, которые по типу организации соответствуют грибам (бокс 11.3).
Первый отдел: лабиринтуловые (Labyrinthulomycota)
Сетчатые слизевики, объединяемые в класс Labyrinthulomycetes, имеют еще организацию слизевиков (см. бокс 11.2). Однако из-за гетероконтного типа организации жгутикового аппарата они сближаются с рассматриваемыми далее Oomycota. Сюда относятся виды, которые поражают как эндопаразиты морские растения (например, Zoslera, Laminaria)1. Для них характерны многоклеточные сетчатые плазмодии; последние возникают путем деления двужгутиковых подвижных клеток внутри увеличивающейся в размере слизистой оболочки. Сетчатые слизевики представляют собой исходную группу среди Нeterokontobionta. Сходные с ними организмы были предками рассматриваемых далее гетеротрофных Ооmycota, а также ставших в результате эндоцитобиоза автотрофными Неterokontobionta2.
1 Среди лабиринтуловых известны также паразиты некоторых пресноводных водорослей (например, видов Сladophora и Vaucherа). Известны виды и из наземных биотопов, например, паразиты корней пшеницы. — Примеч. пер.
2 К этому же отделу обычно относят и еще одну очень важную группу — класс траустохитрневые (Thraustochytriomycetes). Внешне их талломы конвергентно сходны с некоторыми хитридиомицетами (например, типа Rhizophydium, см. ниже), что и отразилось в их названии. Клетки у них, как и у лабиринтуловых, окружены слизистым чехлом. Эта эктоплазматическая сеть развита слабее, чем у лабиринтуловых, и напоминает ризомицелий у хитридиевых грибов. Она служит только для обмена веществ между клетками и окружающей средой, а также для прикрепления к субстрату. Размножение — посредством зооспор (изредка апланоспор и амебоидных клеток). Зооспоры образуются в большом количестве (иногда свыше 100) в зооспорангиях. Цикл развития от зооспоры до нового зооспорангия может занимать всего 2 — 3 ч. Поэтому траусто- хитриевые стали одними из самых обильных морских организмов, в частности в планктоне и нейстоне (например, в гипонейстоне их численность может составлять 2 — 3 млн на 1 мл воды), и играют важную роль в питании морских животных-фильтраторов. Есть указания на наличие у траустохитриевых полового процесса, но доказательства отсутствуют. Траустохитриевые — облигатно морские и галофильные сапротрофы, встречающиеся по всему земному шару. После бактерий это наиболее массовые гетеротрофные организмы в морях и соленых внутренних водоемах. Некоторые известны как паразиты ряда морских беспозвоночных и рыб. Их насчитывается свыше 40 видов. Ряд видов хорошо развиваются на пыльце сосны, помещенной в морскую воду, так что их можно «выманить» для дальнейших исследований. — Примеч. пер.
Второй отдел: Оомицеты, или целлюлозные грибы (Ооmycota)
Этот отдел, включающий около 500 видов, хотя имеет определенное сходство с грибами (бокс 11.3), по ряду признаков отличается от всех типичных грибов. Таллом,
реже одноклеточный (Lagenidiales) и чаще всего сифональный, имеет почти всегда стенки, содержащие целлюлозу. Размножение осуществляется путем слияния ♂ гаметангиев с оогониями (гаметангиогамия); первые образуют оплодотворяющие выросты, внедряющиеся в оогонии. После оплодотворения в оогониях развиваются зиготы в числе от одной до нескольких (так называемые «ооспоры»). Возникающие при бесполом размножении зооспоры гетероконтные: они несут направленный вперед перистый жгутик и направленный назад гладкий жгутик, чаще всего несколько более длинный (см. рис. 11.20, В). Кроме того, оомицеты (единственный класс отдела), согласно всем имеющимся данным, диплонты, так что мейоз у них происходит перед образованием гамет (гаметическая смена ядерных фаз). Сплетения гиф и плодовые тела никогда не образуются.
Что касается обмена веществ, то оомицеты помимо химизма клеточной стенки имеют следующие особенности. Белок клеточной стенки содержит гидроксипролин. Биосинтез лизина осуществляется после связывания пирувата и аспартата в дигидродипиколиновую кислоту по пути диаминопимелиновой кислоты (у других грибов через аминадипиновую кислоту). Никотиновая кислота синтезируется не из триптофана (как у животных и других грибов), а из СЗ- соединений. Ферменты, участвующие в синтезе триптофана, особого, нигде больше не встречающегося типа. Молекулярная масса рибосом ной PHК (25S-фракция) иная, чем у других грибов (за исключением слизевиков). Оомицеты практически все бесцветны; из них не удалось выделить никаких пигментов.
Все названные морфологические и химические признаки, а также диплоидный жизненный цикл говорят об обособленности Ооmycotа от хитиновых грибов (Еumycota).
Образ жизни. Виды, обладающие признаками, считающимися исходными, обитают в воде чаше всего как сапротрофы. Эволюционно более подвинутые обитатели наземной среды — паразиты высших растений. Это подразделение на группы в зависимости от образа жизни соответствует двум важнейшим (1,3) порядкам оомицетов.
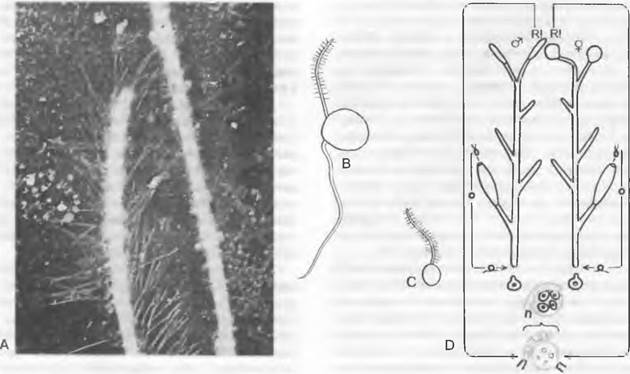
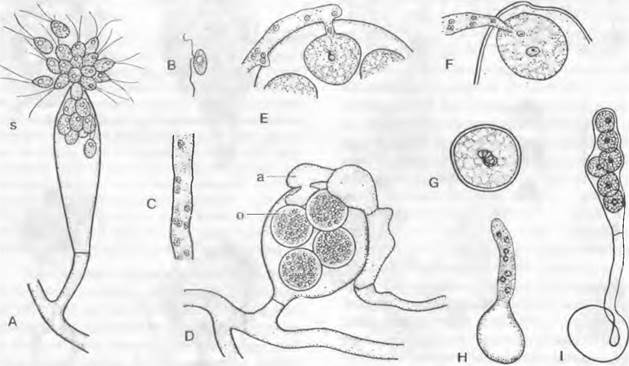
1. Порядок: Saprolegniales. Лишенный поперечных перегородок многоядерный мицелий (см. рис. 11.21, С) обитает в воде, чаще всего в пресной, у некоторых видов также в солоноватой, чаще всего сапротрофно на погруженных туда гниющих частях растений и трупах насекомых; реже он паразитирует на ослабленных живых рыбах. При бесполом размножении концы гиф расширяются в слабо выраженные булавовидные зооспороцисты и отграничиваются перегородкой от несущей гифы. Путем деления плазмы в них возникают одноядерные грушевидные митозооспоры с двумя апикальными жгутиками неравной длины, из которых один несет два ряда волосков (см. рис. 11.21, А; 11.20, А). После плавания в течение некоторого времени жгутики впячиваются. Споры, становящиеся округлыми и одетыми стенкой, образуют на подходящем субстрате проростковую трубку и развиваются в новый организм.
У некоторых оомицетов сначала образуются еще другие зооспоры. Эти вторые подвижные стадии (рис. 11.21. В) отличаются от образуемых первоначально внешним видом и местом прикрепления жгутиков (рис. 11.21, А): они почковидные, и жгутики прикрепляются сбоку (рис. 11.21, В). Это явление дипланетизма отличает род Sapro- legnia от других родов1. У других родов первичные зооспоры встречаются только внутри или же в непосредственной близости от спороцисты (Асhlуа) или вообще не встречаются (Thraustotheca, Dictyuchus)', в последнем случае дипланетизм отсутствует. У Aplanes зооспор нет; споры, инцистирующиеся внутри спороцисты, дают проростковые трубки через ее стенку. Аналогично ведет себя Geolegnia, но с той разницей, что споры высвобождаются из спороцисты до образования проростковой трубки. Для Saprolegnia характерно то, что гифа, несущая спороцисту, после выхода зооспор прорастает новой спороцистой внутрь освободившейся.
1 Более корректно называть дипланетизмом способность зооспор инцистироваться, а потом снова существовать в виде зооспоры; явление изменения морфологии зооспор правильнее называть диморфизмом. — Примеч. пер.
Бокс 11.3. Типы организации грибов
Грибы в более узком смысле слова, как и слизевики, не имеют пластид и хлорофилла; они живут как сапротрофы или паразиты в пресной воде или на суше, реже в морях. Часто их можно культивировать на подходящих питательных средах — в основном это сапротрофные, но также и некоторые паразитные формы; они гетеротрофны не только по отношению к углероду, но и в отношении азота и определенных активных веществ. Эти эукариотные организмы, образующие таллом (слоевище), хотя и рассматриваются традиционно вместе с растениями, занимают особое положение в системе живых организмов. В противоположность слизевикам у них не бывает плазмодия, а галлом чаще всего не является амебоидным, одет клеточной стенкой из хитина, глюканов и т. д. Вегетативное тело редко бывает в виде пузыря или капли, чаще всего оно нитевидное. Отдельные нити гриба называются гифами, а совокупность гиф вне плодовых тел — мицелием. В плодовом теле гифы сплетаются в ложную ткань — плектенхиму.
У грибов можно различить следующие уровни организации:
✵ голые паразитирующие протопласты (например, Olpidium); во всех последующих случаях вегетативная фаза отличается наличием клеточной стенки;
• ризомицелий: содержащий ядро пузырь выпускает в субстрат нитчатые выросты, лишенные ядер (например, Rhizophydium, см. рис. 11.24, А);
• почкующийся мицелий: таллом состоит из каплевидных или несколько вытянутых клеток, которые путем почкования образуют дочерние клетки; в результате их неполного отчленения могут возникать короткие цепочки из клеток (например, у дрожжей; см. рис. 2.34); от этой формы можно вывести
• псендомицелий: здесь возникшие в результате почкования клетки остаются соединенными в разветвленные цепочки (аналогично см. рис. 11.30, G);
• мицелий из гиф и плектенхима плодовых тел: таллом образуется из нитевидных клеток; последние чаще всего разветвленные, у некоторых грибов не разделенные перегородками (сифональные формы; см. рис. 11.28, D), у других они закономерно разделены септами (трихальные формы; см. рис. 11.51, А); гифы часто плотно собраны вместе или сгруппированы в плодовое тело (см. рис. 11.51, В).
Грибы с талломами в форме пузыревидных одиночных клеток или несептированных гиф раньше трактовались как фикомицеты («грибы- водоросли») и противопоставлялись грибам с септированным мицелием. Поперечные стенки последних имеют центральную, простую или комплексную, пору. Пора чаще всего сквозная и может обеспечивать миграцию плазмы и ядер. Плазма находится внутри гиф в интенсивном движении.
Название «фикомицеты» («грибы-водоросли») следовало бы оставить за грибами, для которых установлено родство с водорослями (т. е. за Ооmycota; см. рис. 11.9).
В качестве запасных веществ широко встречаются гликоген и жир, реже маннит и другие вещества; крахмал как запасное вещество у грибов не встречается.
Размножение осуществляется разного рода специальными клетками, которые при эндогенном возникновении называются спорами. Конидии образуются всегда экзогенно и служат для бесполого размножения, как исключение — для переноса ♂ ядер при половом размножении. У грибов, обитающих в воде, споры часто голые, со жгутиками (зооспоры планоспоры). у обитающих на суше они одеты клеточной стенкой и лишены жгутиков (апланоспоры). Споры могут при наличии полового процесса возникать после мейоза (мейо- споры) или образовываться после митотических ядерных делений (митоспоры). Некоторые грибы могут также размножаться путем распада мицелия на отдельные клетки (оидии). Часто возникают покоящиеся структуры в форме жестких клубневидных сплетений гиф (склероциев). Примечательна группировка мицелия в шнуровидные тяжи длиной в несколько метров (ризоморфы), которые служат для распространения (например, у Аrmillarа melleа).
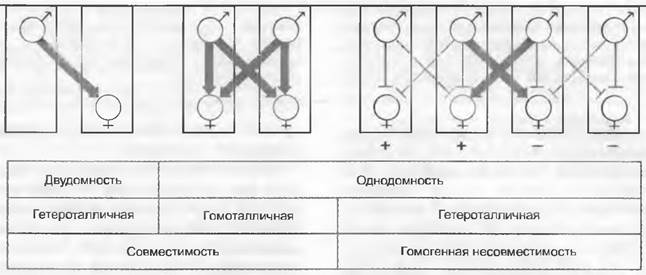
Рис. 11.19. Взаимодействие полов у грибов. Прямоугольники означают мицелии с ♂ (отдающими ядра) и ♀ (воспринимающими ядра) органами. Двудомность: ♂ и ♀ органы на разных мицелиях. Однодомность: ♂ и ♀ органы на одном и том же мицелии. Гетероталлизм: отдельный изолированный мицелий не может образовывать зиготы. У однодомных грибов гетероталлизм может быть обусловлен гомогенной несовместимостью, т. е. только такие ♂ ядра совместимы с ♀ ядрами, которые различаются с ними по фактору скрещивания: - ♂ х +♀ и +♂ х - ♀, но, например, не -♂ х - ♀
При половом размножении копулируют гаметы (изо-, анизо- или оогамия), целые гаметангии (гаметангиогамия), гаметы или же конидии с гаметангиями (гамето- или же конидио-гаметангиогамия) или две клетки таллома, не дифференцированные как специальные половые клетки (соматогамия). Гаметангии (если они имеются) никогда не бывают окружены покровом из многих клеток, поэтому их по аналогии с водорослями называют не антеридиями и оогониями (ср. Вryophytinа), а в зависимости от дифференцировки, способа образования и дальнейшего развития — просто как ♂ или ♀ гаметангии, как спермогонии (♂), сперматангии (♂), оогонии (♀) и аскогоны (♀); имеются и другие обозначения.
Органы, образующие споры, у грибов (как и у водорослей) также никогда не бывают окружены покровами из стерильных клеток. Поэтому в настоящее время их называют не спорангиями, а спороцистами, в отличие от органов с теми же функциями у высших наземных растений (например, мхов, папоротников).
Специальное название для лишенных клеточных покровов структур, которые образуют гаметы или ядра гамет (т. е. название «гаметоцисты» вместо названия «гаметангии»), излишне. Скорее следовало бы сохранить установившееся понятие «гаметангий» и производные от него термины, такие, как «гаметангиогамия» и т.д. Следует также в дальнейшем придерживаться принципа: не вводить без крайней необходимости новых понятий. Гаметангии, защищенные стерильными клетками, напротив, должны называться антеридиями и архегониями.
Часто преобладает бесполое размножение, в некоторых случаях половое размножение неизвестно или же в ходе эволюционного развития было утрачено. Те части таллома, которые образуют репродуктивные структуры без смены ядерных фаз (митоспоры, конидии и т. д), у грибов называются «побочными спороношениями» (анаморфы). В противоположность им основное спороношение (телеоморфа) состоит из частей таллома, в которых происходит слияние ядер (кариогамия) и смена ядерных фаз (мейоз).
Разделение полов и дифференцировку на ♂ и ♀ органы или же репродуктивные клетки часто бывает трудно выявить. Тем не менее отдаваемое ядро можно трактовать как ♂, воспринимающее — как ♀. Исходя из такой предпосылки, можно различать однодомное и двудомное разделение полов (рис. 11.19). Двудомность имеет место тогда, когда один мицелий является либо донором ядер, либо же их реципиентом (рис. 11.19, слева). При однодомности каждый мицелий может быть, как донором, так и реципиентом ядер.
Следующая пара понятий, часто используемая при описании полового размножения грибов, не совсем соответствует рассмотренным выше обозначениям и основана на других генетических особенностях. Гомоталличные грибы в культурах образуют из единственных спор зиготы и соответственно плодовые тела, тогда как у гетероталличных видов для этого необходимы два мицелия различных типов скрещивания (например, «+» и «-»).
У однодомно-гетероталличных грибов слияние ядер одного-единственного мицелия невозможно (как в случае двудомности). Генетически эта несовместимость основывается на наличии по крайней мере двух аллелей одного фактора скрещиваемости, которые обозначают «+» и «-» (или другими символами). Ядра с идентичными знаками (например, «+» и «+») несовместимы и друг с другом не сливаются — в таком случае говорят о гомогенной несовместимости (рис. 11.19, справа). Гетерогенная несовместимость установлена при скрещивании географических рас одного вида; она основывается на несовместимости разных знаков.
Подвижные клетки (зооспоры, гаметы) грибов могут иметь разные типы жгутикового аппарата: опистоконтный — с единственным гладким задним жгутиком; акро-
контный — с единственным перистым передним жгутиком; гетероконтный — с двумя жгутиками, из которых один гладкий, а другой перистый.
Гаметангии отделяются поперечной перегородкой от несущих их гиф. Шаровидные оогонии содержат сначала много ядер, большинство из которых, однако, дегенерирует, после чего вокруг каждого из оставшихся ядер группируется плазма (оосфера), которая сокращается, превращаясь в шаровидную голую яйцеклетку. Эти яйцеклетки, от одной до нескольких, свободно располагаются в оогонии, т. е. не окружены периплазмой. Многоядерные ♂ гаметангии закладываются как целая структура, не образующая половых клеток. Она хемотропически (за счет антеридиола, оогониола) привлекается к оогонию и дает простые или разветвленные оплодотворяющие выросты внутрь оогония, которые достигают яйцеклеток (гаметангиогамия: рис. 11.21, Е, F) аналогично оплодотворению пыльцевой трубкой у семенных растений. В каждую яйцеклетку поступает только одно ядро, которое и сливается с ядром яйцеклетки. После этого яйцеклетка превращается в
зиготу с жесткой оболочкой, устойчивой к микробной инфекции.
Зиготы после периода покоя прорастают без редукционного деления многоядерной проростковой трубкой, которая чаще всего вскоре образует проростковую спороцисту (см. рис. 11.21,1). Имеются однодомные (♂ гаметангии и оогонии на одном и том же талломе) и двудомные виды.
Рис. 11.20. Oomycota: А — Oomycetes, перистый жгутик (слева) и гладкий жгутик (справа) зооспоры Phytophthora infestans (8 000х); В — Achlya (Oomycetes, гетероконтная зооспора с перистым передним и гладким задним жгутиком); С — Rhizidiomyces (Hyphochytridiomycetes, зооспора с единственным передним перистым жгутиком); D— Saprolegniales, жизненный цикл. Светлые линии — гаплофаза, темные — диплофаза; R! — редукционное деление. Диплогенотипическое определение пола (о = ♂ ядро; • = ♂ ядро)
2. Порядок: Leptomitales. Это живущие в воде сапротрофы с регулярно перешнурованными, однако не септированными гифами и пузыревидными спороцистами. В оогонии развивается, как и у представителей следующего порядка, ооспо- ра, окруженная периплазмой. Leptomitus — обитатель сильно загрязненных (сточных) вод.
3. Порядок: Peronosporales. Сюда относятся паразиты, которые поражают преимущественно наземные высшие растения, вызывая у них заболевание, известное как ложная мучнистая роса (особенно Регопо- sporaceae). Растущие между клетками в ткани хозяина гифы гриба посылают в живые клетки короткие выросты — гаустории (рис. 11.22, D). Чаще всего мицелий выходит из устьиц хозяина и образует здесь разветвленные спороцистоносцы (рис. 11.22, А), в совокупности видимые невооруженным глазом как плесневые налеты. Спороцисты отличаются от таковых у Saprolegniales тем, что отрываются от несущих их гиф обычно в виде округлых или эллипсоидных образований. Чаше всего (например, у Plasmo- para) целые (!) спороцисты переносятся ветром на листья других растений, где они при наличии водяных капель (дождь, роса) высвобождают свое разделившееся тем временем содержимое в виде нескольких почковидных зооспор (соответствующих вторичным зооспорам Saprolegnia, рис. 11.22, С; здесь, таким образом, дипланетизма нет).
Рис. 11.21. Oomycota, Saprolegniales: А — спорангий, который дает зооспоры s с двумя жгутиками на переднем конце (200х); В — второй тип спор с боковым расположением жгутиков (примерно 350х); С — участок несептированной гифы с многочисленными ядрами (500х); D — гифа с половыми органами: а — ♂ гаметангий, который дал оплодотворяющие отростки внутрь оогония, о— оплодотворенные яйцеклетки (600х); Е — оплодотворяющий отросток с ♂ ядрами; F — ♂ ядро внедряется в яйцеклетку; G — зигота со слившимися ядрами (Е — G — 600х); Н — проростковая трубка; J — проростковый спорангий с еще неподвижными зооспорами (Н, J — 1 400х). А, В, D — Saprolegnia mixta; С — Thraustotheca; Е — G — Achlya flagellata; Н — Isoachlya intermedia; J — Thraustothreca primoachlya
В связи с прогрессирующей адаптацией к наземному образу жизни(cp. Trentepohliophyceae среди водорослей; см. рис. 11.93) зооспороцисты Peronosporales все больше превращаются в конидии.
У Pythium зооспороцисты, фиксированные на несущих их гифах, всегда высвобождают зооспоры. Спороцисты очень сходны с вегетативными гифами. У Phytophthora, Plasmopara и Pseudopero- nospora спороцисты отрываются и переносятся ветром; они прорастают, как правило, зооспорами, однако при особых внешних условиях (малая влажность!) образуют также проростковые трубки. У Phytophthora спороцистоносцы уже четко отличаются от остальных гиф; после отделения спороцист они могут расти дальше как вегетативные гифы. У Рlasmoрага, Рseudoperonospora и Реronospora спороцистоносцы (конидиеносцы) морфологически дифференцированы специфическим для каждого рода образом; нарастания их после споруляции не происходит. Спороцисты Рeronospora прорастают только инфекционными гифами; гомологичные спороцистам органы стали, таким образом, полностью конидиями, которые за счет изменения формы несущей их структуры при уменьшении влажности могут активно отбрасываться.
Половое размножение. Половые органы возникают внутри растения-хозяина: оогонии — как округлые вздутия концов гиф, ♂ гаметангии — выросты в виде трубок (рис. 11.22, Е). И те, и другие органы отграничиваются поперечной перегородкой и содержат много ядер. Резкое разграничение ♂ гамет отсутствует; яйцеклетка каждого оогония (как правило, единственная) окружена периплазмой. Оплодотворение и формирование зиготы в оогонии представлены на рис. 11.22, Е — G, на примере родов Pemnospora и Albugo.
Рис. 11.22. Oomycota, Peronosporales. Plasmopara, Р. vitícola (A — D); А — спорангиеносцы, выходящие из устьиц; В — оогонии (с cf гаметангием) и зиготами (100х); С — образованиеи выход зооспор (600х); D — прорастание зооспор (z) через устьица в межклетники (250х); Е — Peronospora, Р. parasitica, молодой многоядерный оогоний (о) и ♂ гаметангий (g); F, G — Albugo, A. candida: F — оогоний с оплодотворяющим отростком (b) cf гаметангия, который вводит ♂ ядро, G — зигота в оогонии, окруженная молодой стенкой и периплазмой (р) (Е— G — 600х); Н — Pythium, Р. ultimum, зигота, прорастающая зооспорами (800х) oz — центральная одноядерная часть оогония; 1,2,3 — стадии развития
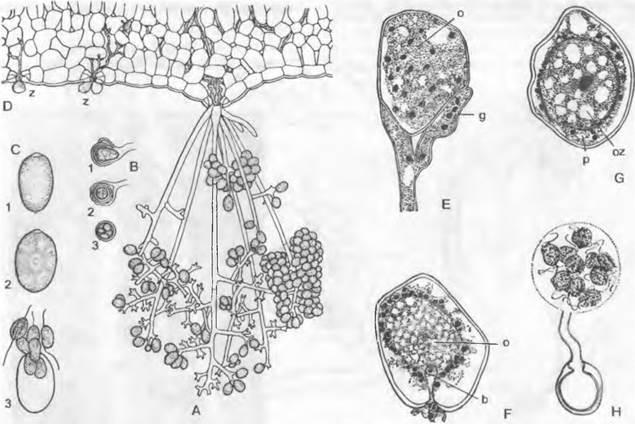
Число способных к оплодотворению ядер в ♂ гаметангии и в оогонии варьирует от многих до одного в зависимости от рода. В оплодотворенных оогониях возникает по одной ооспоре, которая у некоторых видов может содержать много диплоидных ядер (ценозигота). У некоторых Peronosporaceae (например, Basidiophora entospora) способные к функционированию ♂ гаметангии полностью отсутствуют; происходит попарное слияние ядер в одном оогонии (автогамия).
Некоторые виды размножаются на значительной части своего ареала только бесполым путем. Гетероталличный возбудитель фитофтороза картофеля (Phytophthora infestans) у себя на родине в Южной и Центральной Америке представлен обоими типами скрещивания, так что там установлено и половое размножение. Однако в Европу, Северную Америку и др., очевидно, проник только один из двух типов скрещивания; здесь размножение осуществляется исключительно бесполым путем1.
1 В настоящее время в Евразии представлены оба типа скрещивания, так что стал возможен половой процесс. — Примеч. пер.
Прорастание зигот осуществляется либо непосредственно путем образования зооспор, либо, чаще, путем образования проростковой трубки, которая на конце образует спороцисту с зооспорами (рис. 11.22, Н). У более эволюционно продвинутых форм проростковая трубка, не отчленяя спороцисты, непосредственно внедряется в ткань хозяина (аналогично первращению зооспороцист в конидии).
В пределах родов (особенно Peronospora) виды часто приурочены к одному или немногим хозяевам. Видообразование связано с различным выбором хозяев и первоначально более или менее непрерывным, а по мере дальнейшей эволюции все более дискретным изменением морфологических признаков (например, размера конидий). В то же время эти признаки обусловлены вариабельностью внешней среды (например, влияние на размер конидий помимо возраста также температуры, влажности, субстрата), поэтому генетически обусловленный процесс видообразования и связанное с ним разделение видов могут маскироваться модификациями.
Образ жизни и вредоносность Peronosporales. Лишь немногие представители порядка обитают в пресной воде или почве (некоторые представители Pythiaceae). Будучи в большинстве паразитами наземных растений (например, Peronosporaceae, Pythiaceae), они могут вызывать многочисленные заболевания культурных растений. Эти организмы могут быть распространены по всему земному шару, однако для их развития требуется высокая влажность.
Опасный паразит картофеля — Phytophthora infestans (Pythiaceae); гриб сначала вызывает гниль ботвы и переходит также на клубни, так как во время дождя спороцисты смываются в почву и заражают клубни через чечевички. Зооспоры хемотактически привлекаются корнями; у видов грибов, поражающих определенные виды растений-хозяев, это вызывается только соответствующим видом растения-хозяина. В дождливые годы может быть уничтожено более 20 % урожая картофеля. В XIX столетии эпифитотии фитофтороза оставляли без пищи население целых больших регионов. Так, в 1845 — 1846гт. фитофтороз картофеля привел к большому голоду в Ирландии, сильно уменьшившему ее население, после чего последовала волна эмиграции в США. В Ирландии и сейчас не достигнута прежняя численность населения — 8 млн человек.
В хозяйственной деятельности человека значение имеет также вызываемая Plasmopara vitícola (Peronosporaceae) ложная мучнистая роса (милдью) виноградной лозы (рис. 11.22, А), при сырой погоде поражение приобретает характер эпифитотии. Ягоды превращаются в кожистые гнилые образования. Из урожая винограда ежегодно уничтожается примерно 20 % этой и другими, менее важными, грибными болезнями (остальные 20 % — вредителями). Поражения ложной мучнистой росой встречаются также на сахарной свекле, луке, хмеле и других культурных растениях. В 1959 г. в Европе впервые появился (ранее встречалась в Америке и Австралии) возбудитель пероноспороза табака — Peronospora tabacina (эту болезнь еще называют голубой плесенью из-за беловато-голубоватой окраски массы конидий). В дождливое лето 1960 г. гриб уничтожил значительную часть плантаций табака в Средней Европе. Pythium debaryanum — гриб, широко распространенный в почве, вызывает полегание («черную ножку») у сеянцев различных растений, отчего они гибнут. Для борьбы с грибами ложной мучнистой росы (настоящая мучнистая роса — см. Erysiphales) используют опрыскивание листьев содержащими медь фунгицидами (исходно медно-известковую смесь), что подавляет прорастание спороцист.
Обзор оомицетов. Объединяемые в этом классе порядки показывают переход от водного к наземному образу жизни, постепенную замену зооспор конидиями и переход от гидрохорного к анемохорному распространению. С этими эволюционными явлениями, которые достигают вершины у Рeronosporaceae связаны повышение биологических потребностей и все большее проявление паразитических свойств. Это выразилось в переходе от сапротрофизма к паразитизму, специализации относительно определенных хозяев и их органов, а также, наконец, в том. что хозяин начинает поражаться лишь частично. В азотном питании наблюдается прогрессирующее (не связанное с вышеназванными тенденциями) ограничение органическими соединениями азота. Если некоторые Рeronosporales могут использовать наряду с аммонийным также и нитратный азот, то Saprolegniales и Leptomitales не способны к этому, а последние, кроме того, не усваивают и аммонийный азот и используют только его органические соединения. В размножении участвуют лишь определенные части таллома, а остальные продолжают рост («эукарпия»); только у самых примитивных форм (например, Lagenisma из Lagenidiales; Thraustochytridiales1) в кaчестве гаметангия функционирует таллом в целом («холокарпия»).
1 Траустохитриевые в настоящее время относят не к оомицетам, а к отделу либиринтуло- вые. — Примеч. пер.
Филогения. Для оомицетов, родственных автотрофным водорослям из типа Неterokontophyta (см. приложение 4 к Rhodobionta; рис. 11.9). общими признаками являются гетероконтные жгутиковые стадии, сифональное строение таллома и целлюлоза в клеточных стенках. Этим они отличаются от всех прочих грибов. Правда, тип размножения оомицетов — гаметангиогамия и связанная с ней утрата имеющих жгутики гамет сильно видоизменены. В качестве подвижных стадий выступают только служащие для бесполого размножения зооспоры. Последние, за исключением весьма продвинутых в эволюционном отношении форм (например, Регоnospoга), составляют значительную долю отдела.
Небольшой класс гифохитрцдиомицеты (Нуphochytridiomycetes) (15 видов) отличается признаками, частично сходными с оомицетами. Хотя их подвижные стадии обладают только одним, направленным вперед по ходу движения жгутиком, он, как и у оомицетов. перистый (рис. 11.20, С). В клеточных стенках наряду с хитином содержится целлюлоза. Однако этих признаков недостаточно для однозначного определения родственных связей гифохитридиомицетов. Они обитают в пресной и морской воде как паразиты водорослей и грибов или сапротрофы на остатках растений и насекомых. Виды этого класса, например, с простыми бутылевидными клетками внутри клеток хозяина (бурой водоросли Еctocarpus), чаще всего «холокарпные».
Третий отдел: Неterokontophyta
Относимые сюда группы из Нeterokontobionta представлены фотоавтотрофами. Фотоавтотрофность возникла на ранних этапах эволюции у исходно гетеротрофных клеток в результате захвата ими в качестве эндосимбионтов фотосинтезирующих организмов, в этом случае уже эукариотных. Возникшая таким образом новая фототрофная форма организации имеет пока примитивное и чаще всего мало расчлененное строение водорослевого типа организации (см. бокс 11.5). Этот отдел рассмотрен ниже, вместе с прочими фотоавтотрофными эукариотами (см. приложение 4 к Rhodobionta).