БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
Четвертое подцарство: хитиновые грибы (Mycobionta)
Гетеротрофные организмы, эволюционно близкие определенным группам животных (исходным Metazoa из типов губок или стрекающих), в клеточных стенках которых вместо целлюлозы содержится чаще всего хитин, образуют большую группу грибов в более узком смысле. Они не относятся к растениям и даже не родственны им. однако традиционно остаются объектом ботаники.
Единственный отдел: Eumycota
В связи с прогрессирующей адаптацией к жизни вне воды более подвинутые классы «настоящих грибов» (Eumycota) полностью утрачивают снабженные жгутиками зооспоры и гаметы. Они встречаются у наиболее примитивных представителей, но их органелла движения представляет собой единственный гладкий жгутик, на
правленный назад по ходу движения (опистоконтные подвижные стадии). Что касается строения таллома, то в этом, рассматриваемом как монофилетичный, отделе (см. рис. 11.9) достигнуты все известные для грибов уровни организации. В богатых видами более высокоорганизованных классах на определенных фазах развития широко встречаются специальные плектенхимные структуры (плодовые тела).
Клеточная стенка содержит в качестве главного компонента почти всегда хитин (часто совместно с глюканами), целлюлоза практически полностью отсутствует. Представители некоторых групп содержат в стенке маннан-β-глюканы (Saccharomyces) или галактозамингалактаны (Trichomycetes). Некоторые немногие формы, приспособившиеся к паразитическому образу жизни, полностью утратили клеточную стенку, так что в цикле развития встречаются вторично голые протопласты (например, Olpidium, см. рис. 11.23, С).
Оплодотворение может происходить в виде изогамии, анизогамии, оогамии (редко), гаметангиогамии и соматогамии. При гаметангиогамии почти никогда не участвуют оогонии с яйцеклетками. Покоящиеся структуры никогда не возникают внутри оогониев (cp. Oomycota). Большинство представителей — гаплонты, гаплодиплонты или гаплодикарионты; диплонты составляют исключение. В цикле развития все большую и большую часть занимает дикариофаза.
Многие виды образуют каротины (известные у водорослей как дополнительные пигменты); они выполняют функцию фоторецепторов при фототропных изгибах (Pilobolus). Наряду с ними встречаются многие другие пигменты, которые относятся к самым различным структурным типам. Часто присутствуют фенольные пигменты (см. рис. 11.60), также известны азотсодержащие гетероциклы, однако антоцианы и флавоны, широко распространенные у последующих отделов, практически полностью отсутствуют.
1. Класс: хитридиомицеты (Chytridiomycetes)
Хитридиомицеты существуют в виде отдельных клеток или образуют многоядерный таллом без поперечных перегородок (сифональный). Подвижные клетки (гаме
ты и зооспоры) опистоконтные (см. рис. 11.25, F). Вместо ядрышка чаще всего встречается богатый РНК «ядерный колпачок» (см. рис. 11.25, Е).
Особенности размножения можно рассмотреть на примере Allomyces (Blastocladiales). Этот «эукарпный» почвенный гриб уже имеет обильно разветвленные, многоядерные гифы. Жизненный цикл проходит как изоморфная смена генераций (у группы «Eu-Allomyces», см. рис. 11.25, Q).
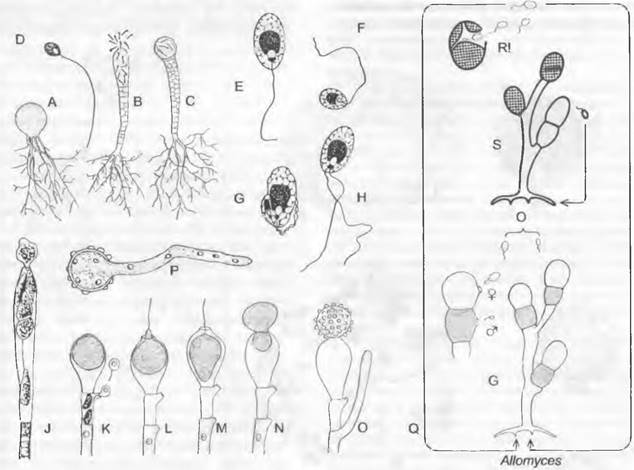
Гаметофит образует на концах гиф отделенные септами гаметангии; ♀ гаметангий чаще всего сидит непосредственно на ♂ гаметангии (однодомность). Оба через специальные поры высвобождают гаметы; ♂ гаметы мельче и окрашены у-каротином в оранжево-красный цвет; ♀ гаметы бесцветные; ♀ гаметы выделяют сиренин (см. 8.2.1.1) и путем хемотаксиса привлекают ♂ гаметы. После анизогамии возникает диплоидная зигота, только в самом начале имеющая жгутики. Из нее прорастает спорофит, который по размеру и общему виду сходен с гаметофитом. На отдельных участках гиф спорофита развиваются спороцисты двух разных типов. Расположенные по бокам гиф, чаще всего попарно друг над другом, тонкостенные митоспороцисты, открывающиеся сосочком, высвобождают после исключительно митотических ядерных делений диплоидные митозооспоры; последние прорастают снова в диплоидный спорофит, так что смена генераций нарушается за счет интенсивного бесполого размножения спорофита (побочное спороношение с митоспороцистами). На концах гиф, часто поодиночке, обособляются толстостенные, имеющие орнаментацию в виде пчелиных сот, темноокрашенные мейоспороцисты, которые отпадают целиком. После периода покоя (как гипноспороцисты) они претерпевают мейоз и высвобождают гаплоидные мейозооспоры, прорастающие в гаметофиты.
Группа «Eu-Allomyces» представлена видами A. arbúscula (♂ гаметангии расположены на ♀) и A. macrogynus (♀ на ♂). Гибрид этих двух видов, встречающихся в разных вариантах плоидности (A. arbúscula: n = 8,16, 24, 32; A. macrogynus: n - 14, 28, 56), — А.х javanicus — дает мейоспоры, имеющие меньший процент прорастания (0,1 — 3,2 %) и образующие гаметофиты с различным расположением гаметангиев. От полной смены генераций имеются отклонения с редукцией гаметофита (например, Вгасhу-Аllоmусеs).
Три порядка хитридиомицетов отличаются строением таллома, характером бесполого размножения и ультраструктурой зооспор. Известно около 500 видов. Большинство видов живет в воде, некоторые также в почве или как паразиты в клетках высших растений1.
1 К сожалению, здесь совершенно не рассматривается крайне интересный порядок Neocallimasticales, ныне также относимый к хитридиомицетам. Виды этого порядка — облигатные анаэробы (этим данный порядок уникален среди всех грибов в целом), обитающие в рубце и слепой кишке жвачных животных. Некоторые виды имеют типичные для хитридиомицетов одножгутиковые зооспоры, у некоторых жгутиков более 10. Митохондрий у этих грибов нет (они считаются утраченными в ходе эволюции) — это естественная ситуация для анаэробных организмов. Для некоторых видов показана способность образовывать покоящиеся споры, которые могут сохраняться в высушенном помете в аэробных условиях. Неокаллимастиксовые — обязательный компонент рубца жвачных. Считается, что они играют важную роль в переваривании пищи. — Примеч. пер.
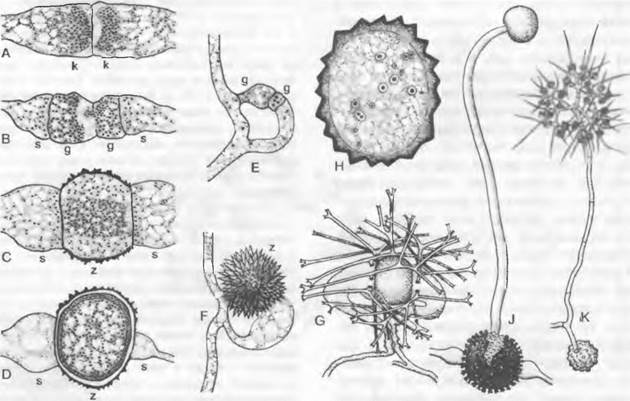
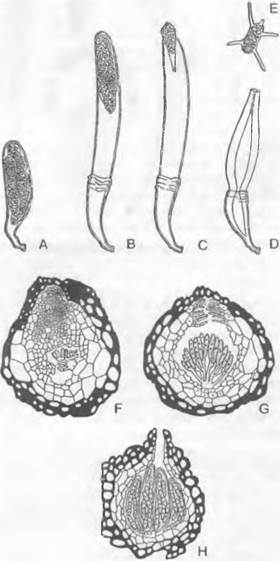
1. Порядок: Сhyridiales. Таллом грибов, относящихся к этому порядку, развит слабо. Чаще всего он одноклеточный, в виде шара или пузыря: мицелий из гиф не образуется, однако часто имеются тонкие придатки одной-единственной клетки, лишенные собственных ядер (ризомицелий) Половое размножение происходит как изогамия, анизогамия или гаметангиогамия. Определение пола генотипическое (Olpidiасеае: Rozellа) или модификационное (Synchytrium). Обычно в образовании спор или гамет участвует весь таллом («холокарпия»); более подвинутые формы на ризомипелии развивают специальные собирательные клетки, которые могут давать начало репродуктивным клеткам («эукарпия»). Зооспоры содержат одну хорошо заметную каплю масла.
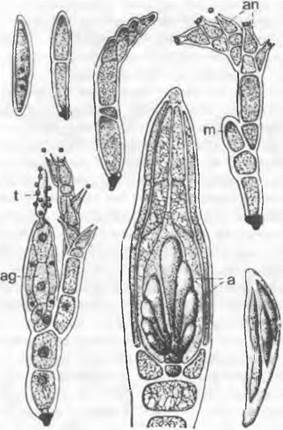
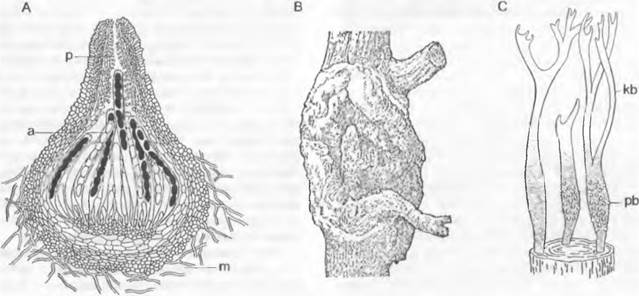
Цикл развития низших Chytridiales можно рассмотреть на примере Olpidiасеае. Голый протопласт ведет паразитический образ жизни в клетке растения-хозяина, увеличивается в раз мере и окружается хитиновой стенкой (рис. 11.23, С, D), претерпевает ядерные деления и деление цитоплазмы и образует в результате опистоконтные подвижные стадии; на их образование идет весь протопласт. Эти подвижные стадии либо инфицируют новые клетки хозяина, как зооспоры, или копулируют, как гаметы, попарно друг с другом (факультативное определение функции; изогамия) с образованием голых двужгутиковых планозигот (рис. 11.23, Б). Последние внедряются в клетки хозяина и позже превращаются там в гипнозиготы с жесткой стенкой; два половых ядра сливаются только следующей весной (прообраз дикариофазы; рис. 11.23, G, Н), а затем образуют (очевидно, после редукционного деления) многочисленные подвижные стадии, которые высвобождаются через специальный сосочек.
Рис. 11.23. Еumycota, Chytridiomycetes, Сhytridiales, Olpidium viciae: А — зооспоры; В — внедрение в клетку хозяина; С — голый протопласт гриба в клетке хозяина; D — зооспорангий, могущий служить как гаметангий; Е — зооспорангий опорожненный; Б — копуляция двух опистоконтных гамет (А — F — 500х); G — молодая, еще двухъядерная зигота (600х); Н — инцистированная зигота; I — инцистированная зигота, прорастающая (Н, J — 120х)
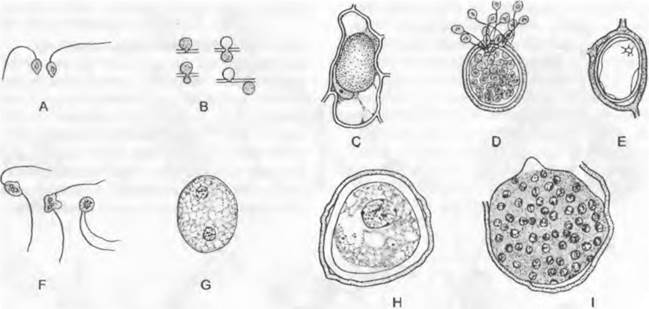
Olpidium brassicae— возбудитель «черной ножки» проростков капусты. Родственные Synchytriaceae живут как эндопаразиты в цветковых растениях, где могут вызывать разрастания типа галл; Synchytrium endobioticum вызывает рак картофеля. Все эти грибы развиваются внутри тканей хозяина — в полностью пораженных клетках; кроме того, они «холокарпные».
У последующих семейств не весь таллом идет на образование репродуктивных клеток («эукарпия»).
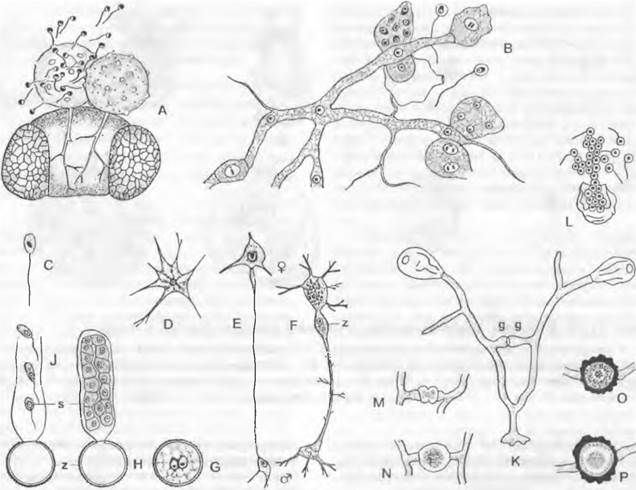
Rhizidiасеае — часто встречающиеся паразиты планктонных водорослей и пыльцы (Rhizophydium; рис. 11.24, А). Таллом «функционально» разделяется на репродуктивный пузырек, расположенный вне субстрата, и питающий ризоид, который внедряется в клетку хозяина. Центр моноцентрического таллома у Polyphagus euglenae образует один-единственный «центральный пузырь», от которого отходит несколько ризоидов. Этот гриб питается за счет того, что он своими ризоидальными выростами внедряется в водоросли из рода Еuglenaи высасывает их содержимое. Один экземпляр Роlyphagus euglenae может поразить свыше 50 эвглен (рис. 11.62, А). Половое размножение — анизогаметангиогамия: более мелкие ♂ особи посылают «поисковый ризоид»; встретившись с центральным пузырем ♀ особи, он сливается с ней, и в место слияния переходят соответственно ♂ и ♀ и ядра копулирующих грибов (см. рис. 11.42, Е, F). Возникающая зигота имеет толстую шиповатую стенку и претерпевает состояние покоя как дикариотическая гипнозигота («покоящаяся спора», G). Слияние ядер (кариогамия) и, вероятно, мейоз происходят при прорастании гипнозиготы (Н); в проростковой трубке в результате ядерных делений и одновременного распадения цитоплазмы образуются многочисленные зооспоры (вероятно, мейозооспоры). После освобождения они садятся на эвглены, инцистируются, а затем циста прорастает и возникает «центральный пузырь» с ризоидами, которые дальше поражают клетки эвглен.
В обоих последующих семействах для выхода зооспор у спороцисты образуется крышечка (оперкулятный тип — в противоположность ино- перкулятному у предыдущих семейств).
У Chytridiaceae формируется моноцентрический, у Мegachytriасеае чаще всего полицентрический таллом с несколькими пузыревидными зооспороцистами, связанными ризоидальными тяжами. Половое размножение в немногих исследованных случаях представляет собой гаметангиогамию. У Zygochytrium (Меgachytriасеае) две копуляционные ветви, в данном случае равнозначные, растут навстречу друг другу, и в месте слияния их концы (рис. 11.24, К) образуют толстостенную гипнозиготу; о поведении ядер до образования зиготы ничего не известно.
Рис. 11.24. Chytridiomycetes Chytridiales: А — Rhizophydium halophilum, зооспорангий с сосочками, через которые выходят опистоконтные зооспоры, на пылинке Pin us с гаусториями внутри; В — Polychitrium aggregatum, небольшой многоядерный неклеточный мицелий с развитыми по-разному спорангиями и двумя опистоконтными зооспорами (А, В — 400х); С — J — Polyphagus euglenae (примерно 450х): С — зооспора, D — таллом, образующий ризоиды, Е — копуляция между мелкой ♂ и более крупной ♀ особями, F — ♂ ядро в будущей зиготе, G — зигота с еще не слившимися ♂ и ♀ ядрами, H, J — развитие и освобождение от зооспор (s) зооспороцисты (z); К — Р — Zygochytrium aurantiacum (350х): К — таллом с двумя расположенными на концах гиф, уже опустевшими зооспорангиями и двумя копулирующими гаметангиями (g), L — зооспорангий в процессе освобождения от зооспор, М — Р — образование зиготы (z) из копулировавших гаметангиев, Р — зрелая гипнозигота (z) (зигоспора)
Имеются случаи, когда копулирующие с образованием или без образования трубки гаметангии многоядерны. Такие процессы очень напоминают картину, с которой мы позже столкнемся у зигомицетов. Если у Сhytridiales гетерофазная смена генераций еще не встречается (только у Рhysoderma1 есть указания на нее), в следующем порядке она достаточно выражена.
1 В настоящее время род Physoderma обычно относят к следующему порядку — Вlastocladiales — Примеч. пер.
2. Порядок: Вlastocladiales. Представители порядка образуют чаще всего таллом, состоящий из гиф, которые обособляют несколько вмести
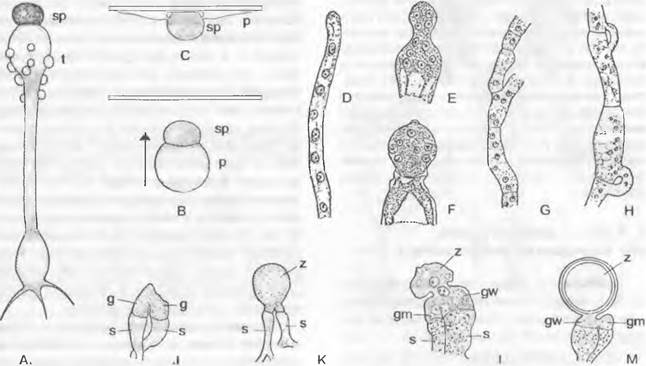
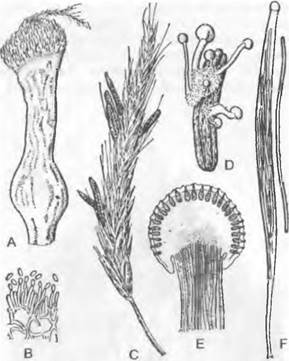
лищ репродуктивных клеток путем образования поперечных перегородок на своих концах. К субстрату таллом прикрепляется ризоидоподобными выростами. Наиболее примитивные представители внешне близки к Сhytridiales, например, обитающая в почве «холокарпная» Вlastocladiella (рис. 11.25, А — D). Зооспоры содержат несколько не особенно крупных капелек масла и прорастают двумя проростковыми трубками. Большинство представителей обитает сапротрофно в почве. воде, часто на остатках растений и животных. Два вида Вlastocladiella поражают синезеленые водоросли. Развитие Аllomyces было рассмотрено выше (введение к хитридиомицетам). Вlastocladiella (рис. 11.25, А) образует талломы с вместилишами репродуктивных клеток только одного типа: каждый гаметофит развивает один гаметангии; сливающиеся изогаметы различаются только окраской (каротин). Спорофит несет либо одну митоспорописту, либо одну мейоспороцисту.
3. Порядок: Моnoblepharidales. Порядок отличается от предыдущих, наличием четко отграниченной базальной части таллома, несущей ризоиды, а также прежде всего оогамным размножением. ♀ гаметы представляют собой лишенные жгутиков яйцеклетки, которые после оплодотворения имеющими жгутик ♂ гаметами (оогамия) развиваются дальше в ооспоры. Несмотря на более прогрессивную, чем у Blastocladiales, форму размножения, они остались гаплонтами: мейоз у них происходит при прорастании зиготы (зиготическая смена ядерных фаз); смена генераций отсутствует.
Для полового размножения служат вздутые одноядерные оогонии (рис. 11.25, К), расположенные чаще всего на концах гиф; их содержимое редуцировано до одной-единственной одноядерной яйцеклетки1. Расположенные под оогонием сперматогонии высвобождают некоторое количество одноядерных и одножгутиковых сперматозоидов (К). Последние проникают через специальное отверстие в оогоний и оплодотворяют яйцеклетку (L, М), которая к этому времени либо остается внутри оогония, либо (у большинства видов) выталкивается через специальное отверстие из оогония (N) и здесь становится толстостенной шиповатой гипнозиготой (О); или зигота даже уплывает оттуда с помощью сохраняющегося ♂ жгутика. Зиготы прорастают не зооспорами, а проростковой трубкой (Р), что считается более прогрессивным признаком. Бесполое размножение осуществляется зооспорами (J). Представители порядка обитают как сапротрофы на растительных остатках в воде.
1 У некоторых представителей Моmoblepharidales в яйцеклетках может быть несколько - Примеч. пер.
При обзоре хитридиомицетов можно отметить некоторые важные прогрессивные тенденции. Вместо исходной «холокарпии» все больше развивается «эукарпия». Наблюдается переход от изогамии с факультативным определением функции репродуктивных клеток через изогамию с генотипическим определением пола гамет к анизогамии, оогамии (Мonoblepharidales) и гаметангиогамии. В отдельных случаях кариогамия происходит не сразу после копуляции гамет и связанной с ней плазмогамией; следствием является включение в цикл развития дикариофазы (Olpidium, Роlyphagus). Наряду с однодомностью встречается и двудомность; в отдельных группах наблюдаются смена генераций и ее дальнейшее развитие в диплонтный жизненный цикл (Allomyces). Голые талломы приспособились к существованию в качестве эндопаразитов. Истинные наземные формы с воздушным мицелием в этой группе еще не возникли.
Рис. 11.25. Chytridiomycetes: А — Н — Blastocladiales, А — D — Blastocladiella variabilis: А — спорофит, В — зооспорангий в процессе выхода зооспор, С — с покоящимся спорангием (А — С ЗЗх), D — зооспора (450х); Е — Н — Allomyces javanicus (1 000х), Е — ♀, F — ♂ опистоконтная гамета; G — копуляция гамет; Н — планозигота; J — Р — Monoblepharidales, Monoblepharis (300х); J — М. macrandra, спорангий с выходящими из него зооспорами, К — Р — М. sphaerica: К — конец гифы с оогонием и расположенным под ним спермогонием. из которого выходит сперматозоид. L — сперматозисщ внедряется через апикальное отверстие оогония к яйцеклетке и сливается с ней, М — слияние совершилось, N — оплодотворенное яйцо выползает из оогония, О — гипнозигота с толстой шиповатой стенкой, Р — прорастание зиготы; Q — Blastocladiales, Allomyces: схема смены поколений, светлые линии — гаплофаза, темные — диплофаза, G — гаметофит, R! — редукционное деление, S — спорофит
Во всех следующих далее классах (2 — 4) имеющие жгутики подвижные стадии (гаметы, зооспоры) полностью отсутствуют. Широко развивалось приспособление к наземному образу жизни.
2. Класс: зигомицеты (Zygomycetes)
Зигомицеты имеют чаще всего развитый мицелий из гиф, обычно не септированных и многоядерных (ценоцитный мицелий, который соответствует сифональному типу организации водорослей); у некоторых форм имеются поперечные перегородки. При половом размножении никогда не образуется гамет: всегда копулируют
друг с другом два гаметангия, чаще всего многоядерные, нередко имеющие одинаковый облик. Они обособляются от концов, растущих навстречу друг другу гиф (гаметангиогамия)и дают покоящуюся зиготу. Эта так называемая «зигоспора» — результат полового процесса. Она прорастает, претерпевая мейоз, в зародышевую спороцисту, в которой за счет разделения многоядерного плазматического содержимого эндогенно в большом числе возникают мейоспоры.
Бесполое размножение приспособлено к наземному образу жизни, однако несколько иначе, чем у оомицетов. Если у оомицетов спороциста отрывалась, чтобы образовать зооспоры на месте своего прорастания, то у зигомицетов окруженные клеточными стенками споры возникают (за счет деления содержимого) эндогенно, внутри спороцист. Высвобождаясь из спороцисты, они распространяются по воздуху. Аналогичным образом у зигомицетов спороцисты превращаются в конидии, дающие проростковую трубку.
Рис. 11.26. Zygomycetes, Mucorales: А — спорангий на продольном оптическом разрезе Mucor mucedo (225х); В — разрез через зрелый спорангий с многоядерными митоспорами Sporodinia grandis; С — Cunninghamella echinulata: 1 — образование конидий (370х), 2 — конидия (1 000х); D — Thamnidium elegans, разветвления, несущие спорангиоли (200х); Е — жизненный цикл: светлые детали рисунка — гаплофаза, темный круг вверху — диплофаза; с — колонка; s — спорангиоспоры; w — стенка; R! — редукционное деление
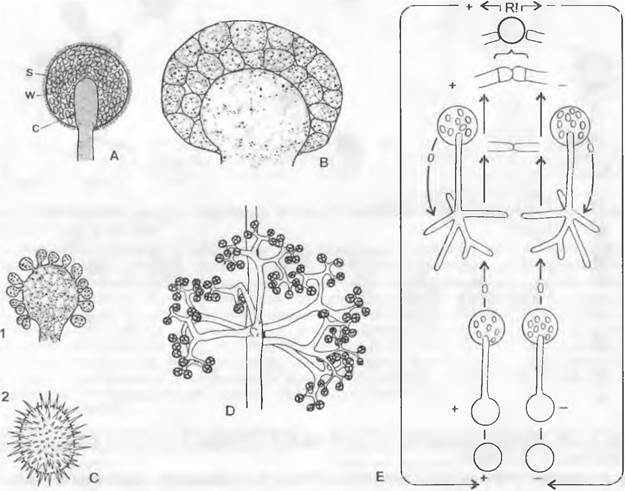
Половое размножение происходит при встрече мицелиев противоположных типов скрещивания (+ и -). Тогда оба мицелия под воздействием выделяемых гамонов образуют булавовидные гаметангии, изгибающиеся навстречу друг другу и, наконец, смыкающиеся концами. Они многоядерные и отделяются от несущих гиф (суспензоров) поперечной перегородкой. Разделительная двойная стенка между гаметангиями исчезает (см рис. 11.27, В), и оба гаметангия участвуют в формировании возникающей зиготы. Последняя представляет собой покоящуюся гипнозиготу с толстой, многослойной, снаружи бородавчатой стенкой («зигоспора» D), в которой попарно сближаются многочисленные половые ядра (+, -). В конце периода покоя немногие ядра, иногда всего одна-единственная пара, претерпевают слияние (кариогамия), тогда как другие пары ядер погибают. Таким образом, в оплодотворении участвуют исключительно гаметные ядра, свободных гамет не бывает. «Зигоспора» прорастает, претерпевая мейоз, проростковой трубкой, причем дальнейшее митотическое деление происходит лишь в одном-единстве ином из получившихся гаплоидных ядер (остальные продукты мейоза дегенерируют): все ядра, следовательно, генотипически одинаковы. На конце проростковой трубки образуется проростковая спороциста (J), которая содержит многочисленные мейоспоры, принадлежащие какому-либо одному (+ или -) типу скрещивания. Внешне она сходна со спороцистой бесполого размножения, однако возникающие здесь споры представляют собой одноядерные мейоспоры одного полового знака (+ или -). В обоих случаях спороцисты отграничиваются от несущей ее гифы поперечной перегородкой, которая в виде так называемой колонки вдается внутрь спороцисты
(рис. 11.26, А). Многоядерная плазма спороцист распадается либо на многоядерные гаплоидные митоспоры, либо на одноядерные гаплоидные мейоспоры.
К этому классу относится примерно 500 видов, преимущественно сапротрофных. Они распределяются по нескольким порядкам.
1. Порядок: Мuсоrаlеs. Споры образуются в спороцистах, причем последние чаще всего вскрываются и высвобождают споры; реже спороцисты содержат немного спор или всего одну спору и отваливаются целиком; их называют спороцистиоли. «Зигоспоры» возникают как часть сливающихся гаметангиев. К этому порядку относятся наземные плесневые грибы, которые ведут преимущественно сапротрофный образ жизни, реже паразитируют на растениях и животных.
Один из наиболее широко распространенных видов — головчатая плесень (Миcor mucedo). Сильно разветвленный, лишенный поперечных перегородок мицелий этого гриба образует белые плесневые налеты на помете животных, хлебе и т. д. Выходящие из субстрата гифы, служащие для поглощения питательных веществ, поднимают вверх вертикальные выросты мицелия, каждый из которых несет на конце округлую спороцисту (рис. 11.26, А). Внутри нее возникают многочисленные приспособленные к наземной жизни споры, имеющие клеточную стенку и способные переносить высыхание. Они округлые, многоядерные, будучи митоспорами, относятся к побочным спороношениям и долгое время сохраняют всхожесть.
У различных видов существует много отклонений в строении спороцист, гаметангиев и зигоспор, а также в особенностях размножения. В частности, имеются такие типы спороцист, которые, как и зооспорангии некоторых хитридио- мицетов, открываются крышечкой (Saksenaеа), тогда как обычно споры освобождаются за счет разрыва стенки спороцисты. Среди побочных спороношений четко прослеживается тенденция к сильному уменьшению числа спор в спороцисте, а также к тому, чтобы сильно уменьшившаяся в размере спороциста распространялась целиком (при утрате колонки, например, у Thamnidium). В конце этого ряда мы встречаем такие спороцисты, которые соответствуют экзогенно отделяющимся конидиям, в которых способность к образованию спор сильно подавлена (например, Нaplosporangium, Вlakestеа, Сhoanephoraа) или полностью пропадает (Сunninghamella). У Сhoanephora в зависимости от внешних условий (питание, температура) образуются «конидии» или же спороцисты со спороцистоспорами. Некоторые виды имеют только конидии. Нередко встречающийся на конском помете пилоболус (Рilobolus) за счет тургорного давления отбрасывает целиком спороцисту черного цвета, расположенную на конце несущей клетки, способной к положительным фототропным изгибам (см. рис. 11.28, А). Дальность полета составляет до 1,8 м в высоту и до 2,4 м в сторону при начальной скорости примерно 10 м/с. При гаметангиогамии у двудомных (рис. 11.27, А) или однодомных видов (Е) участвующие в этом процессе гаметангии обычно имеют одинаковый облик (изогаметангиогамия; А, G) или же отличаются по размеру и поведению (анизогаметангиогамия; Е). Гипнозиготы нередко защищены гифами oбвертки, которые вырастают из суспензоров. Здесь можно видеть прообраз формирования плодовых тел, столь характерных для аскомицетов и базидиомицетов. Под плодовыми телами мы понимаем видимые невооруженным глазом структуры из гиф основного спороношения. У Absidia (анизогаметангиогамия) только суспензор, соответствующий более крупному гаметангию, образует неразветвленные, длинные, коричневые выросты, закрывающие собой зигоспору. У Phycomyces (изогаметангиогамия) зиготу обрастают идущие от обоих суспензоров почти черные, разветвленные придатки (G) У Mortierella развивается вокруг каждой зиготы, а у Endogone (следующий порядок) — вокруг нескольких зигот защитная структура из плотно переплетенных гиф.
Рис. 11.27. Zygomycetes, Mucorales: А — D — оплодотворяющий орган и образование гипнозиготы у Sporodinia grandis (50х); Е, F — то же, у Zygorrhynchus moelleri (75х); G — Phycomyces blakesleeanus, зигота с обрастающими ее придатками, отходящими от суспензоров (30х); Н — Mucor hiemalis, зигота с гаплоидными ядрами, слияние ядер и диплоидные ядра (550х); J — Mucor mucedo, проростковый спорангий (60х); К — Chaetocladium jonesii, прорастание зиготы конидиеносцем (75х); g — гаметангий; к — копуляционная ветвь; s — суспензор; z — зигота
2. Порядок: Endogonales (с родом Endogone и еще несколькими родами). Виды рода Endogone, имеющие клубневидные плодовые тела размером с лесной орех, развиваются под землей и часто образуют микоризу.
Гаметангии после копуляции формируют структуры в виде моста (рис. 11.28, J), из вершины которых выпячиваются шаровидные зиготы (К) Мицелий сначала многоядерный и несептированный, позже появляются поперечные перегородки.
Рис. 11.28. Zygomycetes: А — С — Mucorales, Pilobolus crystallinus: А — несущая клетка (спорангиеносец) со спороцистой (sp); первая покрыта выделяющимися каплями жидкости (t) (20х), В, С —отброшенная спороциста соответственно незадолго до и после столкновения с препятствием: D — Н — Entomophthorales: D — F — Entomophthora muscae (450х): D — конец гифы из мухи, Е— возникающий из нее, прорывающийся наружу конидиеносец, F— образование конидии, G — молодая гифа Entomophthora sciarea (180х); Н — Ancylistes closterii, оплодотворение между соседними клетками (500х); J—М — Endogonales, Endogone, оплодотворение: J — копуляция, К — готовая зигота Е. pisiformis, L — выпячивание зиготы после перехода ядра из ♂ (gm) в ♀ (gw) гаметангий, М — готовая зигота (z) Е lactiflua (300х); р — слизистое кольцо; s — суспензор
3. Порядок: Entomophthorales. Виды порядка размножаются бесполым путем, почти исключительно конидиями, которые, как и у Mucorales, произошли из спороцист. Копуляция гаметангиев осуществляется между двумя гифами или может быть также боковой между соседними клетками одной и той же гифы (рис. 11.28, Н). Гипнозигота, как и у Endogone,образуется в виде выроста слившихся гаметангиев. В гифах встречаются поперечные перегородки; образующиеся таким образом обособленные участки бывают от многоядерных с самым разным числом ядер до одноядерных (рис. 11.28, G), у Basidiobolus ranarum они в основном только одноядерные. У наиболее известного представителя — гриба Entomophthora muscae, вызывающего эпизоотии — заболевания комнатной мухи, многоядерная конидия (рис. 11.28, Е. F) отбрасывается от несущей ее клетки. Попав на муху, конидия образует проростковую трубку, которая внедряется внутрь тела животного и там разви
вается мицелий, убивающий муху. Из трупа в массе вырастают конидиеносцы. От них активно отбрасываются конидии, которые (например, на оконном стекле) окружают мертвую муху светлым венцом. В высохших мухах возникают внутри гиф толстостенные цисты, которые, возможно, следует рассматривать как партеногенетические зиготы.
Близкие к Еntomophthorales представители порядка Zoopagales (4-й порядок) паразитируют на амебах и нематодах, образуя ветви гиф, превращающиеся в гаустории (см. рис. 11.62, в, бокс 11.4)1.
1 К этому же порядку относят и семейство Piptocephalidасеае, представители которого паразитируют на других зигомицетах (Мucoralех). — Примеч. пер.
Спорным является сближение с зигомицетами видов (около 60), объединяемых в класс трихомицеты (Тrichomycetes). Они обитают как паразиты на поверхности или внутри тела насекомых (особенно водных) и имеют сильно редуцированный таллом, клеточные стенки которого состоят из полигалактозамина и галактана. Возникающие в результате гаметангиогамии зигоспоры и отсутствие гамет говорят об их родстве с зигомицетами, которые, однако, имеют в клеточной стенке хитин. Иногда встречающиеся амебоидные стадии свидетельствуют об обособленности этой группы. Примерами родов могут служить Amoebidium, развивающиеся на водных насекомых, и Harpella — в кишечнике мошек (Simuliidae)1.
1 К большому сожалению, здесь не рассматриваются грибы порядка гломусовых (Glomerales, или — неграмотно — Glomales), относимого к зигомицетам или выделяемого в отдельный класс гломеромицеты (Glomeromycetes). Это грибы, образующие наиболее распространенный тип микориз — везикулярно-арбускулярные микоризы (ВАМ). Из числа видов растений, имеющих микоризы, свыше 80 % имеют именно такие микоризы. Представители этого класса, в частности, образуют микоризу с наиболее важными сельскохозяйственными растениями — такими, как злаки. ВАМ относятся к эндомикоризам, т. е. в отличие от эктомикориз, имеющих хорошо выраженный наружный чехол и сеть гиф в межклетниках коры (сеть Гартига), грибы ВАМ сильно не изменяют внешнюю морфологию корней. Только иногда на поверхности корня встречается сетка гиф, соединяющая внутритканевой мицелий с грибом, находящимся в почве. Гриб проникает внутрь клеток корня растения, образуя там древовидно-ветвящиеся образования — ар- бускулы. Помимо них внутри клеток или за их пределами обычно встречаются округлые вздутия — везикулы. Питание растений осуществляется за счет переваривания арбускулов, а иногда к тому же везикулов и гиф. Известны случаи, когда грибная инфекция сосредоточена в специальных отдельных клубеньках. Для воспроизведения и перенесения неблагоприятных условий у этих грибов образуются покоящиеся споры, представляющие собой азигоспоры со сложной 6-слойной стенкой, содержащей хитин и целлюлозу. Они образуются одиночно или рыхлыми группами вне корней растений или, реже, внутри них. — Примеч. пер.
3. Класс: сумчатые грибы, или аскомицеты (Ascomycètes)
Аскомицеты живут, как и базидиомицеты (см. ниже), преимущественно в наземных местобитаниях. Некоторые виды встрeчаются в пресных водах или в морях. По большей же части это паразиты растений или сапротрофы на отмерших растительных тканях и в соках растений. Вегетативное тело — как правило, обильно разветвленный мицелий из септированных гиф; в их поперечных перегородках имеются простые поры. Формы, приспособившиеся к определенному характеру питания, имеют вегетативное тело в виде почкующихся клеток по типу дрожжей. Клеточные стенки состоят из хитина и глюканов (у дрожжей, Еndomycetidае, доля хитина в стенках незначительна или он отсутствует совсем). При сильном увеличении (электронная микроскопия) видно, что стенки двухслойные: внутренний слой светлый, толстый и бесструктурный, внешний слой темный и тонкий.
Половое размножение приводит к образованию сумки, или аска, — характерной мейоспороцисты, часто имеющей мешковидную форму. В сумке происходит: слияние половых ядер (например, «+» и «-»-ядер; кариогамия), редукционное деление (мейоз), а также эндогенное образование мейоспор (= аскоспор) в виде свободных клеток. Кроме того, сумки часто приспособлены к активному выбрасыванию аскоспор. Имеющие жгутики репродуктивные клетки отсутствуют, как у зигомицетов и у полностью приспособившихся к наземной жизни базидиомицетов. В жизненном цикле аскомицетов появляется двухъядерная фаза (дикарион); она характерна для всех аскомицетов (кроме Еndomycetidae) и базидиомицетов.
Аскомицеты, или сумчатые грибы, насчитывая примерно 30 000 известных видов, составляют порядка 30% всех описанных к настоящему времени грибов. Если сюда причислить еше по большей части происшедшие от аскомицетов несовершенные грибы (Deuteromycetes, приложение 1 к Мусоbiontа), то доля аскомицетов в общем числе грибов повысится до 60%. Систематическое подразделение основывается на различном возникновении сумок в жизненном цикле, на строении и типе вскрытия сумок, а также на форме и характере развития плодовых тел. Системы, предлагаемые разными авторами, различаются в зависимости от оценки ими тех или иных признаков. В дальнейшем мы будем исходить из того, что эволюция грибов шла от хитридиомицетов через зигомицеты к аскомицетам.
В первых трех подклассах плодовые тела не образуются.
1. Подкласс: Тафриномицетиды (Taphrinomy cetidae)
Представители этой группы аскомицетов паразитируют на растениях. Плодовые тела не образуются.
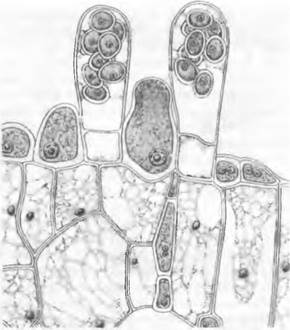
Виды Taphrim могут вызывать у растений различные деформации Некоторые виды провоцируют образование «ведьминых мётел» на косточковых, березах и грабах; Т. deformansобусловливает курчавость листьев персика; Т. pruni превращает завязи сливы в полые внутри, лишенные косточки галлы, так называемые кармашки. Сумки возникают из коротких, сначала двухъядерных, затем одноядерных диплоидных клеток гиф между кутикулой и эпидермисом хозяина, после редукционного деления и нескольких митозов. Они прорываются между клетками эпидермиса растения- хозяина, образуют палисадный слой (рис. 11.29) и открываются на вершине простой трещиной, через которую освобождаются аскоспоры. При прорастании аскоспоры почкуются, напоминая этим клетки дрожжей. Затем на поверхности растения-хозяина возникает сначала гаплоидный, сапротрофно живущий почкующийся мицелий. При слиянии вегетативных клеток или автогамной попарной группировки ядер формируется стадия дикариотичного мицелия, который теперь внедряется как паразит в межклетники хозяина. Его дальнейшее развитие осуществляется независимо от гаплоидного мицелия. В этом плане Taplirinomycetidae сходны скорее с базидиомицетами, имеющими самостоятельную дикариофазу, чем с аскомицетами (рассматриваются ниже).
Рис. 11.29. Ascomycetes, Taphrinomycetidae, Taphrina deformans: кариогамия и зрелые сумки (800х)
K Taphrinomycetidae близки следующие порядки.
Protomycetales. Сюда относятся некоторые паразиты цветковых растений, которые вызывают характерные изменения окраски или пузырчатые вздутия на своих хозяевах.
Ascosphaerales. Ascosphaera apis вызывает болезнь пчел.
2. Подкласс: Эндомицетиды (Endomycetidae)
В этом подклассе объединены дрожжевидные аскомицеты. Дрожжи — это грибы, которые размножаются почкованием, как пекарские дрожжи (см. рис. 2.34); другой отличительной особенностью их может быть размножение фрагментами гиф (артроспорами). Сумки возникают непосредственно из зигот или других отдельных клеток, не после Дикариофазы и не в плодовых телах. Стенка сумки распадается или ослизняется при созревании спор, таким образом аскоспоры не отбрасываются. Таллом у некоторых представителей распадается на отдельные клетки. Обычно он представляет собой почкующийся мицелий, реже — септированный нитевидный мицелий. Грибы, относящиеся к этому подклассу, часто живут в богатых сахарами субстратах (например, в истечениях сока деревьев и кустарников, в нектаре).
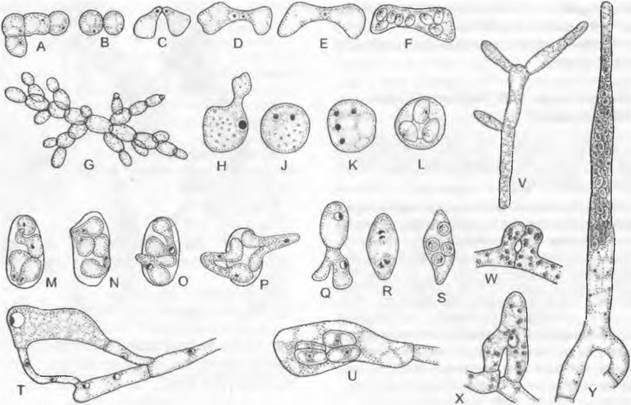
К роду Saccharomyces и близко родственным родам (Hansenula, Schizosaccharomyces1) относятся широко известные, часто применяемые на практике и в научных исследованиях дрожжи. Их округлые или овальные одноядерные клетки размножаются чаще всего почкованием (см. рис. 2.34) и отчасти остаются в разветвленных цепочках разной длины (рис. 11.30, С); у Schizosaccharomyces клетки размножаются за счет образования поперечных перегородок (рис. 11.30, А). Большинство дрожжей (например, пекарские, винные и пивные1) почкуются с образованием рубца. Дочерние клетки возникают здесь таким образом, что у материнской клетки в одном месте образуется почковидное вздутие. Разрастающаяся дочерняя клетка, куда заходит ядро, отрывается после образования разделительной перегородки от материнской клетки. И у материнской, и у дочерней клетки в месте разделения остается рубен. Каждая дрожжевая клетка имеет один рубец, возникший при ее образовании, и много (до 32) рубцов там, где возникали дочерние клетки. В качестве запасного вещества клетки откладывают гликоген и содержат многочисленные витамины, особенно группы В.2
1 В современных системах делящиеся дрожжи типа Schizosaccharomyces обычно считаются далеко отстоящими в эволюционном плане от почкующихся дрожжей типа Saccharomycesи сближаются с тафриновыми. — Примеч. пер.
2 По современным представлениям, пекарские, винные и пивные дрожжи, ранее рассматривавшиеся как самостоятельные вилы, представляют собой один вид — Saccharomyces cerevisiae. — Примеч. пер.
Пекарские дрожжи (Saccharomyces cerevisiae) были первым эукариотным организмом, у которого была полностью секвенирована последовательность нуклеотидов ДНК. При половом размножении (рис. 11.30) две клетки копулируют друг с другом (иногда через короткий копуляционный мостик). Если смешиваются суспензии дрожжей различных типов скрещивания, происходит выпадение осадка в виде облака. Эта агглютинация обусловлена специфическими белками клеточной стенки одного типа скрещивания и полисахаридами клеточной стенки другого типа скрещивания. Соединение в белково-полисахаридный комплекс является причиной агглютинации. Зигота непосредственно или после промежуточной почкующейся стадии становится сумкой, образуя после мейоза 4 или 8 аско- спор, которые после разрыва стенки получившейся сумки освобождаются и прорастают в новые вегетативные клетки.
Можно выделить три типа процесса развития у дрожжей. При гаплонтном типе (Schizosaccharomyces, рис. 11.30, А — F) ядро зиготы делится мейотически непосредственно после ее образования и зигота превращается в сумку; вегетативное размножение осуществляется в гаплофазе. При гаплодиплонтном типе (Saccharomyces cerevisiae, рис. 11.30, G — L) зигота дает начало почкующимся клеткам, в некоторых из них после мейоза возникают аскоспоры. Таким образом, здесь имеет место промежуточная смена ядерных фаз. Из гаплоидных аскоспор снова развиваются почкующиеся клетки, теперь они гаплоидные. Наконец, при диплонтном типе (Saccharomycodes iudwigii, рис. 11.30, М — S) аскоспоры сливаются попарно друг с другом уже в сумке; вегетативное почкование осуществляется исключительно в диплофазе.
Рис. 11.30. Ascomycètes, Endomycetidae, Saccharomycetales; A — F — Schizosaccharomyces, S. octosporus (350x): A — группа соединенных клеток, В — F — копуляция и образование сумки, G — L— Saccharomyces, S. cerevisiae: G — цепочки почкующихся клеток (200х), Н — L— образование сумки (550х); М — S — Saccharomycodes, S. ludwigii (375х): М — Р — копуляция прорастающих мейоспор в сумке, Q — прорастание диплоидной клетки, R, S — образование аскоспор; T, U — Endomyces, Е. magnusii. копуляция и образование сумки (375х); V — Candida, С. reukaufii (375х); W — Y — Dipodascus, D. albidus, копуляция и образование сумки (275х)
Дрожжи (Saccharomyces) находят разнообразное применение как возбудители спиртового брожения, причем как конечный продукт в одних случаях используется спирт (вино, пиво и т.д.), в других — СО2 (для разрыхления хлебного теста). Если винные дрожжи (Saccharomyces ellipsoideus = S. vini) встречаются и в диком виде на ягодах, то пивные дрожжи (S. cerevisiae и S. carlsbergensis с многочисленными расами) известны только в культуре. При брожении более крупные клетки дрожжей оседают. а клетки меньшего размера остаются парящими в жидкости. В сортах пива «поверхностного брожения» (например, белое пиво) большая часть дрожжевых клеток отводится с пеной (в противоположность нормальным сортам пива «низового брожения»). Используемое в хлебопечении кислое тесто содержит кроме дрожжей также молочнокислые бактерии. Так называемый чайный гриб представляет собой смесь из дрожжей и бактерий; он превращает черный чай в кислый на вкус напиток.
Единственный порядок: Endomycetales. Представители порядка имеют типичное для подкласса строение. Помимо Saccharomyces, это строение можно рассмотреть на примере родов Dipodascus и Endomyces. Все эти роды можно рассматривать как самостоятельные семейства. Dipodascaceae: клетки гиф Dipodascus образуют длинные цепочки. Эти клетки могут быть одно- (D. uninudeatus) и многоядерными (например, D. albidus). У D. albidus, который встречается в слизистых истечениях деревьев, образуются клювовидные гаметангии (рис. 11.30, W). которые сливаются концами и затем отграничиваются в основаниях поперечными перегородками (рис. 11.3. X). Ядра одного гаметангия (♂) переходят в несколько более крупный другой (♀). Слияние, однако, происходит только у одной пары ядер. Женский гаметангий вытягивается в длинную сумку. Не слившиеся ядра отмирают, а из диплоидного ядра слияния после мейоза возникают многочисленные гаплоидные ядра, из которых каждое становится ядром аскоспоры. D. iminudeatus обитает на мертвых насекомых. При его размножении сливаются две соседние клетки, становящиеся таким образом гаметангиями; ядра у них крупнее, чем у остальных клеток. Представители семейств Endomycetaceae и Saccharomycetaceae содержат в сумках не более восьми аскоспор.
Endomyces образует нитевидный мицелий. У Е. magnusii, который обитает в слизистых истечениях (возникающих из экссудата флоэмы) дуба, размеры мужской и женской копуляционных ветвей значительно различаются; последняя после слияния ядер и следующего за ним мейоза становится четырехспоровой сумкой (рис. 11.30, Т, U).
Дрожжевидные талломы встречаются не только у Endomycetidae; они имеются (по крайней мере, на определенных стадиях развития) также у других аскомицетов (Taphrinomycetidae), у базидиомицетов (например, Sporobolomyceta- сеае, Exobasidiaceae, Ustilaginaceae) и у несовершенных грибов (например, Cryptococcaceae).
3. Подкласс: Лабульбениомицетиды (Laboulbeniomycetidae)
Мы лишь вкратце коснемся этой группы грибов — паразитов насекомых, в ходе эволюции рано отделившейся от остальных аскомицетов. Для них характерны редуцированные талломы, но строго фиксированное строение мелких органов спороношения (рис. 11.31). Эти грибы чаще всего проникают лишь короткой, темноокрашенной «ножкой» в хитиновый покров своих хозяев. Многие виды (из общего числа примерно 1 500) очень специфичны в отношении хозяев. Талломы развивают в специальном вместилище («перитеции») — женский половой орган (аскогон) с трихогиной. Он оплодотворяется спермациями (не гаметангиогамия!), которые высвобождаются из мелких бутылевидных сперматангиев. После оплодотворения возникают сумки с нежными стенками, которые содержат одно- или двухклеточные аскоспоры. Время от заражения хозяина до созревания спор составляет лишь 10 — 20 дней. Единственный порядок Laboulbeniales этого подкласса показывает определенные родственные связи с Ascomycetidae; согласно другой точке зрения он занимает очень изолированное положение в системе.
Рис. 11.31. Ascomycètes, Laboulbeniomycetidae. Stigmatomyces baerii. Верхний ряд: развитие до образования сперматангиев (аn) и материнской клетки аскогона (m). Нижний ряд: стадия оплодотворения (аg — аскогон, t — трихогина), наполовину зрелый перитеций с молодыми сумками (а), сумка с 4 двухклеточными спорами (400х)
4. Подкласс: Аскомицетиды (Ascomycetidae)
Представители этого подкласса отличаются нитевидным гаплоидным мицелием в вегетативной фазе, гифами с парами ядер (аскогенными гифами) в генеративной стадии, а также плодовыми телами, в которых перемешаны гаплоидные и дикариотичные гифы. Дикариотичные гифы связаны с ran лоидным мицелием пространственно и в отношении физиологии питания. Группировка ядер в пары происходит после различных процессов оплодотворения.
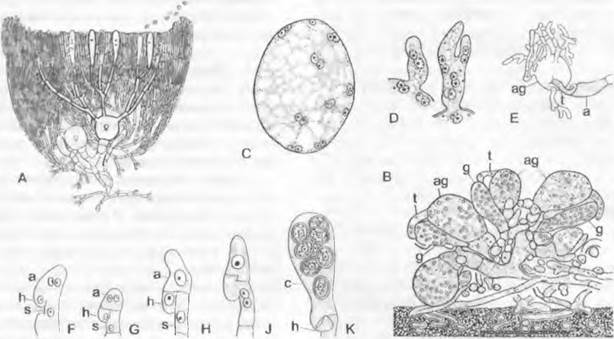
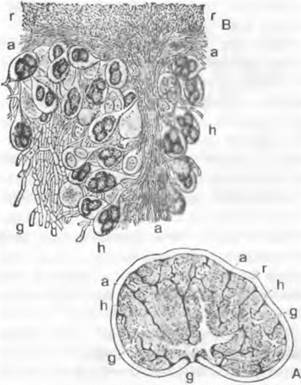
Размножение. При гаметангиогамии, например, у Ругопеmа confinens1, в молодых зачатках плодовых тел на некоторых концах гиф возникают ♀ органы; они состоят из клетки-ножки, расширенного многоядерного ♀ гаметангия — аскогона (рис. 11.32, В: ag; А) и расположенного на вершине аскогона изогнутого многоядерного придатка — трихогины (t). В непосредственной близости от аскогона возникает (также из гаплоидных гиф) булавовидный многоядерный ♂ гаметангий (g). Разные половые органы встречаются иногда группами и растут навстречу друг другу, так что ♂ гаметангий сливается с трихогиной (гаметангиогамия). Трихогина открывается в месте их соприкосновения (ее ядра при этом дегенерируют); ♂ ядра мигрируют из ♂ гаметангия в трихогину и оттуда через временно открывающуюся пору в аскогон (плазмогамия). Там ядра ♂ и ♀ группируются попарно (рис. 11.32, С). После этого из аскогона образуются многочисленные выросты, в которые переходят пары ядер: аскогенные гифы, которые растут, претерпевая клеточные деления, и ветвятся. При всех клеточных делениях в каждой клетке остаются попарно сгруппированные ядра, так как деления ядер происходят конъюгированно (т. е. одновременно). Так возникают клетки дикариофазы с двумя ядрами разных типов скрещивания (здесь ♂ и ♀). У других аскомицетов перенос ♂ ядер в аскогон может происходить не из ♂ гаметангиев, а в результате образования многоядерных или одноядерных конидий (гамето-гаметангиогамия), а также через гаплоидные гифы. При соматогамии гаметангиев нет, а сливаются обычные, особо не дифференцированные гифы. В результате слияния (после плазмогамии) и здесь возникает дикариотичный мицелий. Часто половая функция бывает подавлена. Говорят, о партеногамии, когда внутри аскогона происходит попарная координация ядер без предшествующего оплодотворения ♂ ядрами; об автогамии — когда образование дикариона происходит где-либо без участия аскогонов; об апомиксисе — когда половая функция утрачена и развитие проходит в гаплофазе. Дикариотичные (аскогенные) гифы у аскомицетов часто отличаются наличием у поперечных перегородок своеобразных крючков, которые возникают следующим образом (рис. 11.32, F — J. Растущая конечная клетка образует сбоку, несколько ниже конца гифы (субтерминально), отходящий вниз и в сторону, противоположную направлению роста, крючковидный вырост. Одновременно ядра дикариона делятся, причем одно из образовавшихся дочерних ядер мигрирует в крючковидный вырост. Затем верхняя пара ядер отделяется поперечными перегородками (G), тогда как крючок на конце сливается с основной гифой и находящееся в нем ядро возвращается туда (J). Образование крючка повторяется при каждом новом делении конечной клетки, пока в ней не произойдет кариогамия и не начнется образование сумки (Н). Из конечных клеток аско- генных гиф, после того, как там произойдут кариогамия и мейоз, возникают сумки. Молодые зачатки сумок первоначально двуядерные (рис. 11.32, F, G). После слияния ядер (Н) конечная клетка превращается в булавовидную, первоначально еще одноядерную диплоидную спороцисту. Из ядра слияния после трехкратного деления (имеет место мейоз) возникает 8 ядер, которые в результате свободного деления клеток (см. Ascomycètes) отграничиваются стенками. Таким образом, сумка — это мейоспороциста, в которой к двум делениям мейоза добавляется митоз.
1 Правильное современное научное название этого гриба - Ругоnemа оmphalode. — Примеч. пер.
Рис. 11.32. Ascomycètes, Ascomycetidae: А — плодовое тело однодомного дискомицета (схема), гаплофаза, дикариофаза, диплофаза (крючки не изображены); В — J — Ругопета conftuens: В — зачаток плодового тела (450х), С — попарная группировка ♂ и ♀ ядер в аскогоне, D — переход дикарионов в вырастающие из аскогона аскогенные гифы (С, D 1000х), Е — аскогон с аскогенными гифами (150х), F — J — развитие сумки; К — Bouüiera, молодая сумка (с) с аскоспорами (F — К — 1000х); а — будущая сумка; аg — аскогон с трихогиной t; g — ♂ гаметангии; h — крючок; s — клетка-ножка
Плазма, не используемая при спорообразовании, — периплазма — часто находит применение для отложения еще одного, разнообразно скульптурированного слоя на стенках спор. Сумки развиваются, как правило, внутри плодовых тел, реже — свободно, на незащищенных гифах зачатка плодового тела. Аскогимениальным называют такое развитие, при котором только аскогенные дикариотичные гифы окружаются оболочкой (перидием); таким образом, начало образования плодового тела определяется оплодотворением. При асколокулярном типе развития зачатки плодовых тел или сплетения гиф, которые впоследствии должны превратиться во вместилища сумок, закладываются уже до оплодотворения, а аскогенные гифы врастают в образующиеся после оплодотворения полости (локулы). Систематическое подразделение основывается здесь наряду с прочими признаками на разном онтогенезе и разном строении плодовых тел и сумок. Различают несколько видов плодовых тел: округлое закрытое — клейстотеций; блюдцевидное открытое — апотеций;бутылевидное, с выводным отверстием — перитеций, при аскогимениальном, и псевдотеций при асколокулярном развитии. Псевдотеции могут широко раскрываться или, как клейстотеции, разрываться пассивно.
1. Надпорядок: Eurotianae. Стенки (прототуникатных) сумок недифференцированные, тонкие. Часто они ослизняются до созревания спор, так что аскоспоры освобождаются пассивно.
1.1. Порядок: Eurotiales. Сюда относятся грибы, у которых часто подавлено или отсутствует половое спороношение. Однако характеристика и положение порядка в системе аскомицетов основаны на признаках половой стадии (рис. 11.33, С — Е). Она образуется, например, у Eurotiaceae после слияния булавовидных гаметангиев (аскогона, cf гаметангия). У поперечных перегородок возникающих после этого дикариотичных аскогенных гиф крючки отсутствуют. Булавовидные сумки закладываются внутри закрытых шаровидных плодовых тел. Сумки содержат каждая по 4 или 8 аско- спор, часто дисковидных, и беспорядочно располагаются в большом количестве в плодовом теле, плектенхиматическая стенка которого должна разрушаться, чтобы сумки и аскоспоры могли распространяться. Соответственно плодовые тела здесь — клейстотеций без специально образующегося выводного отверстия. Характерны и бесполые спороношения (рис. 11.33, А, В). Некоторые имеют названия Aspergillus и Pénicillium, они относятся к наиболее частым плесневым грибам («плесень» — не систематическое понятие, а общее обозначение для поверхностно растущих мицелиев грибов). Размножение осуществляется бесполым путем — конидиями (см. бокс 11.3), которые образуются на расположенных близко друг к другу конидиеносцах и часто бывают окрашены в сине-зеленый цвет.
Рис. 11.33. Ascomycetidae, Eurotiales: А — Aspergillus glaucus, леечник, конидиеносец (ЗООх); В — Pénicillium glaucum, «кистевик», конидиеносец (300х): р — фиалида, m — метула; С — Eurotium, спирально закрученный аскогон, охватывающий cf гаметангий (450х); D — Talaromyces, обвивающие друг друга гаметангии (500х); Е— Eurotium, клейстотеций в поперечном разрезэ (250х)
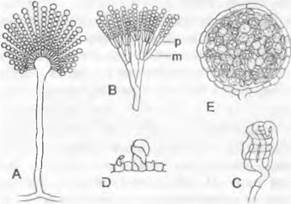
У леечника, или аспергилла (Aspergillus), конидии расположены на конидиогенных клетках (фиалидах), расходящихся во все стороны от шаровидного вздутия на вершине конидиеносиа. Фиалиды последовательно отшнуровывают конидии, которые связаны друг с другом в виде цепочек. У кистевика, или пеницилла (Pénicillium), конидии, также располагающиеся цепочками, возникают на разветвленных конидиеносцах, причем ветви, образующие конидии, называются фиалиды, а расположенные под ними — метулы. Систематические единицы в пределах Eurotiales называют по половой стадии, а когда она отсутствует — по конидиальной стадии. Например, конидиальная стадия у Eurotium, Sartorya— Aspergillus; у Talaromyces, Carpenteles — Pénicillium.
К отдельному семейству относятся обитающие под землей (гипогейно) оленьи трюфели — Elaphomyces (Elaphomycetaceae).Их клубневидные плодовые тела размером 1 — 4 см для человека несъедобны, однако выкапываются и съедаются дикими животными; таким путем распространяются споры.
Применение и наносимый ущерб. Из Pénicillium по fatum, P. chrysogenum1 и других видов получают антибиотик пенициллин, подавляющий синтез клеточной стенки бактерий. Антибиотик выделяется грибом в питательную среду ergillus P. roquefortii и P. camembertii необходимы для изготовления определенных сортов сыра. Asp. wentii продуцирует амилазы и протеазы и поэтому используется в биотехнологии; Asp. flavus образует афлатоксины, которые вызывают рак и повреждения печени. Asp.fumigatus вызывает заболевания легких и бронхов у человека. Другие опасные возбудители грибных заболеваний человека и животных — микозы — также относятся к этому порядку или должны быть отнесены сюда же на основании единственно известных анаморф, сходных с конидиальными стадиями Eurotiales.
1 По современным представлениям, Р. notatum и Р. chrysogenum — один вид (для которого должно быть оставлено второе название). — Примеч. пер.
К прототуникатным аскомицетам относятся и другие, меньшие порядки, перечисленные ниже.
1.2. Порядок: Microascales. К порядку относятся возбудители широко известного отмирания вязов из рода Ophiostoma. Гриб обитает в личиночных ходах жуков-заболонников, поражающих вязы. Длинные шейки их перитециев далеко выступают в личиночные ходы. Анаморфы развивают конидиеносцы. Аскоспоры и конидии собираются в каплевидных выделениях, которые захватываются личинками жуков и распространяются. Отмирание вязов представляет собой болезнь увядания, вызываемую закупоркой сосудов молодых ветвей и выделением токсина, убивающего листья (нератоульмина). Только после заноса вирулентного штамма из Север
ной Америки (Ophiostoma novo-ulmi) болезнь распространилась в виде эпифитотии и по Европе. Другие порядки — Onygenales (Onygena equina на копытах лошадей), чаще всего образующие лишайники Caliciales, а также Coronophorales и Меliolales.
Стенка сумки у всех последующих порядков эутуникатная, т. е. четко различима как более толстый слой, прочная и снабженная приспособлениями для выброса ас- коспор. Сумки первоначально однослойные (у порядков 2 — 4 — унигуникатные).
2. Надпорядок: Erysipliales. Эти паразитические грибы, как и возбудители настоящей мучнистой росы, живут на растениях. Пораженные растения выглядят как бы обсыпанные мукой. Такое впечатление создается из-за белого поверхностного мицелия, который летом образует в больших количествах конидии (рис. 11.34, А). Через гаустории, которые проникают в клетки эпидермиса хозяина (A: h), фиб получает питательные вещества. Половое спороношение представляет собой мелкие клейстотеции от коричневого до черного цвета, видимые невооруженным глазом как точки поверх белого налета мицелия и конидиального спороношения.
Половое размножение. Мужская копуляционная ветвь, которая делится на клетку-ножку и одноядерный ♂ гаметангий. прикладывается к также одноядерному аскогону (рис. 11.34, D). Половое ядро ♂ переходит в аскогон (Е). После этого из каждого оплодотворенного аскогона возникает сумка без промежуточного образования аскогенных гиф или же аскогон (рис. 11.35) дает начало аскогенным гифам, конечные клетки которых образуют сумки. В первом случае пара ядер после синхронного деления (G) сливается в диплоидное ядро зиготы, которое после мейоза дает от 4 до 8 ядер аскоспор. Во втором случае развитие сумок соответствует описанному выше нормальному варианту, но поперечные стенки аскогенных гиф не имеют крючков.
Рис. 11.34. Ascomycetidae, Erysiphales: А — Uncinula necator образование конидий (100х); В — то же, клейстотеций с придатками (30х); С — Microsphaera alphitoides, клейстотеций с придатками (30х); D — G — оплодотворение у Sphaerotheca fuliginosa (250х); а — аскогон; g — ♂гаметангий; h — гаусторий; hh — покровные гифы; к — конидия; s — клетка-ножка
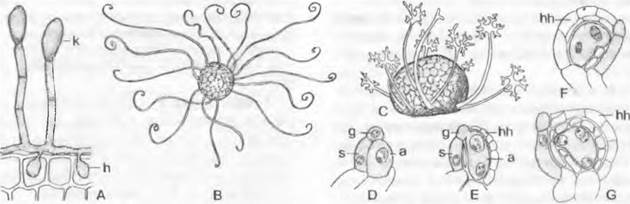
Одновременно с образованием и оплодотворением аскогона он оплетается гифами оболочки, которые в конечном счете образуют светлый основной слой и темный перидий клейстотеция. Последний при созревании разрывается трещиной под давлением набухающих сумок. Чаше всего у основания клейстотеция возникает венец из гиф, часто на конце дихотомически ветвящихся или крючковидно загнутых. Они, по- видимому, способствуют распространению гриба (В, С). Сумки в клейстотеции расположены пучком (если сумка не одна) и открываются (иногда небольшой крышечкой), отбрасывая аскоспоры в воздух на расстояние до 2 см.
Рис. 11.35. Ascomycetidae, сумки до и после выбрасывания спор: А — С — иноперкулятные сумки, вершины сумок с апикальными аппаратами до (В) и после (С) выхода спор; D — оперкулятная сумка, отверстие с крышечкой
Uncinula necator (A, В) поражает листья и ягоды винограда (конидиальная стадия: Oidium tucken). Sphaerotheca mors-uvae (с одной сумкой в клейстотеций) поражает крыжовник; Sphaerotheca pannosa — розы; Microsphaera alphitoides (С) обитает на листьях дуба. Blumeria graminis (Еrysiphe grаmmis) — паразит дикорастущих и хлебных злаков. Для борьбы с «настоящей мучнистой росой» применяются препараты серы.
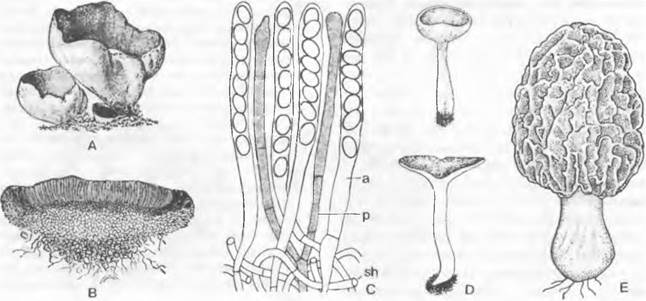
3. Надпорядок: Pezizales. Этот очень разнообразный по особенностям развития и строения порядок насчитывает примерно 1 000 видов, в основном сапротрофов. Типичная форма плодового тела у представителей Реzizaceae и близких семейств — чашевидный до дисковидного апотеций (например, Реzizа), на верхней стороне которого в виде палисадного слоя расположен гимений, состоящий из сумок и гаплоидных стерильных парафиз. Сумки открываются на вершине крышечкой; таким образом, они унитуникагные — онеркулятные, как в предыдущем порядке (рис. 11.35). Споры часто отбрасываются на большие расстояния (Dasyobolus см. 8.3.2.7).
Оплодотворение и образование сумок было впервые открыто и детально изучено у Рyronema (см. рис. 11.32). Р. соnfluens образует относительно мелкие дисковидные плодовые тела. Они часто плотно прижаты друг к другу, образуя почти корковидные налеты на местах, подвергшихся воздействию огня, или на почве. Уже перед копуляцией половые органы оплетаются рыхлым слоем гаплоидных покровных гиф. После оплодотворения (плазмогамии) возникают аскогенные гифы; монокариотичные гаплоидные гифы и дикариотичные аскогенные гифы, чаще всего образующие крючки, переплетаются и совместно образуют плодовое тело.
Образование плодового тела связано с половым процессом, который происходит одновременно или в заранее возникающих зачатках плодовых тел. Г имений у Ругоnета развивается с самого начала открыто на поверхности плодового тела (гимнокарпный тип). У других родов (например, Ascophanus) гимений возникает сначала внутри шаровидного зачатка плодового тела, поверхностный слой которого позже наверху разрывается, так что гимений становится открытым (гемиангиокарпный тип). Размер плодового тела в зависимости от вида — от нескольких миллиметров до более одного дециметра (Sarcosphaera).
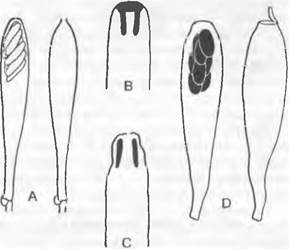
Некоторые представители обладают более крупными апотециями на ножке. Ножка может иметь продольные бороздки (например, Helvetia), а поверхность самого апотеция, края которого отогнуты вниз, может быть подразделена на отдельные камеры (например, Morchella). Увеличение площади гимения и подъем его на ножке обеспечивают более действенное распространение спор. У некоторых родов (например, Helvetia, Gyromitra) не образуется ни аскогонов, ни ♂ гаметангиев. Сливаются друг с другом вегетативные гифы совместимых типов скрещивания (соматогамия). У Morchella часто встречается слияние гиф только одного и того же вида (автогамия) (рис. 11.36).
Рис. 11.36. Ascomycetidae, Pezizales: A — Otidea (Peziza) leporina (2/Зх); В — Pulvmula convexula, разрез через апотеций; на верхней стороне расположен гимений (20х); С — Morchella esculenta, плодовое тело (3/4х); D — Helvella pezizoides (3/4х); Е — Morchella esculenta (3/4х); а — сумка; р — парафиза; sh — субгимениальное сплетение гиф
Чаще всего развивающиеся в лесных почвах плодовые тела настоящих трюфелей (Тиbегасеае) могут быть выведены из открытых блюдцевидных форм и отнесены к Реzizales в связи с наличием переходов. Однако их плодовые тела остаются в почве и замкнуты (гипогейный образ жизни). Освобождение аскоспор происходит в значительной мере за счет питающихся этими грибами животных или при разрушение плодового тела. Плодовые тела, чаще всего имеющие клубневидную форму, пронизаны ходами, которые, по крайней мере на ранней стадии развития, выходят наружу и покрыты своего рода гимением (см. рис. 11.37). Эти ходы представляют собой впячивания гимения внутрь. В широких булавовидных сумках, которые возникают после соматогамии (автогамии) из конечных клеток аскогенных гиф, несущих пряжки (см. Нуmеnоmусеtidае), располагается 1 — 5 скульптурированных аскоспор коричневого цвета. Исходно оперкулятное строение сумок почти не различимо, так как их нежная стенка не дифференцирована.
Рис. 11.37. Pezizales, Tuberaceae, Tuber rufum: A — плодовое тело, вертикальный разрез (Зх); В — разрез части гимения (300х); а — темные вены из плотного сплетения гиф; g — рыхлая плектенхима; h — гимений; г — кора
Более крупные представители Pezizales (например, Morchella, рис. 11.36, С) находят применение как съедобные грибы. Известны также и ядовитые виды. К ним относится Sarcosphaera crassa. Строчок (Gyromitra esculentа), яд которого разрушается при нагревании, иногда используется в пищу после отваривания (отвар сливают), однако из- за опасности отравления использовать его в пищу не рекомендуется. Из трюфелей, живущих в симбиозе с лесными породами деревьев, несколько видов (особенно Tuber magnatum — пьемонтский трюфель и Tuber melanosporum — перигорский трюфель) издревле высоко ценились как съедобные грибы благодаря летучим ароматическим веществам, придающим блюдам из них особую привлекательность. Подземные плодовые тела находят с помощью специально натасканных для этого свиней или собак. Трюфели имеют чрезвычайно высокую цену на рынке («черные бриллианты»)1.
1 1 кг перигорского трюфеля стоит в Европе порядка 1 500 евро. — Примеч. пер.
4. Надпорядок: Leotianae. На вершине унитуникатной и иноперкулятной сумки (см. рис. 11.35) образуется отверстие в виде поры, окруженное или простым расплывающимся валиком, или дополнительно апикальным кольцом, или верхушечным валиком. Последние при использовании реактива, содержащего йод, часто окрашиваются в синий цвет и тогда получают название амилоидных. Механизм выбрасывания спор, вызываемого светом или изменением влажности, полностью не выяснен. По- видимому, разбухание валика или же апикального кольца играет при этом решающую роль, как и тургор внутри сумки. Плодовые тела — преимущественно апотеции (4.1-4.2).
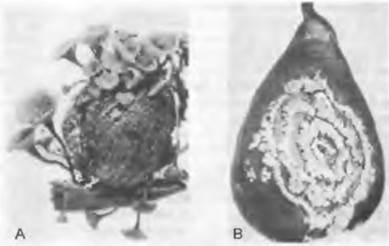
4.1. Порядок: Leotiales. Эти грибы имеют плодовые тела от бокаловидной до блюдцевидной формы, возникающие аскогимениальным путем, однако их размеры, а также размеры сумок и спор часто меньше, чем у Pezizales. Наряду с типичными апотепиями здесь встречаются, как и у Pezizales, другие производные формы плодовых тел. Например, булавовидные — у Trichoglossum, блюдцевидные на ножках — у Sclerotinia или дифференцированные на шляпку и ножку — у Cudonia (результат конвергентной эволюции плодовых тел с Pezizales и т. д.). Большинство видов многочисленных родов живет сапротроф- но, однако некоторые — паразиты: например, Trichoscyphella willkommii— возбудитель рака лиственницы, Pseudopeziza trifolii— возбудитель болезней клевера. Sclerotinia fructigena поражает яблоки и груши; сначала развиваются конидиальные пустулы, часто расположенные в виде концентрических окружностей (что обусловлено ежедневными сменами светового и темнового периодов) со спороношениями типа «Monilia», а весной на мумифицированных плодах возникают апотеции на длинных ножках (рис. 11.38). Конидиальная стадия гриба Sclerotinia fuckeliana, называемая Botrytis cinerea, в дождливые годы вызывает опадание ягод винограда, а при сухой погоде — «благородную гниль» ягод, особенно при большом содержании в них сахара. Обитающий сапротрофно на гнилой древесине Chlorosplenium aeruginosum окрашивает субстрат в интенсивно сине-зеленый цвет.
Рис. 11.38. Ascomycetidae, Leotiales, Sclerotinia fructigena: А — плодовое тело на мумифицированном персике (3/4х); В — конидиальная стадия Monilia на груше. Мицелий образует конидии концентрическими кругами (1/2х)
4.2. Порядок: Phacidiales (включая Rliytismatales). Эта группа, ранее объединявшаяся с предыдущим порядком, отличается от него ас- колокулярным развитием плодового тела. Плоские плодовые тела открываются трещиной или продольными разрывами. Представители порядка Рhacidiales ведут преимущественно паразитический образ жизни. Сюда (Rhytismatales) относятся возбудитель черной пятнистости клена, Rhytisma acerinum, который вызывает осенью черные пятна на листьях (апотеции образуются весной), и Lophodermium seditiosum на сосновых иглах, вызывающий болезнь «шютте».
4.3. Порядок: Lecanorales. Представители порядка составляют основную часть грибов, входящих в состав лишайников наших широт, и поэтому будут рассмотрены в соответствующем разделе (см. приложение 2 к Mа). Грибы, живущие в лишайниковом симбиозе, образуют апотеции, которые между парафизами (см. рис. 11.36, С) содержат особого строения сумки с утолщенными вершинами. Эти сумки булавовидные, толстостенные, иногда их стенка состоит из нескольких слоев (причем слои, в отличие от тех, что наблюдаются у битуникатных сумок, имеют одинаковую эластичность) и имеют вокруг поры на вершине верхушечный валик, окрашивающийся йодом в синий цвет.
Последующие порядки (4.4 — 4.7) характеризуются бутылевидными плодовыми телами (перитециями) с направленным вверх выводным отверстием (оstiolum) на вершине; тип развития у них аскогимениальный. Сумки совместно с многочисленными гаплоидными гифами (парафизами) образуют палисадный плодущий слой (гимений), который выстилает основание и стенки заранее образующейся полости плодового тела (рис. 11.39, А). При созревании одна сумка за другой по очереди выдвигается до тех пор, пока ее вершина не войдет в выводное отверстие перитеция, после чего сумка сразу выбрасывает все 8 спор. Высота, на которую они выбрасываются, может составлять 20 см и более. После этого сумка спадается, так что отверстие перитеция освобождается для следующей сумки.
Рис. 11.39. Ascomycetidae: А, В — Sphaeriales: А — Podospora fimiseda, перитеций (90х); В — нектриозный рак на ветке плодового дерева (возбудитель — Nectria galligena) (в натур. величину); С — Xylariales, Xylaria hypoxylon (натуральная величина); а — сумки; kb — область конидиального спороношения; m — гифы мицелия; р — перифизы; pb — область перитециев
4.4. Порядок: Sphaeriales1. Тупые наверху сумки представителей этого порядка имеют в верхней части сумки образующийся вокруг апикальной поры валик; апикальный аппарат выглядит чаще всего как замок поры в форме пластинки. Как представитель этого порядка может быть упомянута прежде всего Neurosрога. N. sitophila и N. сrassa вызывают красную хлебную плесень и выносят высокие температуры (до 75 °С). Виды Neurospora образуют на каждом отдельном мицелии как аскогоны, так и клетки, которые предназначены для переноса ♂ ядер. В качестве переносчиков ♂ ядер на трихогину аскогона служат многоядерные конидии (мегаконидии), одноядерные спермации или микроконидии, а также соматические гифы. Спермации представляют собой специализированные клетки, предназначенные для этой цели; мега- и микроконидии могут также образовывать проростковую трубку и соответственно давать начало новому мицелию. Перекрестное оплодотворение достигается за счет того, что аскогоны могут оплодотворяться только ядрами противоположного типа скрещивания (гомогенная несовместимость; см. рис. 11.19). После оплодотворения (плазмогамии) из аскогона вырастают обычным путем дикариотичные аскогенные гифы, образующие крючки.
1 В настоящее время в литературе этот порядок чаще разделяется на несколько порядков: Sordariales, Xylariales, Нуpocreales и др. — Примеч. пер.
Большое значение для генетических исследований приобрели обитающие в природе на навозе Роdоsрога аnserinа, Sordaria fimicola и S. mасгоsрога. Сумки этих видов Sordariaсодержат по 8 аскоспор, тогда как у Р. аnserina они 4- споровые. У Р. anserina мегаконидии отсутствуют; ♂ ядра переносятся спермациями. У обоих упомянутых видов Sordaria мега- и микроконидии отсутствуют, аскогоны не образуют трихогины, а группировка ядер в дикарион осуществляется партеногенетически; таким образом, перитеции развиваются в результате самооплодотворения. Правда, у мутантов может происходить также соматогамия между разными мицелиями. В области конфронтации возникают в этом случае перитеции как результат скрещивания, в их сумках может наблюдаться предредукция или же посгредукция.
Перитеции Sphaeriales, часто едва достигающие размера 0,5 мм. располагаются обычно поодиночке, но иногда и группами, которые могут находиться на более или менее четко выраженном сплетении (строме). Строма — это, как правило, твердое образование типа склероция, в которое погружены отдельные перитеции или их группы (тогда это образование представляет собой как бы комплексное плодовое тело). Перитеции Nectria cinnabarina сидят на подушковидных киноварно-красных стромах; последние развивают сначала конидии, позже перитеции; в обеих формах они видны как красные подушечки на отмерших ветвях. Nectria galligena (с бесцветными конидиями) обитает как паразит в коре и вызывает рак плодовых деревьев. Отмершие под влиянием гриба части коры обрастают раневым каллусом, который, в свою очередь, сам отмирает, что приводит к беспорядочным волнистым разрастаниям (рис. 11.39, В) и может, наконец, погубить все дерево. Из культуральных фильтратов паразитирующей на растениях риса Gibberella было выделено ростовое вещество гиббереллин (см. 7.6.3). Из этого же порядка можно еще упомянуть роды Cochliobolus (см. 9.3.3), Daldinia, Gaeumannomyces и Hypoxylon.
4.5. Порядок: Diaporthales. Пора сумки в дополнение к апикальному валику окружена оптически плотным кольцом, которое хорошо красится анилиновым синим. Сумки часто растворяются и под конец вместе со спорами выдавливаются из выводного отверстия плодового тела. В остальном этот порядок, представленный родами Diaporihe, Diaporthella и др., очень сходен с предыдущим. Перитеции возникают внутри строматического сплетения гиф. Виды, многие из которых — паразиты, внедряются своими стромами в ткань хозяина, а из нее обычно несколько выступают наружу только длинные шейки перитециев. Endothia parasitica1 была занесена в Северную Америку и практически полностью уничтожила на всем материке большие массивы настоящего каштана (Castanea dentata, см. 13.8). На территории Европы (например, в Тичино) ущерб от этого гриба также очень велик.
1 В настоящее время в литературе этот вид чаще встречается под названием Cryphonectria parasitica. — Примеч. пер.
4.6. Порядок: Xylariales. Стромы представляют собой более крупные образования в форме подушек, шаров, булав или оленьих рогов, в которые погружены многочисленные перитеции. У часто встречающегося на пнях лиственных пород гриба Xylaria hypoxylon (рис. 11.39, С), имеющего стромы в виде оленьих рогов, в верхней белой части сначала образуются конидии, а позже, в нижней черной части, — перитеции. Апикальный аппарат по строению близок к представителям предыдущего порядка, с тем отличием, что апикальное кольцо на вершине сумки Xylariales красится йодом в синий цвет.
4.7. Порядок: Clavicipitales. Сумки имеют на вершине оптически более плотное расплывающееся кольцо от полукруглой до почти круглой формы. Септированные аскоспоры нитевидные, длинные, а сумки соответственно узкие. Перитеции погружены в стромы, у некоторых представителей дифференцированные на стерильную «ножку» и фертильную «шляпку». В двух последних порядках формирование стромы достигает, таким образом, наивысшего уровня аналогично развитию отдельных плодовых тел (например, у Leotiales); по своему внешнему облику одиночные или «комплексные» плодовые тела часто бывают, несмотря на различия в деталях морфологии и путях эволюционного развития, очень сходны (аналогичная конвергенция наблюдается, например, между Trichoglossum из Leotiales и Cordyceps из Clavicipitales).
Гриб спорынья, Claviceps purpurea, обитает как паразит в молодых завязях злаков и образует там конидии (рис. 11.40, А, В). Выделяемая при этом сахаристая жидкость (медвяная роса) привлекает насекомых, которые переносят конидии на другие цветки. После истощения тканей завязи мицелий превращается в склероций (см. бокс 11.3): гифы плотно срастаются между собой и (прежде всего на периферии) путем поперечных делений образуют псевдопаренхиму (В). Черные снаружи, выступающие из чешуи, твердые склероции (С, D) называют «рожками». Они падают на землю, зимуют и ко времени цветения злаков прорастают в стромы, имеющие облик красноватых головок на ножках. В эти головки погружены многочисленные перитеции (Е). Образование перитециев начинается с копуляции становящихся к тому времени многоядерными аскогонов и ♂ гаметангиев (аскогимениальный тип!). Длинные сумки содержат 8 спор (F). которые переносятся ветром на рыльца пестиков злаков.
Рис. 11.40. Ascomycetidae, Clavicipitales, Claviceps ригригеа: А — пораженная завязь ржи (15х); внизу начинающееся образование склероция, над ним мицелий с конидиями, вверху остатки рыльца; В — образование конидий (300х); С — колос ржи со зрелыми склероциями (2/3х); D — проросший склероций с головчатыми стромами (2х); Е — продольный разрез через строму с многочисленными перитециями (25х); F — сумка и аско- спора (400х)
Виды рода Cordyceps паразитируют на организмах с хитиновыми покровами, например, на грибах с подземными плодовыми телами, таких как Elaphomyces, или на насекомых, которые после заражения заползают в почву. Поднимающиеся из земли булавовидные стромы содержат в своей верхней части многочисленные перитеции. Нитевидные споры уже в сумке становятся за счет поперечных делений многоклеточными и распадаются на отдельные фрагменты. Epkhloé typhina паразитирует на злаках; первоначально белая, затем желтая строма этого гриба охватывает соломину и образует конидии, а позже перитеции. С Clavicipitales родственны грибы из порядков Ostropales и Graphidales, которые чаще всего образуют лишайники.
Применение и вредоносность. Склероции Claviceps ригригеа содержат ядовитые алкалоиды (эрготамин, эрготоксин): при употреблении в пищу зараженного ими зерна происходят сильнейшие отравления («злые корчи», «антонов огонь»)1. Вместе с тем эти вещества находят применение в гинекологии, прежде всего, как средства, способствующие родовым схваткам (отсюда их немецкое название Mutterkorn -«материнское зерно»). Склероции для медицинских целей выращиваются в широком масштабе, например, путем специального заражения ржи; в настоящее время налажено получение алкалоидов из мицелия в ферментерах. Из конидиальной стадии вида Согdусерs (называемой Tolypocladium inflatum) был выделен циклоспорин, который, в частности, применяется при пересадке органов для предупреждения их отторжения.
1 Эти названия связаны с тем, что болезнь встречается в двух формах: гангренозной («антонов огонь») и конвульсивной («злые корчи»). — Примеч. пер.
5. Надпорядок: Dothideales. Стенка сумкиу представителей порядка состоит из двух слоев, по-разному растяжимых; таким образом, в отличие от ранее рассмотренных порядков, она битуникатная. Внешний тонкий слой относительно нерастяжим и разрывается на вершине при увеличении тургорного давления внутри сумки. Толстая внутренняя, растяжимая оболочка сумки после этого сильно вытягивается, причем из-за нарастающего давления аскоспоры одна за другой выбрасываются наружу, сначала замыкая собой пору на вершине (рис. 11.41, С). Повышение осмотического давления внутри сумки обусловлено превращением осмотически неактивных веществ в активные (возможно, гликогена в сахар). Бутылевидные плодовые тела с образующимся в них выводным отверстием чаще всего возникают по асколокулярному типу (см. Ascomycetidae, введение). Будучи внешне сходными с перитециями, из-за различия в характере развития эти плодовые тела получили название псевдотеции (рис. 11.41).
Рис. 11.41. Ascomycetidae, Dothideales: А — Е — Pyrenophora, Р. scirpi, способ вскрытия битуникатной сумки (400х): А — зрелая сумка с 8 четырехклеточными спорами, В — то же, внешняя стенка сумки разорвалась, внутренняя растягивается, С — последняя спора непосредственно перед выбросом, D — освободившаяся сумка, Е — прорастающая спора; F— Н — Мусоsphaerella, М. tulipifera (175х), развитие псевдо- теция; F — ранняя стадия с разветвленным ас- когоном, G — с сумками разного возраста, Н — зрелый псевдотеций
К этому порядку относятся возбудители многих болезней растений. Виды рода Venturia (конидиальная стадия: Fusiclodium) вызывают паршу яблок и груш, образуя темные пятна на опавших или находящихся на дереве плодах (рис. 11.42). Виды рода Сарnodium вызывают буро-черный налет — («чернь») на листьях; будучи сапротрофами, эти грибы используют для питания выделения листьев или тлей. Виды рода Herpotrichia в альпийской зоне обволакивают покрытые снегом ветви хвойных деревьев буро-черным сплетением гиф и вызывают отмирание хвои. Сородичами Dothideales являются и виды, образующие лишайники, в том числе относящиеся к порядку Verrucariales.
Рис. 11.42. Ascomycetidae, Dothideales: парша на груше, вызываемая Fusicladium (по Kirchner и Boltshauser)

Обзор аскомицетов. Среди образующих плодовые тела аскомицетов (Аscomycetidae) в параллельных эволюционных рядах (параллелизм, см. 4.1.1), часто с использованием разных принципов строения, выработались сходные формы, обеспечивающие активное распространение спор, например, булавовидные, бутылевидные плодовые тела или плодовые тела, состоящие из ножки и шляпки. Разные эволюционные линии четко представляют аскогимениальный (перитеций) и асколокулярный (псевдотеций) типы развития. Если не брать во внимание уклоняющиеся Laboulbeniomycetidae и Еndomycetidae, аскомицеты показывают принципиальную общность циклов развития. Половой акт часто осуществляется в аскогонах путем принятия ♂ ядер. Половые ядра партнеров по скрещиванию сразу не сливаются, а сгруппированные парами переходят в аскогенные гифы, размножаются здесь путем сопряженного деления и сливаются в диплоидное ядро только в зачатке сумки (= конечных клетках аскогенных гиф). Плаз могамия и кариогамия разделены далеко в пространстве и во времени двухъядерной стадией (дикариофазой). Дикариотичные клетки функционально уже диплоидны, только их ядра еще индивидуализированы. Типичные аскомицеты — гаплонты с вклинивающимся в жизненный цикл дикарионтом, питание которого, однако, остается зависимым от предшествующего гаплоидного поколения (исключение: тафриномицетиды). Еаплоидный мицелий, чаще всего образующий ♂ гаметангии и аскогоны, может рассматриваться как гаметофит, дикариотичные аскогенные гифы — как спорофит. Фаза дикариотичного спорофита заканчивается образованием сумок (мейоспороцист); в них после слияния ядер и редукционного деления формируются гаплоидные мейоспоры (= аскоспоры). Обычно в каждой сумке возникает 8 аскороспор, однако некоторые виды обладают 1-, 2-, 4- или многоспоровыми сумками. Плектенхима плодового тела состоит из сильно развитых гаплоидных гиф гаметофита. в которые вплетены дикариотичные гифы. Аскогоны и ♂ гаметангии или дающие ♂ ядра клетки (т. е. мегаконидии, микроконидии, соматические гифы) образуются на одном и том же мицелии (однодомность; см. рис. 11.32, В). Самооплодотворению часто препятствует биполярная гомогенная несовместимость (см. рис. 11.19). Среди аскомицетов можно проследить редукцию гаметангиев. Редукция или полная утрата половой функции может наблюдаться у партеногенетических или автогамных видов, которые могут быть выведены от форм, размножающихся половым путем.
Филогения и родственные связи. Вопрос о происхождении аскомицетов остается спорным. Происхождение их от красных водорослей (Rhodophyta) в настоящее время практически уже не обсуждается (его рассматривали ранее на основе сходства формы аскогона и карпогона и деталей развития после оплодотворения).
Отличия от красных водорослей проявляются в отсутствии ассимиляционных пигментов и, что еше более важно, в наличии дикариофазы, в химизме клеточной стенки, а также в аппарате Гольджи. Против признания такого происхождения аскомицетов может говорить и тот факт, что Rhodophyta как очень специализированные водоросли встречаются только с пермского периода. тогда как аскомицеты в формах, соответствующих современным таксонам, известны уже с карбона.
За выведение аскомицетов из грибов со свойствами, характерными для современных зигомииетов и хитридиомицетов, говорят уже имеющаяся у них выраженная или частично сходная гаметангиогамия и в отдельных случаях на мечающаяся дикариофаза, состав клеточной стенки из хитина и соответствующих глюканов (с (5-1,3-, р-1,4-, Р-1,6-связями), синтез лизина через аминоадипиновую кислоту, остающаяся интактной ядерная оболочка при делении ядра. Утрата активно передвигающихся репродуктив ных клеток у аско- и зигомицетов в отличие от хитридиомицетов может быть связана с наземным образом жизни. В таком же плане могут быть истолкованы последовательное усовершенствование плодовых тел и септированность гиф. По морфологическим признакам к аскомицетам ближе зигомицеты. по биохимическим - хитридиомицеты.
Выведение аскомицетов от низших грибов делает рациональным такую их классификацию, при которой тафриномицетиды и эндомицетиды стоят в начале системы. Правда, остается еще в значительной мере сомнительным, можно ли выводить слияние копулирующих клеток, которое можно наблюдать у Saccharomycetales, из зигогамии зигомицетов. Признаки Еndomycetidae, прежде всего отсутствие дикариофазы, свидетельствует в пользу признания их обособленности, которой иногда придают столь большое значение, что их выделяют из аскомицетов в самостоятельный класс. У некоторых Еndomycetidae зигота прорастает в небольшой диплоидный мицелий, и только на нем образуются сумки. Остальные группы аскомицетов, у которых появилась задержка слияния ядер и соответственно более или менее выраженная дикариофаза, отделились в ходе эволюции достаточно рано.
Это обособление могло произойти от предков с признаками Тaphrinomycetidае, которые, однако, по сравнению с Аscomycetidае, имеют более простое строение (отсутствие плодовых тел), но уже обладают дикариофазой (дикариотичный мицелий самостоятелен в плане питания), как все другие аско- и базидиомицеты. Наличие у них дрожжевидных стадий в ходе развития свидетельствует еще о родственных связях с Еndomycetidае. Отсюда выводятся эволюционные линии, с одной стороны, к остальным аскомицетам, с другой стороны — к Ustomycetidае из базидиомицетов. Если у последних дикариотичный мицелий самостоятелен, то у обеих первых названных групп грибов он находится в физиологической зависимости от гаплоидного первичного мицелия. В основание системы аскомицетов должны быть поставлены (после грибов с особенностями Таphrinomycetidае) формы с признаками современных Euro6ales и низших Pezizales (Ascomycetidae). Дальнейшая эволюция шла здесь прежде всего по пути усовершенствования сумок и плодовых тел.
4. Класс: базидиомицеты (Basidiomycetes)
Для базидиомицетов (к ним относится около 30 000 видов т. е. 30% всех грибов) характерна мейоспороциста — базидия, которая в типичном случае отшнуровывает от себя 4 раздельно расположенных мейоспоры. В базидии, как и в сумке, непосредственно друг за другом следуют кариогамия и мейоз (см. рис. 11.52). Возникшие в результате редукционного деления гаплоидные ядра (обычно их 4) мигрируют в кончики палочковидных выростов базидии (стеригмы, см. рис. 11.52, 9) и только здесь происходит «экзогенное» образование базидиоспор. Жизненный цикл, отчасти различающийся в разных группах базидиомицетов, протекает по следующей, приблизительно соответствующей шляпочным грибам, схеме: базидиоспоры прорастают (см. рис. 11.45) в мицелий с одноядерными клетками, способный практически к неограниченному росту; гаметангии, как и у эволюционно продвинугых аскомицетов, не образуются. Если встречаются мицелии противоположных типов спаривания (например, «+» и «-»), то две соприкоснувшиеся вегетативные клетки сливаются между собой (соматогамия, см. рис. 11.52, 3), причем их ядра попарно лишь сближаются, но не сливаются между собой. Возникающая таким образом дикариотичная клетка дает начало дикариотичному мицелию, способному к самостоятельному питанию и не зависящему от гаплоидных и монокариотичных гиф.
Кроме того, базидиомицеты отличаются от аскомицетов и строением поры в поперечной перегородке гифы. Если у аскомицетов это всегда простое отверстие, то у базидиомицетов, у которых также встречаются простые поры, по большей части возле поры имеется расширение в виде бочонка («долипора», лат. dolium — сосуд для вина), покрытое с каждой стороны парентосомой (греч. parenthesis — вдвинутый; soma — тело), которая образуется эндоплазматической сетью (рис. 11.43). Клеточная стенка базидиомицетов показывает слоистую ультраструктуру (ср. Ascomycètes, см. введение).
Рис. 11.43. Basidiomycetes. Поперечные перегородки гифы базидиомицетов: А — с простой порой; В — с долипорой: d — долипора; р — парентосома; w — клеточная стенка
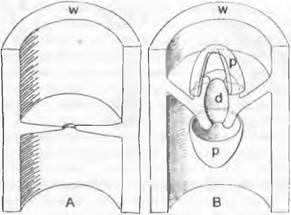
Базидия может быть септированной (фрагмобазидия, рис. 11.44, А, В) или булавовидной и одноклеточной (холобазидия, рис. 11.44, F). Соответственно этому различию в строении раньше разделяли фраг- мобазидиомицеты и холобазидиомицеты.
Рис. 11.44. Basidiomycetes. Формы базидий (750х). A — Platygloea (Auriculariales); В — Bourdotia (Tremellales); С — Tulasnella (Tulasnellales); D — Dacrymyces (Dacrymycetales); E — Sistotrema (Poriales); F — Hyphoderma (Pönales); G — Exobasidium (Exobasldiales); H — Xenasma (Proto- hymeniales); J — Repetobasidium (Poriales); К — Scleroderma (Boletales)
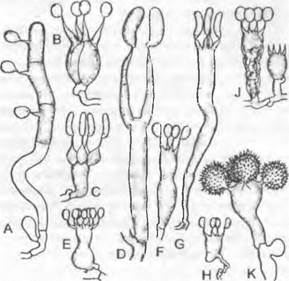
Представители двух первых подклассов, которые рассмотрены ниже, раньше объединялись в подкласс Heterobasidiomycetidae. Однако, согласно молекулярным и ультраструктурным признакам, предлагается другая группировка, отчасти с другим объединением представителей (например, Auriculariales, Tremellales, Microbotryales). Поперечные стенки гиф часто имеют простые поры (см. рис. 11.43, А); пряжки (см. Hymenomycetidae) у значительного большинства отсутствуют.
1. Подкласс: Устомицетиды (Ustomycetidae)
В этом подклассе объединяются облигатные паразиты растений. У тех представителей, у которых базидия разделена поперечными перегородками (фрагмобазидия), предшествующая дикариофаза не размножается дикариотичными спорами. Виды этого подкласса — возбудители головни различных растений («головневые грибы»). Такие названия и заболевания, и грибы получили потому, что пораженные части растений выглядят как бы обугленными.
1. Порядок: Тilletiales. У базидий отсутствуют поперечные перегородки. Базидии образуют на вершине чаще всего 4 или 8 удлиненных мейоспор. Только для разграничения с «головневыми спорами» образуются одна или несколько септ (рис. 11.46, J).
Рис. 11.45. Basidiomycetes. Прорастающие споры (1000х): А — Exidiopsls еffusa со вторичной спорой; В — Auricularia judae: 1 — 3 — базидиоспоры (2 — базидиоспора с поперечными перегородками, 3 — базидиоспора с конидиями); С — Pleurotus ostreatus с проростковой гифой
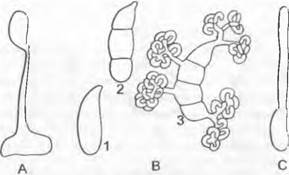
Между мейоспорами разных типов скрещивания возникают — часто уже тогда, когда они сидят на спороцистах, — копуляционные мостики (К), через которые плазма и ядра одной споры мигрируют в другую.
От начинающего после этого расти дикариотичного мицелия отчленяются дикариотичные конидии, которые активно отбрасываются (баллистоконидии). Как дикариотичная, так и гаплоидная стадия могут размножаться конидиями. У пшеницы Tilletia cañes вызывает твердую, или вонючую, головню, —Urocystis tritici — головню стеблей или полосатую головню листьев. Виды Eniyloma поражают преимущественно сложноцветные растения. С Tilletia caries можно успешно бороться путем кратковременного «протравливания» посевного материала высокой температурой или ядовитыми веществами либо путем опыления веществами, которые убивают приставшие к семенам головневые споры. На пораженном растении злака образуется несколько миллионов головневых спор, которые при молотьбе напыляются на посевной материал и после посева заражают молодые проростки (проростковая инфекция), поэтому болезнь может легко распространиться на многие растения; вследствие этого раньше погибало до 20 % (иногда даже 60 %) урожая зерна.
От рассматриваемого дальше порядка Ustilaginales порядок Tilletiales отличается, кроме строения базидии, также тем, что в головневых спорах происходит не только кариогамия, но и чаще всего уже мейоз.
Рис. 11.46. Ustomycetidae: А — Н — Ustilaginales: А, В — Ustilago scabiosae, проросшие головневые споры и образование мейоспор (s) на четырехклеточной базидии (110х); С — Ustilago carbo, копулирующие базидиоспоры (1 200х); D — Entyloma calendulae, мицелий с пряжками и дикарионами; Е — G — Ustilago vuijckii, образование головневых спор, дикариотичные и диплоидные головневые споры; Н — Ustilago sp., прорастающие в питательном растворе головневые спорвы (350х); J, К — Tilletiales, Tilletia caries: J — проросшая из головневой споры базидия с четырьмя парами расположенных на ее вершине мейоспор (300х); К — две копулировавшие базидиоспоры, прорастающие в дикариотичный мицелий, с конидией (к) (650х)
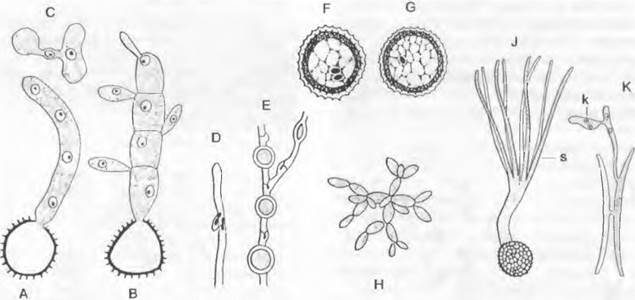
2. Порядок: Ustilaginales. Не имеющие плодовых тел устиляговые обитают как паразиты обычно в межклетниках высших растений и развивают в определенных органах своих хозяев (например, корнях, осях побегов, завязях плодов, тычинках) толстостенные споры. Грибы этого порядка, как и предыдущего, привлекают особое внимание как патогенные для растений организмы. Ustilago mayáis образует на соцветиях (и других частях) кукурузы желваки и пузыри размером с кулак, которые заполнены головневыми спорами. Другие виды рода Ustilago заполняют завязи и частично прилегающие структуры соцветий ячменя и пшеницы пылящим порошком головневых спор (пыльная головня — например, U. avenaeвызывает пыльную головню овса). В случае пыльной головни ячменя (U. hordei) и пшеницы (U. tritici) головневые споры образуются еще до раскрытия цветков в молодых завязях и распыляются, когда растения находятся в полном цвету. Споры переносятся ветром и прорастают в том же году между чешуями здоровых цветков (цветковая инфекция). Развивающийся из базидиоспор мицелий врастает затем в образующееся семя и зимует в его зародыше.
Жизненный цикл. Из базидиоспор, имеющих половой знак «+» или «-» (биполярный гетероталлизм), при благоприятных условиях вырастает дрожжевидный псевдомицелий; он гаплоиден и может жить только сапротрофно. Его можно также культивировать на искусственных питательных средах. Если генотипически различные клетки «+» и «-» псевдомицелиев встречаются между собой, то через копуляционный мостик происходит слияние плазматического содержимого (плазмогамия) и группировка ядер в дикарион. Содержимое одной клетки переходит в другую, поэтому воспринимающая клетка становится двухъядерной. Она вырастает в дикариотичную гифу, которая отныне способна заражать растение-хозяина. Способность к паразитизму ограничена, таким образом, дикариофазой. Дикариотичный мицелий распространяется в хозяине и образует там в определенных органах головневые споры, в которых осуществляется кариогамия. В деталях проходят следующие стадии: дикариотичный мицелий, который у ряда видов имеет пряжки (рис. 11.46, D, Е), проникает в проростке растения-хозяина по межклетникам до апикальной меристемы и растет сначала вместе с растением, не вызывая внешних симптомов болезни, пока не достигнет определенных мест, например, тычинок или завязей. Ткани хозяина полностью разрушаются, а в плотном сплетении мицелия образуются расположенные цепочками наподобие бус вздутия, которые окружаются толстой стенкой коричнево-черного цвета (рис. 11.46, Е) и отделяются от образовавшей их гифы. Они распыляются из места образования, как зола, поэтому их называют «головневые споры». В этой форме головневые споры гомологичны молодым базидиям, поскольку здесь также происходит кариогамия. Однако сами головневые споры еще не представляют собой спороцисты типа базидии, поэтому их называют пробазидии. Головневые споры прорастают (чаще всего только после перезимовки) в гифенный вырост, впоследствии поперечно септированный (рис. 11.46, А, В). При этом происходит мейоз, так что теперь в каждой из 4 отгороженных поперечной перегородкой клеток находится одно гаплоидное ядро. На этой стадии вырост — пробазидия — соответствует септированной фрагмобазидии. Эта пробазидия отшнуровывает вбок гаплоидные споры (так называемые споридии, В), причем возникающие в результате мейоза дочерние ядра мигрируют обратно в базидию. Они имеют генетически детерминированные половые знаки «+» или «-» в соотношении 1:1. При хорошем питании от базидии могут многократно отшнуровываться новые базидиоспоры. Цикл развития гаплодикариотичный: он аналогичен гаплодиплоидному циклу дрожжей.
Как и у дрожжей, некоторые формы (например, у Ustilago mayáis — возбудителя пузырчатой головни кукурузы) чисто гаплоидны. Встречаются также другие отклонения. Некоторые формы уже тетраполярны (см. рис. 11.55).
3. Порядок: Ехоbasidiales. Представители порядка паразитирует, как и Тaphrinales из аскомицетов, на цветковых растениях (в Европе прежде всего на вересковых) и не образуют плодовых тел. Часто они вызывают галлоподобные деформации на пораженных частях растения-хозяина (рис. 11.47, А), которые возникают из-за гипертрофии мезофилла (рис. 11.47, В). Мицелий пронизывает ткань растения, проходя по межклетникам и через клетки. Гифы паразита выходят на поверхность хозяина через устьица или между клетками эпидермиса и образуют там несептированные базидии (рис. 11.47, С). На тупых, сильно растопыренных стеригмах располагаются рядом друг с другом изогнутые споры, которые пассивно отпадают, образуют поперечные перегородки и прорастают конидями (рис. 11.47, С). В чистой культуре из спор вырастают скопления клеток, почкующихся наподобие дрожжей. На основании особенностей ультраструктуры и биохимии Ехobasidiale, относят, как делаем здесь и мы, к Ustomycetidае.
Рис. 11.47. Ustomycetidae, Exobasidiales, Exobasidium vaccinii: А — Vaccinium vitis-idaea с тремя листьями, пораженными Exobasidium (2/Зх); В — поперечный разрез через пораженный лист; слева нормальное образование листовой ткани, справа развитие ткани, гипертрофированной из-за поражения грибом (60х); С — прорывающийся между клетками эпидермиса мицелий с базидиями и прорастание базидиоспор (330х)
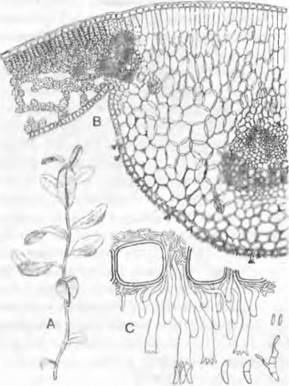
2. Подкласс: Уредомицетиды (Uredomycetidae)
К этому подклассу относятся паразиты, обычно еще не образующие плодовых тел. Характерная особенность — хорошо выраженная дикариофаза, воспроизводимая дикариоспорами (уредоспорами). Она предшествует образованию четырехклеточных фрагмобазидий с поперечными перегородками (см. рис. 11.49, D, F). Последние возникают всегда из округлых пробазидий; такие структуры часто отсутствуют в других подклассах базидиомицетов (например, среди Ustomycetidae y Tilletiles и Exobasidiales; среди Нуmenomycetidае — у подавляющего большинства). Этот подкласс, насчитывающий несколько тысяч видов, включает грибы-возбудители широко распространенного заболевания растений — ржавчины.
1. Порядок: Uredinales (ржавчинные). Ржавчинные грибы живут как паразиты, прежде всего в межклетниках, не убивая пораженные ткани. В клетки хозяина внедряются гаустории(рис. 11.48, А). Изредка мицелий обильно разрастается по всему растению, но чаще всего он распространяется лишь вблизи места заражения. На дикариотичных гифах пряжки отсутствуют. С аскомицетами ржавчинные сходны по следующим признакам: биполярный гетероталлизм, наличие спермациев и воспринимающих гиф в качестве половых органов, септы с простыми порами, хорошо представленные анаморфы. За редкими исключениями, ржавчинные грибы не образуют каких-либо заметных плодовых тел. Это можно считать приспособлением к паразитированию на чаще всего недолговечных вегетативных органах высших растений. Ржавчинные грибы характеризуются большим разнообразием типов спор (полный цикл развития охватывает 5 разных типов спор; см. рис. 11 50), которые закономерно сменяют друг друга и связаны со сменой ядерных фаз, а часто и со сменой хозяев.
Рис. 11.48. Uredomycetidae, Uredinales: А, В — Puccinia graminis: А — пикнида на барбарисе на продольном разрезе (140х), В — эцидий на барбарисе (140х); С — Gymnosporangiumclavariaeforme, пикнида на листе боярышника, прорывающая эпидермис на верхней стороне (450х); D — Peridermium strobi, отшнуровывание одноядерных спермациев (1 200х); Е — Phragmidium speciosum, базальные клетки (b) с копуляционным мостиком (1 200х); F — Phragmidium violaceum, зрелая эцидиоспора (800х); а1, а2 — двухъядерные эцидиоспоры; ак — цепочки дикариотичных эцидиоспор; е — эпидермис; eh — воспринимающие гифы; h — гаусторий; m — гаплоидный межклеточный мицелий; р — перифизы; рр — псевдоперидий; pz — палисадные клетки с гаусторием (h); s — спермации; z1, z2 — промежуточные клетки
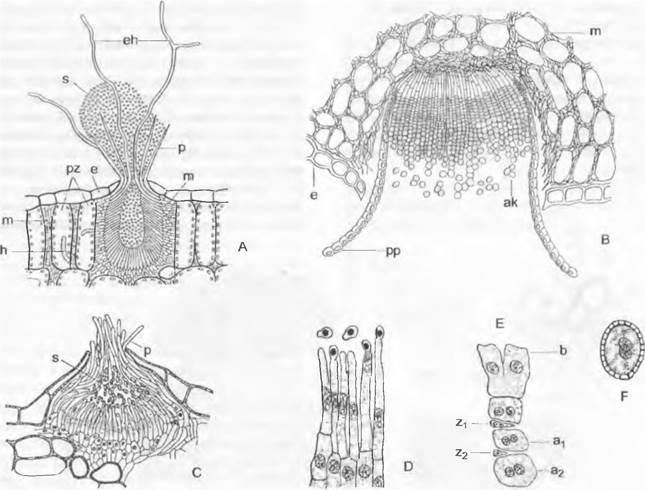
Дикариотичные гифы возникают из базальных клеток зачатка эцидия, когда они получат ядро одним из двух возможных путей. При переносе ядер спермациями важную роль играют вышеназванные пикниды. Их кувшиновидные плектенхиматические мицелиальные тела при созревании прорывают верхний эпидермис пораженных листьев барбариса и становятся видны как желтоватые пустулы (рис. 11.48, А). Они содержат, кроме стерильных гиф возле выводного отверстия (перифиз), короткие, плотно прижатые друг к другу гифы в центре, которые отчленяют мелкие одноядерные эллиптические спермации (так называемые пикноспоры, рис. 11.48, D). Последние, однако, прорастают на питательном растворе в короткую проростковую трубку, а при переносе на здоровый лист не могут заражать его. Их функция состоит в том, чтобы перенести свое ядро на так называемые воспринимающие гифы (рис. 11.48, А: еh). Это ответвления гаплоидного мицелия, которые выходят между клетками эпидермиса и перифизами и выдаются над поверхностью листа. Поперечных перегородок у них нет. Спермации сливаются только с воспринимающими гифами противоположных типов скрещивания (+ х -), что при смешанной (+, -) инфекции не составляет сложности. Кроме того, пикниды выделяют пахучую жидкость («нектар»), которую собирают насекомые, так что спермации могут быть перенесены ими и на другие листья, первоначально инфицированные только одним типом скрещивания.
Внедрившееся в воспринимающие гифы ядро мигрирует (в примыкающие к ним гифы) от клетки к клетке через перфорации в клеточной стенке до зачатка эцидия, где в базальных клетках возникает стадия дикариона. Второй путь состоит в том, что перенос ядра происходит при соматогамии: сливаются простые «+»- и «-»-гифы в ткани хозяина, если имела место смешанная инфекция. Базальные клетки зачатка эцидия, ставшие теперь дикариотичными, разрастаются в бокаловидные эцидии яркооранжевой окраски, которые прорываются на нижней стороне листа. В них возникают многочисленные цепочки, состоящие чаще всего из правильно чередующихся между собой дикариотичных эцидиоспор и мелких, позже расплывающихся и исчезающих промежуточных клеток (рис. 11.48, Е: z1/z2). У видов некоторых родов (например, Puccinia) самые конечные споры каждой цепочки, а также все споры периферических цепочек утрачивают репродуктивную функцию и склеиваются друг с другом в плотный покров (псевдоперидий, рис. 11.48, В: рр). За счет давления постоянно образующихся у основания цепочек новых спор (у Рuccinia graninis свыше 10 000 в одном эцидии) псевдоперидий и эпидермис разрываются, так что споры (сначала угловатые из-за давления со всех сторон, потом округлые) могут разноситься ветром.
Со сменой ядерной фазы (с гаплоидной на дикариотичную) изменяется также круг растений-хозяев. Эцидиоспоры прорастают только на хлебных и дикорастущих злаках. Их проростковая трубка внедряется через устьице в ткань этого второго хозяина и развивается в межклеточный (интериеллюлярный), локально ограниченный мицелий. Он содержит дикарионы, но не имеет пряжек. Вскоре мицетий переходит к интенсивному образованию дикариотичных конидий, которые здесь называются уредоспоры, или «летние споры» (рис. 11.49, Е). Последние возникают поодиночке из вздувающихся конечных клеток, несущих их гиф. Внешне это спороношение выглядит как мелкие черточки ржавой окраски (отсюда название — ржавчинные грибы!), прорывающие эпидермис. Они обеспечивают распространение гриба летом (перенос на другие экземпляры того же хозяина). Каждая отдельная уредопустула образует очень много (на восприимчивом хозяине — миллионы) уредоспор. Они сразу же инфицируют другие растения злаков, на которых уже через 3 недели после заражения развиваются новые уредопустулы. Таким путем болезнь очень быстро распространяется, причем и на большие расстояния.
К осени дикариотичный мицелий в уредопустулах или в других местах дает еще одну форму спороношения — двуклеточные телейтоспоры (рис. 11.49, А: t; С). В клетках этих спор пары ядер сливаются между собой (кариогамия). Телейтоспоры имеют толстые стенки, устойчивы к высыханию и холоду и служат для перезимовки в состоянии покоя. Следующей весной каждая из двух диплоидных клеток (= пробазидия) телейтоспоры претерпевает мейоз и прорастает через образуемую заранее проростковую пору (рис. 11.49, D: к) в вытянутую базидию (D, F). Между четырьмя гаплоидными ядрами образуются поперечные перегородки, и из каждой из этих четырех клеток вырастает базидиоспора (мейоспора), в которую переходит ядро (D). Базидиоспоры отбрасываются и переносятся ветром на первого хозяина — барбарис. Этим замыкается цикл развития, в ходе которого имеются монокариотичные гаплоидные ба- зидиоспоры (1) и гаплоидные спермации (2), дикариотичные эцидио- (3) и уредоспоры (4), а также сначала дикариотичные, а потом диплоидные телейтоспоры (5) — всего пять различных типов спор.
Рис. 11.49. Uredomycetidae, Uredinales: А — D — Puccinia graminis: А — группа уредоспор (u) с одной толстостенной двухклеточной телейтоспорой (t) между ними, В — прорастающая уредоспора (А, В — 300х), С — стебель хлебного злака с телейтоспороношением (150х); D — двухклеточная телейтоспора, прорастающая двумя базидиями (300х); Е — Phragmidium rubí, край уредоспороношения после разрыва отброшенного влево эпидермиса (е) растения-хозяина; споры разной степени зрелости (565х); F, G — телейтоспоры (500х): F — Uromyces appendiculatus, одноклеточная телейтоспора, проросшая базидией (ядра не показаны), G — Phragmidium vioiaceum; внизу с парными ядрами, которые в обеих верхних клетках уже слились; к — проростковые поры; р — парафиза
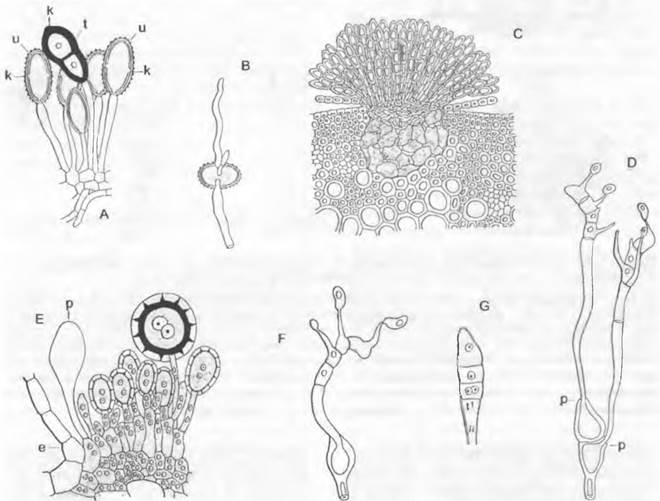
При так называемом Еu-типе цикл развития полный и имеется смена поколений. Стадия гаплоидного гаметофита подавлена, а дикариотич- ная спорофитная фаза преобладает; эта тенденция еще усиливается за счет повторного образования уредоспор (рис. 11.50), реже — эцидиев (виды Сronartium). Наряду с видами, имеющими облигатную смену хозяев (разнохозяинность) и регулярную последовательность гаплоидной и дикариотичной фаз, есть такие виды, жизненный цикл которых проходит на одном и том же хозяине (однохозяинность). За счет подавления той или иной формы спороношения жизненный цикл также упрощается; могут выпадать: эцидиоспоры (Brachy-тип, например, Uromycesfabae); эцидио- и уредоспоры при наличии периода покоя у телейтоспор (Micro-тип, например, Tranzschelia fusca) или без него (Lepto-тип, например, Puccinia malvacearum)',уредоспоры (Opsis- тип, например Gymnosporangium juniperinum); уредо- и телейтоспоры (Endo-тип, например, Endo- phyllum sempervivi); эцидио-, телейто- и базидиоспоры (несовершенные ржавчинные грибы). При этом всегда сохраняется выраженная дикариофаза. В местностях с коротким вегетационным периодом такие укороченные типы развития, ограниченные одним хозяином, имеют преимущество.
Рис. 11.50. Uredomycetidае, Uredinales. Схема развития Рuccinia graminis. Светлые линии: гаплоидная фаза; двойные темные линии: дикариотическая фаза; жирные темные линии: диплоидная фаза. Мелкие клетки — барбарис, крупные — злак в качестве растений- хозяев (ср. рис. 11.48, 11.49). Аес — эцидий, В — базальная клетка; Рус — пикнида; R! — редукционное деление; Sо — соматогамия; Sр — оплодотворение спермациями; Т — телейтоспороношение (число спор, как у U, уменьшено); U — уредоспороношение
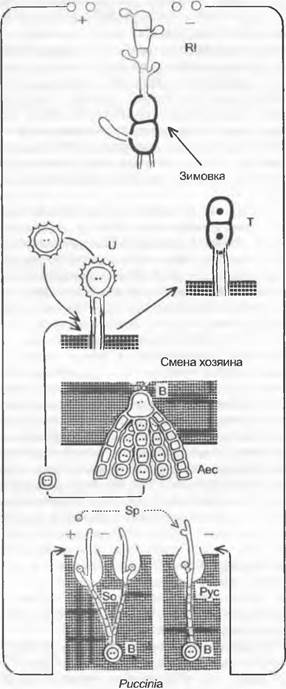
Так же, как и возбудитель стеблевой ржавчины злаков, возбудитель широко распространенной ржавчины гороха (Uromyces pisi) относится к Eu-типу. Пикниды и эцидии встречаются на видах молочая (Euphorbia cyparissias и др.). Пораженные растения остаются неветвящимися, имеют желтоватые, короткие, толстые листья и чаще всего не цветут. Уредо- и (здесь одноклеточные) телейтоспоры развиваются на листьях гороха (Pisum sativum) и видах Lathy rus. На одном хозяине проходит все развитие возбуди теля ржавчины фасоли (Uromyces phaseoli). Пикниды и эцидии встречаются летом на несколько гипертрофированных, горбатых листочках фасоли, на которых осенью образуются уредоспоры и, наконец, телейтоспоры. Так же ведут себя паразитирующие на листьях Rosaceae виды рода Phragmidium (см. рис. 11.49, Е). Эцидии здесь не окружены псевдоперидием, как у возбудителя стеблевой ржавчины злаков, а цепочки эцидио- спор прорывают, часто на обширной площади, эпидермис таким же образом, как и уредоспоры (тип Саеоmа). Некое подобие плодовых тел представляют собой слизистые скопления телейтоспор, приподнимающиеся вверх, как палочка, у видов Gymnosporangium, и образования из цепочек телейтоспор у видов Cronartium.
Филогения. Ржавчинные грибы представляют собой эволюционно очень древнюю группу и встречаются уже в карбоне как паразиты на папоротниках. Образующие базидии Uredinales и образующие сумки Taphrinomycetidae, возможно, произошли от одной предковой группы. Их общий предок должен был иметь самостоятельно питающуюся дикариофазу. И у тафриномицетид паразитирующие на папоротниках представители свидетельствуют о древности этой группы. В мезозое Uredinales перешли на голосеменные, особенно хвойные, и, наконец, с верхнего мела — на покрытосеменные. С проникновением в области более холодного климата у них, возможно, и возникли покоящиеся пробазидии, т. е. телейтоспоры, с толстыми стенками.
Ржавчинные грибы — опасные возбудители болезней растений. Особенно значительны вызываемые ими потери урожая зерновых (в отдельных случаях до 25 %, однако в целом они заметно не превышают примерно 5%). По всему земному шару распространен вид Puccinia graminis, вызывающий стеблевую ржавчину, которую из- за темной окраски, скопления телейтоспор называют также черной ржавчиной. Он поражает все наши виды хлебных злаков и многочисленные дикорастущие злаки.
В Средней Европе наносимый им ущерб не столь велик, как в более теплых странах, так как там развитие этого относительно теплолюбивого гриба протекает быстрее. В Германии особенно опасна желтая ржавчина (возбудитель — P. striiformis со скоплениями уредоспор от светло-желтого до оранжевого цвета), поражение которой может носить характер эпифитотий, особенно пшеницы, а также ржи, ячменя и различных дикорастущих злаков. Промежуточный хозяин гриба неизвестен.
P. coronata — возбудитель корончатой ржавчины овса и некоторых дикорастущих злаков — имеет промежуточным хозяином Rhamnus cathartica и еще некоторые виды, a P. hordei,возбудитель карликовой ржавчины ячменя, образует эцидии на видах Ornithogalum.
Другие виды Puccinia встречаются на спарже, моркови, луке, крыжовнике и других культурных растениях; виды Uromyces — на горохе, фасоли и сахарной свекле; Gymnosporangium — на листьях груши («сетчатая ржавчина»). К другим семействам ржавчинных грибов относятся Melampsora Uni — однохозяинный возбудитель ржавчины льна, который разрушает его лубяные волокна; имеющие значение для лесного хозяйства грибы Melampsorella caryophylla- сеагum («ведьмины метлы» и рак пихты белой, уредо- и телейтоспоры на гвоздичных); Cronartium ribicola (его эцидиальная стадия поражает веймутову сосну и часто вызывает ее гибель; эцидии прорываются как крупные пузыревидные скопления из коры дерева; другой хозяин в жизненном цикле — смородина).
Надежда на то, что с разнохозяинными ржавчинными грибами можно бороться путем устранения промежуточных хозяев, может оправдываться лишь очень ограниченно, поскольку у подавляющего большинства видов могут зимовать и уредоспоры или уже осенью заражаются молодые всходы озимых злаков, а также различные кормовые и декоративные злаки (в случае желтой ржавчины — также пырей); кроме того, уредоспоры могут заноситься из чрезвычайно отдаленных регионов (в Германию — даже через Альпы). Поскольку применение химических средств борьбы небезопасно, пытаются выводить устойчивые к ржавчине сорта культурных растений. Это имеет свои трудности, так как у каждого вида ржавчинных грибов насчитывается большое число физиологических рас, морфологически чаще всего не различимых, которые специализируются относительно различных сортов культурных растений, а за счет мутаций и рекомбинаций постоянно возникают все новые расы. Подобное расообразование у грибов- возбудителей болезней растений играет в фитопатологии очень большую роль, поэтому работа селекционеров в области выведения устойчивых сортов никогда не закончится.
Сюда примыкает порядок Septobasidiales, представители которого живут совместно со щитовками (симбиоз?).
На основании анализа ДНК и особенностей ультраструктуры скорее сюда, а не к головневым грибам, следует отнести грибы-возбудители головни пыльников гвоздичных. Виды рода Microbotrium (Microbotryales)заполняют пыльники хозяина черно-фиолетовыми головневыми спорами вместо пыльцы. У женских растений раздельнополых смолевок (Silene alba и S. dioica) при инфекции таким грибом стимулируется образование пыльников, в которых затем развиваются споры гриба.
3. Подкласс: Hymenomycetidae
У представителей этого подкласса мицелий чаще всего образует плодовые тела — макроскопические структуры из гиф, на которых базидии располагаются палисадным слоем (гимением). Ход развития совпадает с тем, который имеет место у пластинчатых грибов: на гифах возле септ часто образуются пряжки (рис. 11.51).
В жизненном цикле,который сходен у всех образующих плодовые тела базидиомицетов (рис. 11.51), базидиоспоры прорастают в гаплоидные гифы, состоящие из одноядерных клеток. После слияния гиф (соматогамии) тотчас же образуется дикариотичный мицелий, часто несущий пряжки (см. рис. 4.6) Слияние гиф, партнеров скрещивания, контролируется генетически.
Биполярная и тетраполярная несовместимость. У многих базидиомицетов пол определяется не одним геном (пара аллелей «+» и «-»), а двумя несцепленными генами. Аллели одного фактора обозначаются А1 и А2, другого — В1 и В2. Диплоидное ядро зиготы содержит, таким образом, А1/А2 и (!) В1/В2. После редукционного деления одно плодовое тело содержит базидиоспоры 4 типов: А1В1, А2В2, А1В2 или А2В1. Только скрещивание с различными А- и В-факторами может продолжить жизненный цикл, дав начало дикариотичному мицелию (часто с пряжками у септ гиф); так, сочетание А1В1 х А2В2, совместимо, но А1В1 х А1В2 или А1В1 х А1В2, напротив, несовместимы (рис. 11.55).
Здесь говорят о тетраполярной гомогенной несовместимости. С ней связан тот факт, что при встрече гаплоидных монокариотичных гиф, которые прорастают из спор одного плодового тела, фактически только в 25 % случаев возникает мицелий с пряжками. Генетической рекомбинации мицелиев разного географического происхождения способствует явление множественных аллелей, так как в ней могут участвовать и другие аллели, например, А3В3 и А4В4. Поэтому скрещивания с факторами А1/А2 или В1/В2 х А3/А4 или В3/В4(например, А1В1х А3В3) на 100% совместимы. Механизм биполярной или тетраполярной несовместимости, который преимущественно связан с наличием множественных аллелей, способствует таким путем перекрестному оплодотворению (аутбридингу), причем этот эффект при тетраполярной несовместимости сильнее (25%: 100%), чем при биполярной (50 % : 100 %). Если у аскомицетов встречается только биполярная несовместимость, то у базидиомицетов отмечается все больше случаев тетраполярного механизма.
Пряжки гомологичны крючку аскомицетов. Они принимают в себя временно одно из двух дочерних ядер, возникших при делении клетки, с тем, чтобы оно потом опять перешло в ту из дочерних клеток, которая расположена дальше от конца гифы (рис. 11.51, 5 — 7).
Рис. 11.51. Hymenomycetidae: А — развитие пряжкового мицелия: 1 — генотипически различные (+, -) споры, 2 — прорастание спор в мицелий без пряжек, 3 — 4 — копуляция, 5 — 7 — образование первой пряжки, 8 — образование следующей пряжки, 9 — пряжковый мицелий с дикариотичной материнской клеткой базидии (а), молодой базидией с диплоидным ядром (b) и зрелой базидией (с) со спорами разного типа скрещивания (с); В - схема развития шляпочного гриба. Светлыми линиями показана гаплоидная фаза; тонкими темными — дикариотичная фаза; жирными темными — диплоидная фаза. Пряжки не изображены, базидии изображены непропорционально крупными по отношению к шляпке
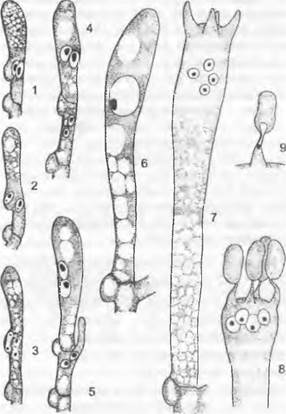
Процесс образования пряжки повторяется при каждом клеточном делении, так что возникает обильно разветвленный «пряжковый мицелий», содержащий в каждой клетке дикарионы и имеющий пряжки у каждой поперечной перегородки (рис. 11.51, 8, 9). Устанавливающаяся таким образом дикариотичная фаза (в отличие от аскомицетов) способна к самостоятельному существованию. Такой мицелий может годами разрастаться в почве, древесине и других субстратах и претерпевать бесчисленные дальнейшие клеточные деления с конъюгированным делением ядер дикариона, пока под влиянием еще не известных нам условий не разовьются плодовые тела. В противоположность аскомицетам (см. рис. 11.32) плодовое тело базидиомицетов состоит исключительно из дикариотичных гиф (рис. 11.51). Его возникновение, таким образом, не связано, как у аскомицетов, каждый раз с новым актом плазмогамии при половом процессе. Последний у базидиомицетов происходит только один раз для возникновения многолетнего дикариотичного мицелия, способного, в отличие от аскомицетов, из года в год давать плодовые тела. На поверхности или внутри плодовых тел (чаще всего на их нижней стороне) булавовидно вздутые конечные клетки дикариотичных гиф располагаются в виде палисадного слоя — гимения (рис. 11.51, В). Только в этих конечных клетках — молодых базидиях — два ядра дикариона сливаются между собой (кариогамия, рис. 11.52,6), после чего сразу же происходит мейоз (с определением типа скрещивания) и образуются четыре гаплоидные мейоспоры — базидиоспоры. Развивающиеся из базидиоспор гаплоидные мицелии соответствуют гаметофитам; возникающий из них в результате соматогамной копуляции дикариотичный мицелий может рассматриваться как дикариотичный спорофит.
Рис. 11.52. Нуmеnоmycetidае, Аgаricales. Образование пряжек и развитие базидии: 1 — 7 — Оudemansiella mucida (620х): 1 — начало образования пряжки в двухъядерной конечной клетке, 2 — клеточное ядро отходит в пряжку, 3 — одновременное ядерное деление, 4 — образование стенки внутри пряжки и рядом с ней, материнская клетка базидии отделяется от несущей ее клетки (клетки-ножки), 5 — слияние пряжки с клеткой-ножкой, 6 — два гаплоидных ядра материнской клетки базидии сливаются в диплоидное ядро, 7 — молодая базидия с четырьмя ядрами будущих базидиоспор, образовавшимися в результате мейоза (вверху четыре зачатка стеригм); 8, 9— Рsathyrella (1 500х): 8 — базидия с 4 ядрами перед их переходом в молодые базидиоспоры, расположенные на вершине базидии, 9 — переход клеточного ядра через стеригму в базидиоспору
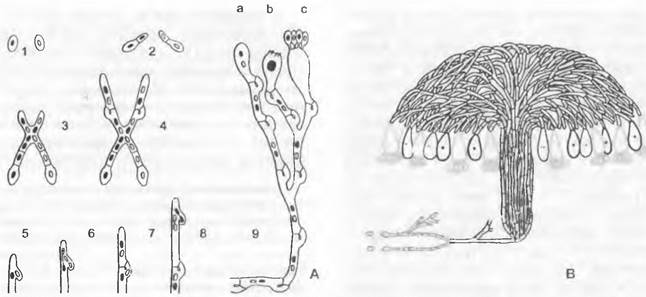
При развитии базидиоспор концы стеригм вздуваются, образуя споровые мешочки (рис. 11.52, 8). Из 4 гаплоидных ядер каждое протискивается через свою стеригму (рис. 11.52, 9), и теперь в каждом споровом мешочке образуется одна спора. Однако почти всегда стенка споры рано сливается со стенкой мешочка, так что двойная природа споровой оболочки становится незаметной; стенка мешочка образует при этом периспорий. Таким образом, мейоспоры только внешне кажутся образующимися экзогенно. Они преимущественно эллипсоидные, и уплощены, с одной стороны. Споры отбрасываются лишь на короткое расстояние за счет того, что находящаяся в состоянии сильного тургора базидия выдавливает из вершины стеригмы каплю, которая увлекает с собой спору (рис. 11.53, 11.54).
Рис. 11.53. Нуmеnоmусеtidае: А, В — Аuricularia auricula-judае: А — плодовое тело (натуральная величина), В — продольный разрез через гимений (400х); С, D — Тremella mesenterica:С — плодовое тело (натуральная величина), D — поперечный разрез через гимений (400х)
Рис. 11.54. Hymenomycetidae: А — L — Dacrymycetales: Calocera cornea (900х), отбрасывание базидиоспоры. А, В — вытягивание стеригмы. С — F — отшнуровывание базидиоспоры (продолжительность примерно 40 мин), G, Н — образование капельки жидкости (t) у места прикрепления споры (длительность примерно 10с), I — отбрасывание споры вместе с капелькой (t), J, К — опадание стеригмы; L — Schizophyllales, Schizophyllum commune (1 500x), место прикрепления базидиоспоры к стеригме; bw — стенка базидии; sw — стенка споры
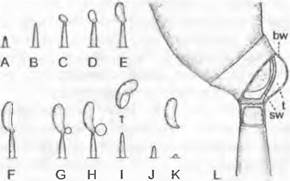
К рассматриваемой группе относится большинство организмов, известных в обиходе как грибы (см. рис. 11.57; 11.59; 11.61). Их мицелий часто можно культивировать, однако в культуре плодовые тела образуются редко. Многие виды часто имеют быстрорастущие плодовые тела, особенно в конце лета и осенью. Можно различать гимениальные и гастроидные плодовые тела.
У гименнальных плодовых телгименомицетов (рис. 11.55 — 11.57) в более узком смысле слова гимен ий в ходе развития становится открытым, а базидиоспоры активно отбрасываются от базидий. Гимений содержит базидии и в отдельных случаях цистины, расположенные в виде палисадного слоя (рис. 11.56, 11.59, С). Гименофор представляет собой макроскопическую структуру, увеличивающую спороносную поверхность. Морфология гимениапьных плодовых тел весьма разнообразна — от корковидных или булавовидных до сильно разветвленных, консолевидных или дифференцированных на шляпку и ножку («шляпочные грибы»). Столь же разнообразно и строение гименофора: он может быть гладким или складчатым, иметь вид пчелиных сот, пор, трубочек, шипов или пластинок. Существуют почти все возможные комбинации форм плодовых тел и типов гименофора, причем в ходе эволюции достигалось максимально возможное число спор при наиболее благоприятном пути их распространения. Плодовое тело гимениального типа развивается различным образом. У гимнокарпного плодового тела гимений (гименофор) с самого начала закладывается на открытых внешних поверхностях. В случае гемиангиокарпного развития (рис. 11.59, А, В) гимений образуется сначала внутри еще молодых плодовых тел. Однако при вытягивании ножки и раскрытии шляпки исходная обвертка разрывается. Ее остатки часто сохраняются и на зрелом плодовом теле как части общего и/или частного покрывала. Частное покрывало (velum partiale — vp) образует в таком случае кольцо (т) или еще недолгое время соединяет шляпку и ножку в виде паутинистой вуали (cortina) либо же полностью исчезает. Общее покрывало (velum universale — vu) оставляет обвертки в виде влагалища у основания ножки (volva — v; например, у бледной поганки) и/или белые лоскутья и клочья на поверхносп и шляпки (например, у Amanita muscaria — мухомора).
Рис. 11.55. Нуmenomycetidае, Рleurotus ostreatus. Обеспечение перекрестного оплодотворения путем гомогенной несовместимости (тетраполярный механизм, множественные аллели). Плодовые тела с несколько различающимися по окраске шляпками — штаммы одного и того же вида разного географического происхождения. А. В — факторы скрещивания используемых для скрещивания монокариотичных мицелиев; • — скрещивание дает дикариотичный пряжковый мицелий
Рис. 11.56. Hymenomycetidae. Формы цистид (Poriales, 750х): А — Tubulicium; В — Vararía; С — Peniophora; D — Tubulicrinis; Е — Stereum, F — Hyphoderma
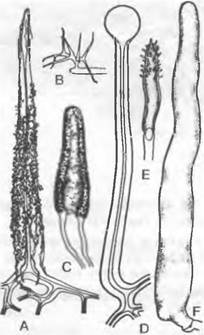
Рис. 11.57. Hymenomycetidae. Различные гимениальные плодовые тела: А — Ramaria botrytis (1/2х); В — Stereum hirsutum (12х); С — Dendropolyporus umbellatus (1/Зх); D — Hydnum repandum (1/2х); Е — Phellinus igniarius, многолетнее плодовое тело с годичными зонами (1/2х)
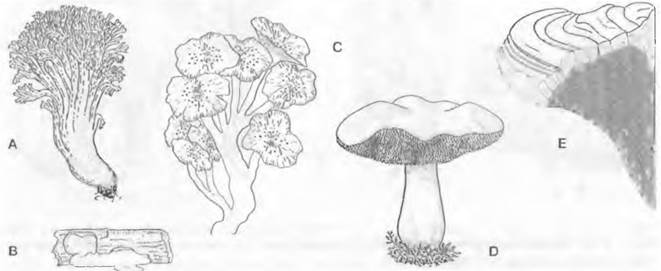
Псевдоангиокарпное развитие первоначально сходно с гимнокарпным, однако край шляпки сильно завертывается внутрь, так что его гифы сплетаются с гифами коры ножки.
Отбрасывание спор. Споры отбрасываются от базидий на небольшое расстояние. Оно составляет у грибов с трубчатым гименофором примерно половину диаметра трубочки (у грибов с пластинчатым гименофором — половину расстояния между пластинками). Полет быстро превращается в падение по вертикали за пределы трубочки или пространства между пластинками, где споры подхватываются движением воздуха. Собственно, распространение спор осуществляется воздушными потоками. Если положить зрелую шляпку пластинчатого гриба пластинками вниз на лист бумаги, то уже через несколько часов выпадающие споры образуют на нем четкий отпечаток пластинок. Зрелое плодовое тело шампиньона полевого (Agaricus campestris) диаметром 10 см имеет поверхность гимения 1 200 см2, которая в совокупности производит около 1,2 млрд спор; за один час отбрасывается приблизительно 40 млн спор.
В гимении наряду со зрелыми находятся и молодые базидии, а также стерильные гифы с дегенерировавшими дикарионами и более крупные, также стерильные конечные гифы разной формы — цистиды (см. рис. 11.56). Последние выполняют функцию защиты и выделения (например, у Poriales) или, возможно, препятствуют слипанию пластинок (например, у Coprinus). Они важны для систематического подразделения и определения видов. Плектенхима плодового тела — трама (рис. 11.58) — также дифференцируется: например, толстостенные «скелетные гифы» служат для прочности; толстостенные разветвленные «связующие гифы» схватывают другие гифы; тонкостенные «генеративные гифы» образуют базидии.
Рис. 11.58. Hymenomycetidae, Poriales, Tra- metes versicolor. Разрез через край растущего плодового тела с двумя зачатками трубочек гименофора. Толстостенные: скелетные гифы; тонкостенные: гифы, образующие базидии; полностью черные: связующие гифы (150х)
Рис. 11.59. Hymenomycetidae, Agaricales. Схематические продольные разрезы через плодовые тела с покрывалами: А — частное покрывало (velum partiale, vp); В — общее покрывало (velum universale, vu) и частное покрывало, слева — у молодого, справа — у зрелого плодового тела; С — разрез через гимений Нypholoma (1 000х); b — базидия; bо — молодая базидия; с — цистида; f — остатки общего покрывала на шляпке; m — кольцо, остаток частного покрывала; s — базидиоспора; st — стеригма; sh — субгимений; t — трама; v— обвертка (volva), остаток общего покрывала у основания ножки
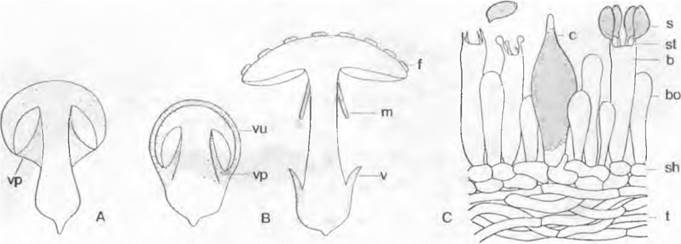
Рис. 11.60. Нуmenomycetidae, Boletales. Вариегатовая кислота (производное пульвиновой кислоты) из Suillus variegatus: вверху — неокисленная (желтая) форма, внизу — окисленный (синий) анион; R = ОН
Гастроидные плодовые тела «гастеромицетов» (рис. 11.61) образуют базидии внутри себя; первоначально закладывающийся на ранних стадиях развития гимений разрушается ко времени созревания спор или вообще не образуется. Развитие плодовых тел ангиокарпное или гемиангиокарпное. Плодовые тела могут быть закрытыми, подразделенными или неподразделенными внутри на отдельные камеры, или же споровая масса выносится вверх на способном к растяжению элементе (рецептакуле) из оболочки (перидия) плодового тела (рис. 11.61, А — С). Споры не отбрасываются от базидий. Распространение спор осуществляется ветром, а в отдельных случаях также насекомыми и млекопитающими.
Рис. 11.61. Hymenomycetidae, «Gasteromycetes»: А — С — Phallales: А — Phallus impudicus, зрелое плодовое тело с каплями оплывающей глебы на шляпке и молодое плодовое тело на продольном срезе (1/2х), В — Anthurus archeri (1/2х), С — Clathrus ruber (1/2х), D — Boletales (ранее относили к Sclerodermatales), Scleroderma citrinum, на срезе видна глеба с белыми бесплодными участками в виде прожилок (1/2х); Е, F — Nidulariales: Е — Cyathus striatus (натуральная величина), F — Sphaerobolus stellatus, справа — отбрасывание эндоперидия (Зх); G — Geastrales, Geastrum quadrifidum (1/2х)
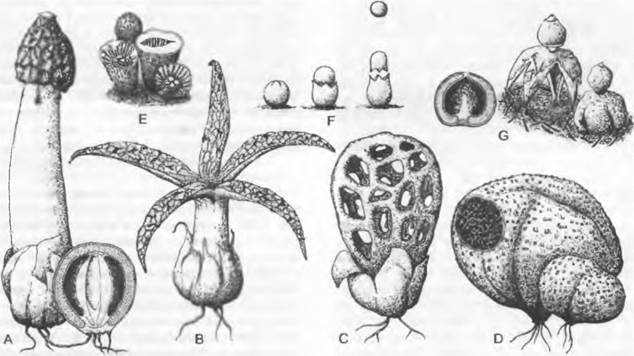
Между гимениальными и гастроидными плодовыми телами существуют переходы, так что большинство «гастеромицетов» имеет тесную родственную связь с типичными гименомицетами, поэтому сейчас его помешают в общий подкласс — Нуmеnomycetidae.
Сначала выстраиваются преимущественно порядки, представители которых имеют базидии с поперечными или продольными перегородками (фрагмобазидии). Базидиоспоры прорастают конидиями, вторичными спорами или дрожжевидными клетками (см. рис. 11.45). Вторичные споры — однократно отшнуровывающиеся структуры, в которые переходит единственное ядро базидиоспоры.
Морфологическая взаимосвязь между Uгеdomycetidae (Uredinales, Septobasidiales) и следующих за ними порядков (Tremellales, Auricula- riales) становится очевидной благодаря наличию переходных форм. Так, представители некоторых семейств Uredinales (например, Chrysomyxa с эпи- диями на Picea и уредо- и телейтоспорами на Ericales) вместо толстостенных телейтоспор развивают базидии типа Aitricularia. Uredinella coccidiophaga обитает аналогично Septobasidiales на щитовках, однако имеет пробазидии, сходные с телейтоспорами. Patouillardina (Tremellales) с беспорядочно септированными базидиями является промежуточным звеном между Auriculariales и Tremellales. У Bourdotia (Tremellales) продольные перегородки в базидиях не доходят до их основания; такие формы представляют собой переход от фрагмобазидии к холобазидии. Если у Uredinales базидии лишь в отдельных случаях образуются на своего рода примитивных плодовых телах, то Auriculariales и Tremellales, за исключением немногих форм без плодовых тел, показывают усложняющийся ряд разнообразнейших типов плодовых тел: плоские налеты, булавовидные, состоящие из ножки и головки, консолевидные, состоящие из ножки и шляпки. Сходные формы плодовых тел, возникавшие в ходе параллельного эволюционного развития, будут нам встречаться неоднократно и среди других групп базидиомицетов (см. рис. 11.57). Поверхность гимения может увеличиваться за счет образования складок, структур типа пчелиных сот или зубчиков. В пределах одного вида Auricularia auricula-judae можно наблюдать переходы от гладкой спороносной поверхности через складчатую к поверхности, имеющей облик сот или почти трубочек (= тенденция к выработке гименофора). У Auriculariales (частично) и Tremellales (закономерно) впервые в пределах упомянутых эволюционных рядов встречается пряжковый мицелий; он возникает после соматогамии и отличает дикариофазу.
1. Порядок: Tremellales, дрожалковые. Фрагмобазидии этих грибов разделены продольными перегородками, расположенными крест-накрест (см. рис. 11.53, D; 11.44, В). Они обитают преимущественно на отмершей древесине, изредка на других субстратах или же паразитируют на других грибах. У наиболее примитивных представителей плодовые тела отсутствуют. Виды рода Тremella образуют студенистые плодовые тела от мозговидной до листовидной формы, имеющие желтую, буроватую или черную окраску (см. рис. 11.53, С). Студенистая «ткань» плодовых тел служит для накапливания влаги. Шляпковидное плодовое телоимеющее боковуюnножку, несет на нижней стороне шипы, которые покрыты гимением. Exidiopsis — ср. с рис. 11.45.
2. Порядок: Dacrymycetales. Эти грибы отличаются 2-споровой холобазидией в форме камертона (Dacrymyces, см. рис. 11.44, D), чаще всего септированными базидиоспорами, прорастающими конидиями, и тем, что они обитают как сапротрофы на древесине. Плодовые тела имеют вид простых корочек или пустул либо представляют собой образования от состоящих из ножки и головки, бокаловидных до разветвленно-булавовидных. Таким образом, здесь мы имеем набор вариантов, конвергентно сходных с имеющимися в других эволюционных линиях. Плодовые тела, окрашенные каротиноидами, различны — чаще всего от мягких до вязко-студенистых. Порядок показывает определенные общие черты с уже обсуждавшимися паразитическими Еxobasidiales. В частности, это касается прорастания базидиоспор, имеющих поперечные перегородки, крючковидно изогнутыми конидиями — эта особенность не встречается у прочих базидиомицетов, имеющих холобазидии. С другой стороны, здесь имеется также родство с Тulasnellales (С).
3. Порядок: Auriculariales (уховидные грибы). С представителями Uredomycetidae виды этого порядка сближает наличие фрагмобазидии с поперечными перегородками. Существование родов, виды которых являются паразитами без плодовых тел (например, Неrpobasidium, Неlicobasidium), указывает на возможную родственную связь с Uredinales. Иудино ухо (Аuricularia auricula-judae, см. рис. 11.53, А) — вид со студенистыми темнокоричневыми плодовыми телами в форме ушной раковины, вырастающими из стволов бузины, может рассматриваться как эволюционно продвинутый. На своей вогнутой поверхности, которая может варьировать от гладкой до складчатой, они несут гимений. Базидии, как у всех Auriculariales. разделены поперечными перегородками на расположенные друг над другом 4 клетки, от каждой из которых вырастает вбок стеригма с мейоспорой (см. рис. 11.53, В; 11.44, А). У некоторых Auriculariales базидия имеет в основании шаровидное вздутие. В этой «пробазидии», которая некоторое время представляет собой конец дикариотичной гифы, происходит слияние ядер. После мейоза из нее вырастает собственно базидия. Базидиоспоры имеют ко времени прорастания (например, у Аuricularia) несколько поперечных перегородок и образуют от каждой возникшей таким образом клетки множество конилий. К этому порядку примыкают Тulasnellales (см. рис. 11.44, С).
Последующие порядки базидиомицетов, имеющие исключительно холобазидии, раньше объединялись в группу Нolobasidimycetidae. Споры всегда прорастают гифами. Парентосомы у долипоровых септ пронизаны порами и образуют, таким образом, ситовидную структуру. Форма и размер холобазидий разнообразны (см. рис. 11.44). Наряду с широко распространенной булавовидной формой (F) встречаются, например, сильновыпуклые (Е), возникающие сбоку на несущей гифе плевробазидии (Н), многократно прорастающие друг из друга репетобазидии (J). Имеется большое разнообразие форм плодовых тел и характера поверхности слоя, несущего гимений (гименофора). Как и у рассмотренных ранее порядков (в том числе характеризующихся наличием фрагмобазидии), здесь также в разных эволюционных линиях происходит выработка сходных типов плодовых тел, так что у этих грибов, как и у водорослей, можно видеть разные типы дифференциации. Внешняя форма плодовых тел здесь часто служит критерием для различения родов и семейств того или иного порядка.
По внешней форме плодовых тел раньше различали искусственные (т. е. не отражающие родственных связей) группы афиллофоровых (Роriales = Аphyllophorales в широком смысле), агариковых, или пластинчатых (Аgaricales в широком смысле), и гастеромицетов (Gasteromycetales). В настоящее время при систематизации этих организмов пытаются отражать родственные связи, а именно исходить из сходства как можно большего числа признаков разного рода: например, таких микроскопических структур, как строение гимения и плектенхимы плодового тела (= трамы), состава метаболитов, ДНК и т.д. При группировке таксонов более низкого ранга в таксоны более высокого ранга (например, порядки) важную роль играют переходные формы, которые, с одной стороны, имеют признаки разных таксонов, а с другой — показывают переходы признаков. Из-за наличия плавных переходов между вышеназванными формами пло
довых тел афиллофоровых, агариковых и гастеромицетов системы, построенные на особенностях плодовых тел, оказались несостоятельными.
4. Порядок: Cantharellales. Наиболее известный представитель порядка — лисичка настоящая (Cantharellus cibarius; Cantharellaceae), ценный съедобный гриб. Его состоящее из ножки и шляпки плодовое тело имеет гименофор из толстых, часто анастомозирующих между собой складок, и гимений с удлиненными стихически- ми базидиями, т. е. в них оси веретен ядерного деления располагаются по продольной оси базидии. В качестве пигментов имеются и каротиноиды, которые вообще редки у высших холобазидиомииетов. Некоторые виды имеют почти гладкий гименофор, как, в частности, виды рода Clavulina, имеющие плодовые тела в форме оленьих рогов с булавовидными расширениями на концах. Ежовик желтый (Hydnum repandum. см. рис. 11.57, D) отличается состоящими из шляпки и ножки плодовыми телами с шиповидным гименофором.
5. Группа порядков- Gomphanae. Выделение этой группы порядков основано на данных анализа ДНК. Морфологически они весьма гетерогенны: с одной стороны, сюда относятся виды с плодовыми телами от булавовидных до имеющих форму оленьих рогов (Gomphales с родами (iomphus, Romana, см. рис. 11.57, А), ас другой стороны — также виды с гастроидными плодовыми телами (Geastrales, Pliallales, см. рис. 11.61, А — С, G).
5.1. Geastrales. Плодовые тела земляных звездочек (Geastrum, см. рис. 11.61, G) имеют характерный облик: экзоперидий разрывается на лопасти, которые отгибаются вниз и освобождают бумагообразный эндоперидий с заключенной в нем глебой (G). Рифы имеют возле септ пряжки (в отличие от Lycoperdales — см. ниже). Базидии выпуклые; на них на коротких стеригмах образуются споры (часто их больше 4).
5.2. Phallales. Плодовые тела на ранних стадиях развития окружены слизистой оболочкой, которая позже разрывается и становится сходной с вольвой (velum universale) некоторых пластинчатых шляпочных грибов. Первоначально заключенная внутри оболочки глеба (см. поря док 10.4) подразделяется на камеры и при созревании образует слизистую массу с сильным неприятным запахом, содержащую базидиоспоры. Эта масса у многих представителей порядка выносится вверх способным к растяжению осевым элементом (рецептакулом). Распространение спор осуществляется насекомыми, которые привлекаются запахом Глебы, а у некоторых видов еще и яркой окраской рецептакула. Броские формы (так называемые «грибы-цветы») распространены особенно в тропиках. Обитающая в умеренной зоне веселка (Phatíus impudicus, см. рис. 11.61, А) внешне несколько напоминает сморчок, относящийся к аскомицетам, но имеет совершенно другой тип развития и структуру (конвергентное сходство). Молодое плодовое тело, окруженное мягкой белой оболочкой, в Западной Европе называют «ведьмино яйцо». Воль- ва состоит из внешнего и кожистого внутреннего перидия, и студенистого промежуточного слоя. Развитие плодового тела можно наблюдать на извлеченном из земли снаружи мягком, а внутри твердом «ведьмином яйце»: рецептакул — уже заложившийся внутри молодого плодового тела— вытягивается в течение нескольких часов до длины примерно в 15 см, разрывая при этом оболочку, которая остается в виде бокала, вынося наверх «шляпку». Последняя закладывается уже в «ведьмином яйце» в виде окружающего ножку слоя, имеющего форму колокола; снаружи она примыкает к вольве. «Шляпка» состоит из кожистого несущего слоя, разделенного на камеры, и расположенной на нем чернозеленоватой слизистой споровой массы, имеющей неприятный запах. Эта «шляпка» согласно одной точке зрения в целом соответствует Глебе, согласно другой — она отчасти представляет собой вырост рецептакула (несущий слой), а отчасти соответствует Глебе (споровая масса). Споровая масса расплывается и капает с ячеистой поверхности «шляпки». Мухи (синие и зеленые падальные) распространяют споры на поверхности своего тела. У видов тропического рода Diclyophora от вершины ножки отходит ажурная сеточка, которая сначала находится между «шляпкой» и ножкой, а потом конусообразно спускается вниз (почему немцы называют этот гриб «дама с вуалью»).
Рещеточник (Clathrus, см. рис. 11.61, С) и цветохвостник (Anthurus, В) имеют сходный тип развития плодовых тел с тем только различием, что у первого рода окрашенный в красный цвет рецептакул имеет форму сетки, а у второго на вершине расходится несколькими лопастями.
На основании строения и развития плодовых тел и отсутствия гимения оба рассмотренных выше порядка могут рассматриваться как весьма изолированные среди Hymenomycetidae.
6. Порядок: Hymenochaetales. Гифы окрашены коричневыми пигментами (стирилпиронами), локализованными в мембранах. Пряжки у септ гиф отсутствуют, а в гимении обращают на себя внимание чаще всего заостренные, толстостенные, окрашенные в коричневый цвет цистиды (= щетинки). Споры гладкостенные.
Подразделение на роды отражает различия в формах плодовых тел. Гладкие корочки, которые частично в виде ребрышек отстают от субстрата, представлены в роде Hymenochaete.Многолетние консолевидные плодовые тела со слоистым трубчатым гименофором отличают род Phellinus. По чередующимся слоям трубочек и трамы на поперечном срезе можно судить о возрасте плодового тела (см. рис. 11.57, Е). Трубочки с узкими выходными огверстиями в виде пор показывают у таких плодовых тел (как и в других порядках) положительное геотропическое направление роста с высочайшей точностью, что обеспечивает в подобном случае выход спор. Coltricia имеет жесткое, хорошо сохраняющееся плодовое тело с трубчатым гименофором, состоящее из шляпки и ножки.
7. Порядок: Russulales. Грибы, известные как сыроежки и млечники, однозначно отличаются часто амилоидной (см. объяснение при описании Leotianae) орнаментацией спор, а также чаще всего группами округлых клеток между вытянутыми гифами и органами выделения, проводящими терпеноиды (млечные гифы, синеющие в ванилин-Н2SO4 цистиды). Сыроежки (Russula) с их хрупкими, легко разламывающимися пластинками имеют в качестве пигментов азотсодержащие, растворимые в воде руссуптеридины. Плодовые тела млечников (laclarías) выделяют при повреждении млечный сок, иногда несколько окрашенный. В пределах порядка наблюдаются плавные переходы от форм с плодовыми телами, дифференцированными на шляпку и ножку, с развитым гимением к формам с подземными гастроидными плодовыми телами. Сюда же примыкают растущие на гнилой древесине виды Stereum (Stereaceae). У них многослойные распростертые плодовые тела частично отстают от субстрата, а гименофор гладкий (см рис. 11.57, В). Сходное изменение трактовки родственных связей необходимо принять также для Heterobasidion (см. Portales) и Albatrellus (овечьего трутовика).
8. Группа порядков: Aphyllophoranae. Представители группы ооразуют трубчатый гименофор.
8.1. Polyporales. Жесткомясистые, состоящие из шляпки и ножки плодовые тела видов рода Polyporus, давшего название порядку, могут обрастать своим исключительно трубчатым гименофором посторонние предметы (например, ветки, соломины злаков). В культурах были получены плодовые тела также из монокариотичного мицелия, в данном случае гаплоидного. У Dendropolyporus (см. рис. 11.57, В) возникает несколько шляпок на общей сильноразветвленной ножке.
8.2. Portales. В настоящее время эта группа пока еще выделяется в порядок. Большинство видов способно тем или иным образом обрастать своими плодовыми телами посторонние предметы. Гимений располагается свободно на плодовом теле, он образуется рано, по гимно- карпному типу и по мере увеличения плодового
тела дает все новый и новый прирост. Виды типового рода Poria (Роnaceae) образуют плоские, прижатые к субстрату корки, которые несут на верхней стороне трубчатый гименофор. У Corticium (Corticiaceae) плодовые тела также прижатые, с гладким гименофором без особого пространственного членения. Часто встречающиеся на пнях деревьев виды рода Trámeles (например, Т. versicolor) имеют отходящие от субстрата кожисто-корковые плодовые тела с трубчатым гименофором. Такой же гименофор образуют консолевидные многолетние плодовые тела настоящего трутовика (Femes fomentarius). который в Германии особенно часто встречается как паразит на буке. У березового трутовика (березовой губки, Pipío poms belulinus) внутренняя часть однолетнего плодового тела относительно мягкая. У других трутовиков гименофор переходного вида — от лабиринтовидного (Daedaleaquercina, дубовая губка) до сходного с пластинчатым (Daedaleopsis tricolor). Различные плодовые тела и типы гименофора образуются также в пределах рода Gloephyllum\ плодовые тела заборного гриба (С. sepiarium) растут в виде отстающих от субстрата корок и имеют гименофор типа пластинчатого, тогда как пахучий трутовик (С. odoratum) с запахом, напоминающим запах фенхеля, с нижней стороны консолевидного плодового тела имеет трубчатый гименофор. Оранжево-бурая окраска плодовых тел обоих видов обусловлена траметином. Вид Heterobasidion annosus — корневая губка — вызывает гниль (особая форма белой гнили, см. бокс 11.4, с. 124) и по признакам микроструктуры и характеристикам ДНК должен быть отнесен скорее к Russulales.
9. Порядок: Thelephorales. Представители порядка отличаются от Poriales и Hymenochaetales пигментированными спорами с бугорчатой поверхностью, имеющими чаще всего попарно расположенные бородавки или шипы. Эти грибы накапливают телефоровую кислоту в форме пигментированных отложений на гифах трамы. В пределах порядка существуют переходы от плоских, прилегающих к субстрату плодовых тел с гладким или шиповатым гименофором (Tomentella) к булавовидно-разветвленным или шляпковидным плодовым телам (Thelephora). Плодовые тела, состоящие из ножки и шляпки с шиповатым гименофором, отличают, в частности, род Sarcodon (например, сарксщон черепитчатый, или ежовик пестрый, S. imbricatus), а трубчатый гименофор — род Boletopsis. Ежовик желтый (Hydnum repandum. Cantharellales), имея внешне сходное плодовое тело с шиповатым гименофором, однако отличающийся совершенно другими микроскопическими и химическими признаками, не может быть отнесен к этому порядку (конвергенция!).
10. Группа порядков: Agaricanae (включая Lycoperdanae). Представители этой группы — прежде всего грибы, имеющие недолговечные плодовые тела с четким разделением на ножку и шляпку, не способные обрастать посторонние предметы и образующие с нижней стороны пластинчатый гименофор. Четкие родственные связи таких типичных пластинчатых грибов с родами, отличающимися другой формой плодовых тел, в частности афиллофороидного или гастероидного типа, привели к новой концепции порядков в данном случае.
10.1. Порядок: Schizophyllales. Щелелистник (Schizophyllum) — объект, часто используемый в экспериментальных исследованиях (см. рис. 11.54, L); гименофор здесь в форме жестких, продольно расщепленных пластинок, деформирующихся при изменении влажности. Плодовые тела могут рассматриваться как комплексные, составленные из простых бокаловидных плодовых тел (какие сохранились еще у Суphellopsis).
10.2. Порядок: Agaricales. Как и Роriales, этот порядок выделен временно. Его окончательный состав (в Средней Европе около 2000 видов) определяется соответствием набора признаков типовому роду Agaricus (шампиньон). У многих представителей этой группы гимений образуется внутри плодовых тел в схизогенных полостях, но при их раскрытии он становится свободным. Однако наряду с гемиангиокарпным развитием встречаются также псевдоангиокарпное и гим- нокарпное развитие. Плодовые тела недолговечные. У представителей некоторых родов (например, навозник, или чернильный гриб, Сорrinus) они разрушаются уже через несколько часов после возникновения. Заложение гименофора осуществляется не постепенно и не от центра к периферии (как у Aphyllophoranae), а единовременно. Гименофор чаще всего состоит из радиально расходящихся вертикальных пластин, у зрелых плодовых тел выстилающих нижнюю сторону шляпки, расположенной на ножке. Основное сплетение гиф пластинок — трама — несет на внешней стороне слой, состоящий из округлых клеток — субгимений, который покрывается гимением. Гимений состоит главным образом из базидий разной степени зрелости и часто также из цистид (см. рис. 11.59, С). Веретено деления ядра располагается перпендикулярно продольной оси базидий (хиастобазидии).
К этому порядку относятся как съедобные грибы, подлежащие охране, так и опасные ядовитые грибы. В широком масштабе культивируется шампиньон двуспоровый (Аgaricusbisporus). Две (не 4!) споры на его базидиях содержат по два совместимых («+» и «-») ядра; таким образом здесь в отличие от большинства других Аgaricales не требуется половой процесс для начала нового жизненного никла, и при культивировании этот гриб можно размножать посредством дикариотичного посевного материала. Опасные и коварные ядовитые грибы (симптомы отравления появляются только по прошествии от 6 до 24 часов после употребления в пищу!) — различные виды рода Amonita, особенно бледная и весенняя поганки (соответственно A. phalloïdes и A. virosa). Они содержат ядовитые циклические пептиды (ама- и фаллотоксины). Их плодовые тела имеют свободно закругленные, т. е. не приросшие к ножке (в отличие от шампиньона) пластинки, остающиеся белыми или беловатыми, а также мешковидную обвертку у основания ножки — остаток общего покрывала. Пластинчатые грибы, имеющие определенное сходство с Polyporales (см. 8.1), такие, как пилолистник (Lentinus) и вешенка (Pleuralus), в настоящее время опять относят к Agaricales. P. eryngii при определенных условиях культивирования образует вместо пластинчатого гименофора неправильные трубочки.
К Agaricales относятся также роды Kuehneromyces, Psilocybe, Armillariu. Omphalina, Clitocybe. Lepista, Strobilurus и Termitomyces.
10.3. Порядок: Boletales. К порядку относятся не только грибы с трубчатым гименофором типа белого гриба (Boletus edulis), но и определенные грибы с пластинчатым гименофором, с замкнутыми и корковидными плодовыми телами. Это связано с наличием переходных форм или следующих признаков: пигментированные, часто веретеновидные споры и / или дивергирующие (расходящиеся) возле гимения гифы трамы гименофора, пигменты типа производных пульвиновой кислоты (например, вариегатовая кислота). Этими пигментами обусловлено при наличии оксидаз часто наблюдаемое синее окрашивание плодового тела (см. рис. 11.60). Эволюция в пределах порядка, как предполагается, шла от обитателей древесины (бурая гниль, см. бокс 11.4, с. 124) с прилегающими в виде корок к субстрату плодовыми телами с гладким гименофором (Coniophora) либо уже гименофором в виде складок или сот: серпула плачущая, или настоящий домовый гриб (Serpula lacrymans), образует быстро растущие, мягкомясистые плодовые тела размером до 1 м2. На древесине обитают также представители порядка с плодовыми телами — от имеющих форму раковины до дифференцированных на ножку и шляпку, образующих пластинчатый гименофор, как Tapinella atrotomentosa. Следующий эволюционный уровень — чаще всего образующие микоризу шляпочные грибы с гименофором в виде пластинок, трубочек или промежуточного типа. Paxilhts involutus образует микоризу с разными деревьями, Р. filamentosas — только с ольхой (см. 9.2.3; бокс 11.4). У Phylloporus пластинчатый гименофор из-за наличия многочисленных анастомозов между пластинками подразделен на отдельные камеры. Cюmphidius (мокруха), несмотря на ярко выраженный пластинчатый гименофор, имеет такие же микроскопические и химические признаки, как и представители порядка с трубчатым гименофором. Грибы этого порядка с трубчатым гименофором отличаются от имеющих сходный гименофор представителей других таксонов (например, Роlyporus из Рolyporales, Boletopsis из Тhelephorales, Соltricia из Нуmenochaetales) рядом особенностей. К отличиям относятся: недолговечность плодовых тел, не способных обрастать посторонние предметы; отделяемые от остальной части шляпки трубочки; другие микроскопические признаки и пигменты. Как приспособление к вулканическим почвам, сухим периодам года и т. д. можно рассматривать появление эволюционно наиболее молодых гастроидных форм, которые также сплошь образуют микоризу, а часть из них очень специализирована в отношении физиологии питания. Виды рода Gastroboletus, растущие в Америке, можно расположить в ряд, где ножка становится короче, трубчатый гименофор претерпевает все большую деградацию, плодовое тело замыкается за счет соединения края шляпки с ножкой и начинает развиваться под землей. Подобные ряды можно проследить в родах Тruncocolumella и Rhizopogon с исключительно подземными плодовыми телами.
Далее следуют порядки, объединяющие типичные формы с замкнутыми плодовыми телами (гастеромицеты, однако в настоящее время без Geastrales и Рhallales). Это грибы, у которых плодовые тела, по крайней мере в молодости, замкнутые, гастроидные, ангиокарпные, их оболочка (перидий) вскрывается или распадается только после созревания спор (см. рис. 11.61, Е, F).
10.4. Порядок: Lycoperdales. Представители порядка известны под названием дождевики. Гастроидные плодовые тела, имеющие форму от шаровидной до булавовидной, только на ранних стадиях развития находятся под землей. Снаружи они имеют оболочку, чаще всего двухслойную, состоящую из экзо- и эндоперидия. У зрелых плодовых тел экзоперидийразрывается; на жестком кожистом эндоперидии от него остаются зернышки, бородавочки или комочки. Эндоперидий часто открывается порой на вершине. Внутренняя часть плодового тела сначала подразделена на камеры, поверхность которых выстлана гимением. Базидии короткие, булавовидные. Они несут на сильно вытянутых стеригмах шаровидные споры, которые активно не отбрасываются, а освобождаются за счет разрушения базидий. Зрелое плодовое тело содержит только порошковидную массу — глебу,которая состоит из бесчисленных спор и волокон. Она возникает в результате разрушения гимения и стерильной ткани, так что остаются и составляют глебу только споры и волокна ткани (капиллиций) — остатки длинных гиф трамы. У представителей некоторых родов нижняя часть плодового тела стерильна (субглеба). В лесах часто встречается дождевик шиповатый (Lycoperdon perlatum), из плодовых тел которого при надавливании высвобождается облачко порошащей бурой споровой массы. У гигантского дождевика (Langermanniagigantea), плодовое тело которого может достигать 50 см в диаметре, Глеба содержит до 7,5 млрд спор. Если бы каждая спора развивалась в плодовое тело, то уже в 21-м поколении потомков образовалась бы такая грибная масса, которая в 800 раз превышала бы массу Земли.
Виды рода порховка (Bovista) растут преимущественно на лугах и пастбищах; их плодовые тела, как, например, у порховки гигантской, не имеют стерильной субглебы. Правда, ложнодождевик (Scleroderma; см. рис. 11.61, D) относится к Boletales (пигменты, ДНК!).
10.5. Порядок: Nidulariales. В плодовом теле заключены отдельные фрагменты глебы, которые могут распространяться как целые единицы — перидиоли. Внутри перидиолей базидии расположены в виде гимения. У Cyathus (бокальчика) перидиоли при созревании лежат как маленькие диски в бокаловидном перидии (см. рис. 11.61, Е). Сфероболюс (SphaeroboIus) образует одну-единственную перидиоль, которая из-за мгновенного выворачивания внутреннего слоя экзоперидия отбрасывается на расстояние до 1 м (см. рис. 11.61, F).
Родственные связи последних двух порядков с относящимися к Agaricales родами Сорrinus и Agaricus (разделение порядков 10.4 и 10.5 с 10.2 выражено, таким образом, слабо!), а также с растущей на дубовой древесине печеночницей (Fistulina) вырисовываются достаточно ясно.
Применение. С древности до середины XIX столетия трут, изготовляемый из ткани плодового тела Fames fomentarius, служил для получения огня (откуда название гриба — настоящий трутовик). Так, в поясном мешке найденного в леднике трупа неолитического человека из долины Эца («Эци») был найден этот гриб, так же, как и укрепленные на полосках кожи куски плодовых тел березового трутовика (Piptoporus betulinus), возможно, для применения как лекарственного средства. Многочисленные виды гименомицетов собирают для использования в пищу, некоторые из них, например, шампиньон (Agaricus bisporus), также культивируются. Ежегодный сбор культурного шампиньона во всем мире 670 000 т. Наряду с шампиньоном для питания выращиваются, особенно в Восточной Азии, различные другие базидиомицеты (например, сиитаке, Lentinus edades). Пытаются «одомашнивать» и ранее не возделывавшиеся съедобные грибы. Однако некоторые очень ценные виды (например, белый гриб, лисичка обыкновенная) не дают в культуре плодовых тел. При введении грибов в культуру интересуются не только получением плодовых тел как приправы и пищи для человека и животных, но и использованием их для переработки различных отходов; помета животных, соломы, опилок и других материалов, содержащих лигнин и целлюлозу (рециклизации). Путем заражения древесины мицелием соответствующих грибов (например, летнего опенка, Kuehneromyces mutabilis, или вешенки устричной, Pleurotus ostreatus) изменяют ее свойства; она становится легкой и как «микодревесина» используется для промышленной обработки (например, при изготовлении карандашей вместо древесины кедра). Виды Psilocybe (Agaricales) содержат галлюциногенные индольные производные, которые используются также при религиозных ритуалах, например, в Мексике. Различные грибы выделяют в культуральную среду антибиотики, которые используются отчасти в фармацевтических целях (например, пенициллин), отчасти как фунгициды (стробилурин из Strobilurus).
Некоторые грибы ядовиты. Об опасных поганках уже упоминалось; в целом известно примерно 150 видов ядовитых гименомицетов (аскомицетов — лишь около 10 видов), но среди них сильноядовитых немного. В старых плодовых телах съедобных грибов также образуются ядовитые вещества, как при порче мяса.
Бокс 11.4. Встречаемость и образования грибов (включая целлюлозные грибы)
Грибы, насчитывающие около 100000 видов (Oomycota 500, Eumycota свыше 90 000), обитают повсеместно как гетеротрофы, и в противоположность водорослям прежде всего на суше. Обитатели водной среды (менее 2 % всех видов) — это либо первичноводные организмы из числа размножающихся зооспорами Oomycetes и Chytridiomycetes, либо вторичноводные организмы — Ascomycètes и Fungi imperfecti (Moniliales). Водные грибы живут преимущественно в пресной воде, однако в последнее время выявлено некоторое число морских видов (особенно аскомицетов. но также и несколько базидиомицетов). Часто на их спорах имеются приспособления для парения в воде; например, у Moniliales конидии нитевидные или с 3 — 4 лучами.
Ископаемые грибы сохранились весьма незначительно. Древнейшие находки — это хитридиомицеты во фрагментах панцирей морских животных; они восходят к кембрию. В девоне встречены лишенные поперечных перегородок гифы в остатках наземных растений; в карбоне на папоротниках уже имелись ржавчинные грибы (и вероятно, также аскомицеты), а на корнях деревьев — микоризы; экземпляры хорошо сохранившегося пряжкового мицелия свидетельствуют о том, что уже в каменноугольном лесу имелись высшие базидиомицеты (гименомицеты?).
Грибы питаются как сапротрофы, биотрофные или некротрофные паразиты (последние вызывают быстрое отмирание живых организмов; по образу жизни близки к сапротрофам на отмерших остатках) или как компоненты симбиотических ассоциаций (например, микоризы лишайников). Нередко грибами заселяются особые субстраты, например, насекомые, мхи, другие грибы и т. д. (часто лишь определенные виды).
Поскольку грибы могут разлагать самые разные органические субстраты и поражать самые разные живые организмы, причиняя им вред или убивая их, они наносят значительный ущерб. Гриб Amorphotheca resinae (Eurotiales) освоил даже масла, бензин и смолы, питаясь содержащимися в них углеводородами; в авиации он наносит ущерб, вызывая закупорку системы питания двигателя и коррозию алюминия. Вызывая разрушение древесины, болезни человека, животных и растений, а также порчу продуктов питания и текстиля, грибы оказывают огромное отрицательное воздействие в хозяйственной деятельности. Вместе с тем велика и положительная роль грибов. Об этом говорилось выше, при рассмотрении отдельных классов грибов (спиртовое брожение — см. 6.10.2.1; антибиотики — см. Eurotiales; употребление в пищу— см.: применение базидиомицетов; стимуляция роста растений при микоризном симбиозе - см. 9.2.3). Из огромного разнообразия приспособлений к особым условиям жизни нам надо будет выбрать лишь немногие, экологически наиболее важные.
Грибы как разрушители древесины
В природе древесина отмирающих деревьев и пни разлагаются преимущественно грибами. Разложение древесины животными и бактериями, напротив, имеет лишь очень незначительный масштаб. Согласно современным исследованиям, бактерии, по-видимому, не бывают агрессивными разрушителями древесины. Из грибов в разложении древесины участвуют прежде всего базидиомицеты с гимениальными плодовыми телами (гименомицеты, например, Poriales, Hymenochaetales, Polyporales, Agaricales), отчасти аскомицеты (например, Ceratocystis) и несовершенные грибы. Некоторые из этих дереворазрушителей поражают как паразиты древесину еще живых растений, как Phellinus pini и Heterobasidionarmosum — возбудители гнилей сосны и ели. а также Phellinus alni на яблонях и других лиственных деревьях или настоящий трутовик (Pomes fomentarius) на буках и березах. Многие грибы живут сапротрофно только на отмершей древесине (например, виды Coriolus, Trámeles и GloeophyUym). После паразитирования возможны сапротрофный образ жизни на отмершей древесине, как у опенка осеннего (Armillaria mellen). который обитает на отмерших пнях деревьев и образует на них свои плодовые тела, но оттуда может переходить на живые деревья, прежде всего когда они (например, после длительной засухи) физиологически ослаблены («паразиты ослабленных хозяев»).
Видообразование шло по пути специализации на определенных хозяевах (например, Phellinus hartigii на пихте, Ph. robustus на дубе, Ph. hippophaecoki на облепихе), особенно среди родов, представленных преимущественно паразитами. Некоторые грибы — опасные разрушители древесины на складах и в постройках: например, домовые грибы Coniophoraputeana и особенно Serpula lacrymans могут наносить серьезный ущерб постройкам, начиная с сырых мест. Разрушение древесины грибами происходит в виде деструктивной (бурой)или коррозионной (белой, особая форма: красная) гнили. В первом случае гриб разлагает преимущественно целлюлозу, так что лигниновый компонент древесины сохраняется; древесина становится бурой, с поперечными трещинами и распадается на прямоугольные фрагменты (например, Coniophora, Serpula). Возбудители коррозионной гнили (например, Phellinus igniarius на иве) разлагают лигнин и целлюлозу, причем чаще всего они в противоположность грибам бурой гнили выделяют в субстрат фенолоксидазы; гнилая древесина вследствие процессов обесцвечивания становится белой и продольно-волокнистой. Отдельные древесные породы противостоят разложению с помощью ядов, которые прежде всего содержатся в ядровой древесине (токсины ядровой древесины). Некоторые грибы, однако, все-таки способны разрушать защищенную таким путем древесину, причем в обезвреживании токсинов, возможно, участвуют фенолокеидазы. Только в недавнее время стали обращать больше внимания на Moderfàule как на третий тип разложения древесины. Этот вид разложения вызывают микуромицеты с мелкими плодовыми телами (например, аскомицетами) или грибы без полового спороношения (Fungi imperfecti). По характеру разложения болезнь представляет собой медленно развивающуюся бурую (реже белую) гниль. Примером могут служить различные виды Chaetomium (Ascomycètes), гифы которых поражают вторичную стенку трахеид или же сосудов древесины. При названных типах гнилей снижается, иногда очень значительно, сопротивляемость древесины сжатию и изгибу. Однако синева древесины сосны не влияет на ее статические качества, так как ее возбудители (виды Ceratocystis) разрушают только содержимое клеток паренхимы древесины. В процессе приобретения серой окраски строительной древесиной. например в горах, по-видимому, уча ствуют несовершенные грибы, если этот процесс обусловлен не исключительно погодными условиями. Некоторые дереворазрушающие грибы вызывают ночное свечение (биолюминесценцию), как, например, опенок осенний (Armillaria melleà); у поселяющегося на старых оливковых деревьях гриба Omphalotus olearius светятся даже плодовые тела. Физиологически разложению древесины родственно разложение растительного опала (листьев) на лесной почве, в котором наряду с бактериями участвуют грибы (грибы лесной подстилки), и тем самым они вносят существенный вклад в образование гумуса.
Грибы как симбионты
Значительная часть высших растений образует с грибами симбиотическую ассоциацию — микоризу (см. 9.2.3). Различают облигатно и факультативно микотрофные растения. В Европе эктотрофная микориза наиболее характерна для хвойных пород и Fágales из покрытосеменных. Микориза (чаще всего эндотрофная) широко распространена и у культурных растений, таких, как земляника, томат, горох и хлебные злаки. У орхидных в природных условиях без микоризного гриба не могут прорастать мельчайшие семена, лишенные запасных питательных веществ Микориза отсутствует лишь у немногих высших растений, например, Cyperales, Plumbaginales, Brassicaceae. Из грибов не образуют микоризы оомицеты и хитридиомицеты, в остальных классах настоящих грибов они распространены, особенно среди аскомицетов и базидиомицетов (к которым относится большинство шляпочных грибов, обитающих на лесной почве), а также и среди зигомицетов (Endogone).
Часто грибы образуют плодовые тела только в ассоциации с определенным микоризным партнером или могут жить вообще исключительно в симбиозе с ним. Так, среди «обабков» (род Leccinum) подберезовик (L. scabrum) и подосиновик желто-бурый (L. testaceoscabrum) связаны с березой, L. vulpinum — с двуиглыми соснами, дубовый обабок (L. quercinum) — с дубом, подосиновик красный (L. aurantiacum) — с осинами, L. сагpini — с грабом, лещиной или осиной. Микоризные симбиозы могут «вырождаться» или переходить во взаимоотношения хозяин — паразит. Тогда или некоторые грибы паразитируют на растении-хозяине, или высшее растение, исходно служившее как партнер гриба, становится его паразитом (например, Neottia, см. 9.2.3). Исследования микориз находят практический выход при выращивании лесов на ранее безлесных территориях. Они обеспечивают теоретические обоснования внесения посевного материала грибов — микоризных партнеров. О симбиозе грибов с водорослями, который привел к стабильному партнерству в лишайниках, говорится в разделе, посвященном этим комплексным организмам (см.: Лишайники, 11.2; 9.2.4).
Исключительно многообразны симбиозы грибов с животными. Узкоспециализированные в отношении питания животные, такие, как кровососущие, питающиеся древесиной или растительными соками насекомые, содержат в определенных частях своего пищеварительного тракта или в особых органах, так называемых мицетомах, симбионты — бактерии и дрожжи, для переноса которых от одного поколения к другому существуют различные приспособления. Грибы-симбионты отчасти дают своим партнерам питание (древесина для личинок рогохвоста большого), отчасти гриб поедается насекомым, так что оно обходным путем — через гриб — может использовать в пищу древесину (например, жуки-точилыцики). Во всех этих случаях симбионты обеспечивают своим хозяевам выживание таким образом, что они по-разному дополняют обмен веществ партнера. Тропические муравьи-листорезы культивируют в своих подземных постройках мицелий определенных видов грибов (относящихся к Agaricales): утолщенные, богатые питательными веществами концы грибных гиф идут им в пищу; гриб, разлагающий целлюлозу, вносится муравьями на субстрат, образованный из разжеванных кусочков листьев, и при закладке нового гнезда переносится в него. У термитов также тщательно культивируемый грибной мицелий (Termitomyces, Agaricales) служит для питания только царицы и личинок. Сходное значение имеют так называемые грибы-амброзии, которые обитают в ходах короедов (Ipidae) и используются в пищу их личинками.
Грибы как возбудители болезней
Из 162 важных инфекционных заболеваний возделываемых в Средней Европе культурных растений 83 % вызываются грибами. Потери ежегодно бывают огромными и уже приводили к случаям массового голода (см. Phytophthora). Среди организмов, которые являются паразитами на растениях, наряду с животными, бактериями и вирусами грибы играют доминирующую роль. Некоторые группы (среди слизевиков — плазмодиофоромицеты, среди других, например, Peronosporales, Erysiphales, Uredinales, Ustilaginales) существуют почти исключительно как паразиты и возбудители болезней высших растений. Наряду с этим к паразитам растений относятся также многочисленные несовершенные грибы (см. 11.2. Deuteromycetes). Развитие паразитического гриба на растении-хозяине начинается с заражения. Гриб проникает через порезы, устьица или непосредственно через наружную стенку эпидермиса. Внедрение через кутикулу эпидермиса происходит при участии специальных экзоферментов (кутиназ) или механическим путем, когда кутикула прободается острым выростом инфекционной гифы. Обычно заражаются восприимчивые органы хозяина, например, корневые волоски, органы проростков, лепестки или рыльца пестиков. Для успешного заражения часто бывает достаточно одной споры. Ведущее значение для прорастания споры наряду с температурой имеет достаточная влажность, поэтому в сырые годы наблюдается особенно сильное развитие грибных болезней.
На внедрение гриба растение-хозяин реагирует защитными реакциями механической или химической природы — утолщением клеточных стенок, образованием защитных веществ, например, дубильных (которые обезвреживаются фенолоксидазами) или специфических фитоалексинов. Произойдет ли наконец заражение хозяина, зависит от вирулентности гриба и устойчивости хозяина. Устойчивость хозяина обусловлена его генетической конституцией, а также различными механическими, химическими и физиологическими факторами резистентности. С другой стороны, предрасположенность хозяина к заражению в соответствующее время зависит от внешних условий и может иметь решающее значение для заражения. Хорошее азотное питание приводит, например, к меньшей механической прочности клеток при быстром росте, из-за чего чаще всего повышается предрасположенность к заражению.
Вирулентность гриба показывает в пределах вида большие различия, обусловленные генетически. Среди 53 аскоспоровых культур даже из спор одного плодового тела у возбудителя мучнистой росы ячменя было получено 14 различных патотипов. У многих паразитных грибов виды разделяются на большое число форм, специфичных относительно определенных хозяев.
В качестве источников инфекции выступают все конидии и мейоспоры. Перезимовка осуществляется в виде стром, которые весной образуют плодовые тела (например, Venturia, Сlaviceps, Rhyviceps), в виде прорастающих весной зигот (Реronosporales) и телейтоспор (Uredinales); грибы могут зимовать в корневищах, клубнях, зимующих почках растений-хозяев и т. д. Распространение возбудителя болезни осуществляется (часто очень быстро) такими же путями, как распространение плодов и семян высших растений: чаще всего потоками воздуха, нередко животными и в последнее время также человеком, который уже перенес многие болезни растений с одной части земного шара в другую.
Эпифитопии грибных болезней мы знаем только в монокультурах отдельных растений, которые в ряде регионов даже были полностью уничтожены. Например, виноградарство на Тенерифе и Мадейре полностью прекратилось около 1850 г. из-за поражения виноградной лозы возбудителем мучнистой росы Uncinula necator.
При борьбе с болезнями растений (в противоположность бактериальным и грибным («грибковым») болезням человека) терапевтические меры играют лишь второстепенную роль. Важнейшими мерами здесь являются профи лактикческие: препятствование контакту возбудителя и хозяина (соответствующие агротехнические мероприятия, севооборот, удаление промежуточных хозяев ржавчинных грибов) либо уничтожение патогенного гриба до или во время его прорастания (протравливание посевного материала, опрыскивание или опыление фунгицидами). Важную роль играет также выведение сортов, устойчивых к возбудителям болезней (селекция на устойчивость). Предпосылкой успешной борьбы является точное знание цикла развития и условий жизни паразита.
У человека и животных грибы вызывают самые различные заболевания. Косвенно с образом жизни грибов связаны микотоксикозы и микогенные аллергии. Такие аллергии могутбыть вызваны спорами грибов, попадающими в дыхательные пути из воздуха. Содержание спор в воздухе часто чрезвычайно велико: так, водной сельскохозяйственной постройке было однажды обнаружено до 21 млн спор Aspergillus на 1 см3 воздуха. Вне помещений, напротив, эти показатели значительно ниже, а именно порядка 0,25 — 7 спор на 1 см3 воздуха. В легких только что забитых коров было насчитано до 1 700 колоний грибов на I г сырой массы, и именно тех видов, споры которых, имеющие величину менее 10мкм, могут перемещаться по дыхательным путям. Aspergillusflavus растет на различных орехах и образует афлатоксины, вызывающие поражение печени. В противоположность таким микотоксикозам, которые могут вызываться также, например, Claviceps, при пищевом отравлении грибами говорят о мицегизме (см. Agaricales).
Грибы, вызывающие микозы, могут рассматриваться как паразиты теплокровных животных и человека. По картине болезни микозы разделяются на поверхностные (поражаются кожа, волосы, ногти, перья, когти и копыта) и глубокие (развиваются внутри тела и могут приводить к смерти). Примерно 30 — 50 возбудителей микозов представляют собой дрожжевидные (Candida, Torulopsis, Cryptococcus) и мипелиальные (Aspergillus, Trichophyton, Мисог) грибы. Некоторые виды поражают исключительно теплокровных хозяев, другие живут в почве, откуда и происходит заражение. «Ножной гриб» (Trichophyton rubrum) заражает человека поросшими его мицелием мельчайшими кожными чешуйками, рассеянными по почве. Глубокие микозы практически не передаются при контакте. Заражение ими происходит через желудочно-кишечный тракт и особенно через органы дыхания. Виды Histoplasma — возбудители широко распространенного в теплых странах заболевания легких.
Хищные грибы
Хищные грибы (рис. 11.62) составляют особую группу факультативных паразитов. Эти грибы улавливают мелких почвенных животных (нематод, коловраток) или подвижные водоросли (Euglena) с помощью различных приспособлений, однако часто они могут культивироваться в отсутствие своих жертв на обычных питательных средах. К Chytridialesотносятся Polyphagus euglenae (рис. 11.62, А; см. также 11.2. Chytridiomycetes) и виды рола Amaudovia. Последние живут на поверхности воды (нейстон) и улавливают одноклеточные организмы с помощью шести длинных гиф с выростами в виде тонких волосков (рис. 11.62, В). Zoopagales (Zygomycetes) — все «охотники за животными»: они живут ловлей амеб и нематод. На гифах Zoopage thamnospira (рис. 11.62, G) удерживаются нематоды, в которые потом врастают служащие для питания гриба гаустории. Хищные грибы группы Arthrobotiysотносятся к несовершенным грибам (Hyphomycetes). Они ловят своих жертв (нематод) посредством клейких гиф или сетей (рис. 11.62, С), клейких «пуговиц» (Dactylella) и несжимающихся и сжимающихся колец (Dactylium). У Oomycetes также возникло хищничество, а именно в роде Zoophagus. Гифы образуют здесь придатки, покрытые слизистым выделением, на которых зависают коловратки (рис. 11.72, Е, F). Среди базидиомицетов хищничество наблюдалось у некоторых видов Pleuro- tus. В целом описано около 80 видов грибов, улавливающих животных.
Рис. 11.62. Хищные грибы: А — Polyphagus, Р. euglenae с 10 сжавшимися эвгленами на разных стадиях их переваривания (200х); В — Arnaudovia, А. hyponeustica с пойманной особью Tylenchus (150х); С, D — Arthrobotrys, А. Oligospora: С — с ловчими петлями, D — с пойманной нематодой (150х); Е, F — Zoophagus: Е — Z. insidians с тремя пойманными коловратками (90х), F — коловратка, пойманная особью Zoophagus, таллом которой разрастается внутри тела животного (125х); G — Zoopage, Z. tham- nospira с двумя амебами (500х)
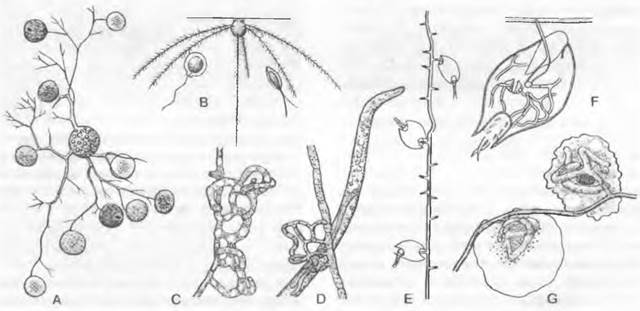
Обзор базидиомицетов. Родственные связи между аскомицетами и базидиомицетами и происхождение их от некоторой общей предковой группы (близкой к Taphrinomycetidae) подтверждаются гомологиями. Как и у аскомицетов, при половом размножении между плазмогамией и кариогамией вклинивается характерная двуядерная стадия (дикариофаза).Стадия парных ядер может возникать не непосредственно после плазмогамии. Более активное из двух ядер может еще пройти через многие клетки мицелия, воспринявшего его, пока не образует с генотипически отличающимся ядром пару ядер (например, у аскомицетов Nеurospora или у базидиомицетов Тyphula). Встречающееся на дикариотичном мицелии многих базидиомицетов образование пряжек гомологично образованию крючков у аскомицетов; многие аскомицеты даже образуют уже пряжки (Тuberасеае) вместо крючков. Незначительное различие между крючком и пряжкой состоит в том, что первые закладываются терминально, а последние — латерально; кроме того, у аскомицетов образование крючков часто ограничено только конечными клетками аскогенных гиф. Напротив, плодовые тела базидиомицетов нельзя го- мологизировать с плодовыми телами аскомицетов, поскольку они состоят только из дикариотичных гиф, тогда как у аскомицетов — из гаплоидных и дикариотичных аскогенных (см. рис. 11.32, А; 11.51, В). Если сплетению этих гиф в плодовые тела у аскомицетов всегда предшествует половой процесс или оба явления тесно связаны между собой, то образованию многих плодовых тел базидиомицетов может предшествовать один акт слияния гиф (соматогамии): однажды возникший дикариотичный мицелий у базидиомицетов может при воздействии соответствующих внешних факторов вновь и вновь образовывать плодовые тела. У аскомицетов дикариофаза ограничена некоторым небольшим числом клеток: в значительной мере жизненный цикл определяется гаплоидными гифами, которые питают дикариотичные гифы. Только у Таphrinomycetidae, которые эволюционно близки к базидиомицетам, дикариотичные гифы питаются независимо от гаплоидных. У базидиомицетов самостоятельная дикариофаза выходит на передний план всего жизненного цикла. Uredinales, рассматриваемые как исходные базидиомицеты, обеспечивают доминирование дикариофазы наличием нескольких типов спороношений, а более эволюционно подвинутые гименомицеты — выживанием дикариотичного мицелия в течение многих лет и вегетативным размножением (оидии, фрагменты гиф и т. д.). В эволюции аскомицетов определенное значение имеет упрощение полового процесса. У базидиомицетов оно достигает своего наивысшего пункта: половые органы больше не закладываются, и соматогамия становится правилом (исключение — Uredinales). Также и повторное образование плодовых тел на дикариотичном мицелии базидиомицетов может рассматривался как ограничение полового процесса.
1. Приложение к Mycobionta: несовершенные грибы (Deuteromycetes)
Естественная система грибов основывается, в частности, на ходе их развития и связанных с половым размножением структур полового спороношения (телеоморфы). Однако у многих грибов (порядка 30 000 видов) известны только особенности их бесполого размножения посредством конидий — бесполое спороношение (анаморфа). При этом следует различать два случая: или половое спороношение нам неизвестно, или у гриба утрачена способность к нему. Все эти грибы объединяются в искусственную группу несовершенных грибов, или дейтеромицетов (Fungi imperfecti, или Deuteromycetes).
Несмотря на отсутствие половой стадии, генетическая рекомбинация может осуществляться благодаря наличию парасексуалыюго цикла (происходящее иногда слияние гиф разных мицелиев: слияние ядер из этих разных мицелиев в диплоидные гетерозиготные ядра; митотический кроссинговер; гаплоидизация путем последовательной утраты хромосом).
Накопление знаний, касающихся общего набора признаков (типы гиф и септ, химизм и ультраструктура клеточной стенки, обнаружение соответствующих телеоморф), а также анализ ДНК позволяют все больше и больше распределять несовершенные грибы по классам и порядкам в естественной системе грибов. Подавляющее большинство их относится к аскомицетам и лишь немногие — к базидиомицетам.
Предварительное искусственное подразделение несовершенных грибов, служащее практическим целям, основывается на структурах, образующих конидии. Конидии возникают почтивсегда на конидиеносцах, которые располагаются поодиночке, в виде палисадного слоя или внутри вместилищ (пикнид).
1. Sphaeropsidales. Конидии образуются в напоминающих перитеции вместилищах (пикнидах) или полостях (камерах). Septoria apii вызывает септориоз («ржавчину») сельдерея. Сюда относятся также Fusicoccum (см. 7.6.4.1 и 9.3.3) и Lasiodiplodia (см.7.6.6.2).
2. Melanconialcs. Конидии возникают на строматическом ложе. Cloeosporium fructigenum вызывает горькую гниль яблок (Sphaceloma, см. 7.6.3.1).
3. Mondiales (Hyphomycetales). Конидии не на строматических ложах; они развиваются на конидиеносцах, часто сильно разветвленных, которые располагаются поодиночке или в пучках (ко- ремиях) либо же объединены вместе со стерильными гифами в слизистые ложа (спородохии). Примеры; Aspergillus, Pénicillium с многочисленными видами; для некоторых видов известна связь с половой стадией (см. Eurotiales). Сюда также относятся Histoplasma, Trichophyton, Arthrobotrys, Dactylella и Dacrylium (см. бокс 11.4). Fusarium oxysporum f. lycopersici вызывает фузариозное увядание растений томата. (См. также Cercospora, 7.6.4.1.)
4. Blastomycetales. Дрожжевидно-почкующиеся формы без половых стадий, например, Cryptococcaceae (близкородственные аскомицетам) (Candida и Torulopsis, см. бокс 11.4).
5. Mycelia sterilia. Мицелии, для которых неизвестно никаких специальных репродуктивных структур (например, некоторые микоризные грибы, склероции, ризоморфы).
2. Приложение к Mycobionta: лишайники (Lichenes)
В лишайниках гифы определенных видов грибов образуют с автотрофными водорослями (см. 9.2.4) симбиотическое объединение, которое представляет собой морфологическое и физиологическое единое целое. Находящиеся в лишайниках эктосимбиотические водоросли (фотобионты) — одноклеточные или нитчатые представители уже обсуждавшихся цианобактерий (например, Chroococcus, Gloeocapsa, Scytonema, Nostoc) или зеленых водорослей (например, Соссотуха, Cystococcus, Trebouxia, Chlorella и Trentepohlia). Грибы (микобионты), участвующие в образовании лишайников. — это прежде всего аскомицеты (чаще виды, образующие апотеции, реже — виды, образующие перитеции), и лишь в очень редких случаях базидиомицеты (например, Сorticiасеае, Сlavariасеае). Принадлежность лишайниковых грибов к различным классам в системе грибов явно говорит о том, что лишайниковые симбиозы неоднократно возникали в разных эволюционных линиях. Этим обусловлено появление новой формы организации таллофитов с собственными признаками. В результате совместного существования гриба и водоросли развились определенные морфологические и химические особенности. Лишайниковые грибы утратили в условиях симбиотического существования свою самостоятельность; они могут расти в природе только совместно с соответствующими водорослями. На этом основании лишайники рассматривались раньше как отдельная систематическая единица— как отдел Lichenes.
Рис. 11.63. Лишайники: А — Dictyonema pavonia; В — Cladonia rangiferina, С — Cladonia pyxidata, слоевище с бокаловидными подециями; D — Thamnolia vermicularis; Е — Dermatocarpon miniaturrr, F — Graphis scripta; G — Parmelia acetabulum; H — Rhizocarpon geographicum-, J — Roccella boergesenir, К — Usnea florida (A — К— 1/2x)

Морфология. Облик лишайника в редких случаях зависит от строения водоросли, чаще же всего — от строения гриба. Примерами первого случая могут быть нитчатые лишайники (например, Ерhebe), у которых гриб опутывает нитчатую цианобактерию, или слизистые лишайники. У подавляющего большинства родов именно гриб определяет облик лишайника. У медленно растущих накипных, или корковых, лишайников, которые живут на поверхности камня, почвы или коры деревьев, слоевище прочно соединено с субстратом. Чаше всего в определенной мере оно проникает в субстрат и имеет чаше всего четко выраженный облик (рис. 11.63, Н). Плоское, преимущественное рассеченное на лопасти слоевище листоватых лишайников (G) соединено с субстратом тяжами (ризинами)1. У некоторых листоватых лишайников (Е) дисковидное слоевище прикрепляется только в середине. Наконец, кустистые лишайники прикрепляются к субстрату очень небольшим основанием и ветвятся наподобие кустиков (J). Слоевища арктоальпийского вида Тhamnolia vermicularis(D) свободно лежат на земле и лишь частично могут прикрепляться немногими гифенными тяжами. У видов рода Сladonia (В, С) от обычно слаборазвитого первичного слоевища листоватого типа поднимаются выросты — подеции — в виде бокальчиков или кустиков, которые несут апотеции.
1 Слоевище листоватых лишайников может прикрепляться к субстрату не только ризинами (представляющими собой выросты сердцевины), но и ризоидами (гифами, идущими от нижней коры) или гомфом (единственной прикрепляющей подошвой). — Примеч. пер.
Гистология и физиология.Поперечный разрез через слизистый лишайник (рис. 11.64, А) показывает более или менее равномерное распределение водоросли и гриба в слоевище (гомеомерное строение); в слизи колонии Nostos здесь обильно развиваются гифы гриба. Гифы часто более плотно смыкаются между собой на верхней и нижней стороне и могут образовывать коровый слой. У кустистых и листоватых лишайников (рис. 11.64, В), а также у многочисленных накипных лишайников водоросли образуют определенный слой, расположенный параллельно поверхности слоевища (гетеромерное строение). В верхнем коровом слое гифы гриба часто смыкаются между собой в прочные сплетения. У листоватых и кустистых лишайников коровые слои чаще всего дифференцированы сильнее, чем у накипных лишайников (ср. В и С). У эндофлеоидных (живущих внутри коры деревьев) и эндолитных (живущих в трещинках камня) лишайников слоевище проникает внутрь субстрата столь глубоко, что едва выступает на поверхность.
Рис. 11.64. Лишайники: A — С — поперечные разрезы слоевищ: А — Collema pulposum (200х), В — Sticta fuliginosa (250х), С — Graphis dendritica (200х); D — соредий Parmelia sulcata(450х); Е — J — гаустории: Е - аппрессории, F — оплетающие гифы, G — внутриклеточный гаусторий, Н — внутримембранный гаусторий, J — окруженный отложением целлюлозы внутримембранный гаусторий (Е, F — 450х; G — J — 600х); К — Peltigera aphthosa (200х)

Гриб и водоросль живут в тесном симбиозе друг с другом, причем гриб опутывает клетки водоросли (рис. 11.64, Е, F) и внедряется в них. При этом часто возникают гаустории — выросты гриба внутрь клетки водоросли (G). Гриб, как правило, остается отделенным от протопластов водорослей, так как они отграничивают места внедрения стенками. У многих лишайников грибы образуют только внедряющиеся в стенки водорослей апрессории (Е), причем водоросль проявляет защитную реакцию в виде утолщенных стенок (J).
У некоторых лишайников наблюдаются также водоросли второго рода, которые существенно отличаются от первых по систематическому положению. Эта вторичная водоросль находится или в определенных местах внутри слоевища (например, у Solotina сгосеа), или же в маленьких головчатых структурах на его поверхности — так называемых цефалодиях (рис. 11.64. К): последние содержат азотфиксирующие синезеленые водоросли (например, Nostoc) и встречаются у лишайников, которые в остальной части слоевища содержат только зеленые водоросли (Chlorophyta). К нормальной симбиотической ассоциации водоросли и гриба может присоединиться второй гриб, который существует как «парасимбионт» или даже как настоящий паразит; известно большое количество таких «лишайниковых паразитов». Наконец, закономерно встречаются лишайники-паразиты в слоевищах других видов.
Углеродный обмен гриба (микобионта) полностью зависит от водоросли (фотобионта). Грибы получают от них чаще всего сахара или сахароспирты. Заключенные в сплетении гиф гриба водоросли нуждаются в получении от него воды и минеральных веществ. Кроме того, гриб обеспечивает защиту от слишком интенсивного света. С симбиотическим образом жизни взаимосвязано наличие многочисленных специфических лишайниковых веществ, которые никогда не образуются изолированными друг от друга партнерами; они откладываются преимущественно на внешней стороне гиф и придают лишайникам ту или иную характерную окраску. Это вещества самой разной природы: алифатические кислоты, депсиды, депсидоны, хиноны, производные дибензофурана и т. д.
Размножение. Водоросли в слоевище лишайников размножаются только вегетативно; их клетки здесь крупнее, чем при жизни в свободном состоянии, так как, очевидно, при симбиотическом образе жизни у них подавляется клеточное деление. Однако грибы развивают характерные плодовые тела (апотеции, перитеции, псевдотеции). Их гимений чаще всего не содержит водорослей. Таким образом, новое слоевище может образоваться при этом только тогда, когда прорастающий из споры мицелий случайно встретится с соответствующей водорослью. Уже удалось осуществить в отдельных случаях экспериментально подобный «лишайниковый синтез». Только у немногих лишайников (например, Еndocarpon) в гимении располагаются водоросли, которые при выбрасывании спор могут увлекаться вместе с ними, так что начинающий развиваться гриб уже имеет в распоряжении нужную водоросль. Какую функцию выполняют пикнидии, встречающиеся у многих лишайников, еще неясно. Размножение лишайников осуществляется у листоватых и кустистых форм часто (в наших местах преимущественно) вегетативным путем. Прежде всего для этого служат соредии (рис. 11.64, D) — оплетенные гифами гриба клетки водоросли, которые часто образуются в определенных местах слоевища — соралях — и распространяются ветром, чтобы на подходящем субстрате снова вырасти в лишайник. У других видов на поверхности слоевища возникают выросты в виде палочек или кораллов (изидии), которые легко отламываются и также служат для вегетативного размножения. Наконец, у лишайников каждый отломанный кусок может снова вырасти в нормальное слоевище.
Встречаемость и образ жизни. Лишайники растут на самых разных субстратах: скалах, почве, коре лиственных и хвойных деревьев, мертвой древесине и т. д. В тропиках мелкие, так называемые эпифилльные, лишайники обитают также на листьях. Живущие на каменистом субстрате накипные лишайники, которые могут растворять известняк (но не кварц), как первые поселенцы подготавливают субстрат для высших растений. Небольшое число видов лишайников обитает как на суше, так и в пресной воде, некоторые живут в море или в зоне попадания морских брызг (см. рис. 11.102). Наиболее пышно лишайники разрастаются в горных лесах умеренной зоны, в условиях высокой влажности воздуха и в туманных лесах тропических высокогорий, а также в тундрах, где почва часто на большом протяжении покрыта преимущественно лишайниками; они образуют здесь собственные растительные формации. Лишайники в целом избегают каменных пустынь больших городов, где они повреждаются газами, загрязняющими атмосферу (прежде всего SO2). Из-за своей различной чувствительности (некоторые накипные лишайники даже в значительной мере устойчивы) они могут служить как индикаторы уровня загрязнения воздуха.
Поглощение воды осуществляется гифами гриба. Особенно у крупных листоватых лишайников нередко часть гиф не увлажняется, так что даже при полном насыщении слоевища влагой сохраняется его аэрация; иногда на нижней стороне слоевища регулярно встречаются дыхательные поры (цифеллы). Обитатели незатененных поверхностей скал выносят не только большой нагрев (до 70 °С), но и полное высыхание на протяжении месяцев. Некоторые почвенные лишайники находятся в высушенном состоянии почти половину дней в году и тогда тоже метаболически неактивны; однако при увлажнении фотосинтез возобновляется уже через несколько минут (пойкилогидричные растения, см. 6.10.3.6). Лишайники с синезелеными водорослями в качестве фотобионтов нуждаются для фотосинтеза в жидкой воде, лишайники с зелеными водорослями могут довольствоваться и водяными парами.
Рост лишайников происходит по сравнению с другими слоевцовыми растениями очень медленно. Даже крупные листоватые и кустистые лишайники наших широт нарастают в год не более чем на 1 — 2 см. У растущего на скалах в альпийской зоне накипного лишайника Rhizocarpon geographicum (см. рис. 11.63, Н) при опре
деленных условиях ежегодный прирост составлял примерно 0,5 мм. По среднему возрасту таких обитающих на скалах накипных лишайников определяли возраст послеледниковых морен. Продолжительность жизни лишайников колеблется от одного года (эпифильные лишайники в тропиках) до нескольких сот, возможно, даже тысяч лет (арктоальпийские накипные лишайники, обитающие на каменистом субстрате).
Лишайники как форпосты жизни внедряются в холодные пустыни высокогорий, а также Арктики и Антарктики; некоторые могут выдерживать охлаждение до -196°С без повреждения и при -24 °С еще связывать СO2.
В ископаемом состоянии лишайники известны только с третичного периода (янтарь), однако уже как различные высокоразвитые виды, практически не отличающиеся от современных.
Применение. Cetraria islándica (исландский мох) — вид, распространенный в сухих лесах и пустошах от тундры до высокогорий, используется как целебное растение (отхаркивающее средство). Из нескольких лишайников недавно были выделены антибиотики. Считается, что обитающая в степях Северной Африки и Ближнего Востока «манна небесная» (Lecanora esculenta) — лишайник, имеющий слоевища от мелколопастных до клубневидных, — может использоваться в пищу. Некоторые лишайники, как, например, виды Roccella (см. рис. 11.63, J) в Северной Африке и на Канарских островах, дают краситель лакмус. Из Cladonia stellaris, чаще всего собираемой в Северной Европе, делают Dauerkranze. Evemia prunastri(дубовый мох) дает ароматическое вещество (mousse de сhkne). Кладония оленья (Cladonia rangiferina, В) наряду с другими кустистыми лишайниками составляет основной корм северного оленя. Letharia vulpina — желтый эпифитный кустистый лишайник, в основном распространенный в альпийской зоне, — единственный в Европе ядовитый лишайник; раньше он использовался для травли волков.
Систематика. Отдельные классы и порядки лишайников располагаются в филогенетической системе согласно систематическому положению микобионтов (ср., например, Lecanorales). Разграничение порядков и семейств лишайников, которых известно около 400 родов и более 20 000 видов, осуществляется по строению плодовых тел гриба, так как здесь мы имеем признаки, наиболее надежные для подразделения на основе родства. Важный признак — реакция на йод, если гифы или сумки окрашиваются в синий цвет, их называют амилоидные. Для различения видов служат и другие химические признаки.
Geosiphon pyriforme — замечательный пример осуществляющегося в настоящее время эндоцитобиоза между грибом (из класса зигомицетов) и цианобактерией типа Nostoc. В этом случае совместное существование в отличие от циклических эндоцитобиозов каждый раз устанавливается заново. Изучение этой чрезвычайно важной ассоциации проливает свет на возможный путь приобретения хлоропластов путем эндоцитобиоза и, кроме того, может служить моделью формирования отдельных элементов клетки. Однако из-за эндосимбиотического образа жизни фотобионта в гифах гриба Geosiphon не следует относить к лишайникам. То же самое можно сказать о соответствующих ассоциациях между слизевиками (Myxobionta) и водорослями в условиях культуры.
I. Ascolichenes (см. класс Ascomycètes)
Порядки лишайников очень близки к соответствующим порядкам грибов в системе аскомицетов и отчасти уже обсуждались при рассмотрении последних (например, Lecanorales). Также мы встречаем порядки с переходами между лихенизированными формами и такими формами, которые образуют с водорослями морфологически высокоразвитые слоевища лишайников. Как и для аскомицетов, в основе разделения на порядки лежит строение сумки. За Caliciales с часто прототуникатными сумками следуют Ostropales и Graphidales с унитуникатными, Lecanorales — с преимущественно унитуникатными, а также Ругеnulales, Verrucariales и Arthoniales — с биту-
никатными сумками. Dothideales отличаются битуникатными сумками в плодовых телах, имеющих чисто асколокулярное развитие (см. Ascomycètes)
1. Порядок: Caliciales. В этом порядке имеются и нелихенизированные представители; лихенизированные формы образуют накипные, листоватые или кустистые слоевища. Апотеции чаще всего на хорошо выраженной ножке. Сумки распадаются, так что споры с растущими дальше парафизами образуют рыхлую массу (мазедий). Зрелые споры — от одно- до многоклеточных — находятся чаще всего по 8 в сумке. Виды рода Calicium встречаются прежде всего на коре, рода Sphaerophorus— преимущественно на силикатном каменистом субстрате.
2. Порядок: Ostropales. Сюда относятся накипные лишайники с апотециями. Бесцветные амилоидные или коричневые аскоспоры могут быть разделены как поперечными, так и продольными и поперечными перегородками; они находятся в количестве от 1 до 8 в неамилоидных сумках. Плодовые тела развиваются, как и у Caliciales, по гемиангиокарпному типу. Апотеции — или широко раскрывающиеся, дисковидные, или узкие, бокаловидные, на ножках — погружены в слоевище или в бородавочки на слоевище.
3. Порядок: Graphidales. Апотеции чаще всего в виде черточек, реже округлые, погруженные или сидячие. Сумки располагаются между простыми или разветвленными парафизами. Водоросли относятся к Chlorococcales или это виды рода Trentepohlia. Примерно 300 видов рода (Graphis обитают преимущественно в тропиках и субтропиках на коре деревьев; в Германии на коре буков распространен «письменный лишайник» (Graphis scripta, см. рис. 11.63, F), апотеции которого напоминают старинные письмена.
4. Порядок: Lecanorales. Этот очень разнообразный по строению слоевища обширный порядок (несколько подпорядков) отличается округлыми, часто блюдцевидными апотециями. Стенка сумки чаще всего толстая, унитуникатная (см. Ascomycètes), у Lichininae прототуникатная, у Peltigerinae двухслойная. Фотобионты — зеленые водоросли (например, Trebouxia, Соссоmуха) и синезеленые водоросли (например, Nostoc).
Виды рода Ephebe (Lichininae) образуют войлочные нити слоевищ на влажных силикатных породах в горах средней высоты и на высокогорьях. Род Соllеmа (Collematinae) отличается слизистыми, напоминающими листья слоевищами с видами Nostoc в качестве симбионтов; слизистые лишайники (см. рис. 11.64, А) обитают на камне и почве в равнинных местностях до снежного пояса.
Представители последующих родов в пределах Lecanorales имеют гетеромерные слоевища. Накипному типу слоевища соответствуют роды Pertusaria, Rhizocarpon (географический лишайник, рис. 11.63. Н) и Lecidea. Слоевища от накипно-чешуйчатых до чисто листоватых имеют виды Xanthoria, Acarospora и Lecanora; слоевища видов рода Xanthoria, часто имеющие окраску от желтой до оранжевой, встречаются в местах, богатых азотом. Acarospora sinopica растет на силикатном субстрате, богатом тяжелыми металлами (см. также Acarosporetum, 6.2.2.4). К листоватым лишайникам относятся виды Parmelia и Physcia. К Peltigerinae, часто имеющим весьма крупные слоевища, относятся: Peltigera — с расположенными по краям слоевища щитовидными апотециями и у некоторых видов с цефалодиями: Lobaria pulmonaria, растущий только в условиях чистого воздуха эпифит на старых лиственных деревьях, преимущественно в горах; Sticta с продырявленной цифеллами нижней стороной слоевища; Sobrina с расположенными по всему слоевищу апотециями и часто также с синезелеными водорослями в слоевище, которые нередко могут в значительной степени вытеснять зеленые водоросли. Умбиликариевые (Umbilicariinae; Umbilicaria\слоевище в диаметре до 15 см и более) обитают на силикатных скалах в горных местностях холодной и умеренной зон. Кустистые лишайники представлены следующими родами: Alectoria, Ramalina; / etharia; Evernia; Anaptychia; Usnea (бородач, К) — со слоевищами в виде длинных (до 8 м) бород, свисающих с ветвей деревьев; Thamnolia (червячный лишайник, D) — на всех высокогорьях и в арктических областях; Cladonia и Stereocaulon (Cladoniinae; В, С) представлены многочисленными разнообразнейшими формами, распространенными по всем климатическим зонам; сюда же относится и исландский мох (Cetraria islándica).
5. Порядок: Pyrenulales. Накипные лишайники с перитециями, развивающимися по аскоги- мениальному типу. Многоклеточные споры располагаются обычно по 8 в сумках, которые находятся между нитевидными парафизами. Фотобионты — виды Trentepohlia. Многочисленные виды рода Pyrenula встречаются на коре деревьев прежде всего в тропиках и субтропиках.
6. Порядок; Verrucariales. Лишайники с сидячими или, чаще, погруженными в слоевище апотециями, которые развиваются по аскогимениальному типу. Настоящие парафизы отсутствуют. Булавовидные или цилиндрические сумки содержат 1 — 8 бесцветных или коричневых спор. Водоросли — Chlorococcales. Многие виды Verrucaria могут быть от эндо- до эпилитов на известковых скалах, некоторые вилы живут постоянно или временно под водой в пресных ручьях и по морским побережьям. Слоевища видов Dertnatocarpon, от чешуйчатых до листовидных, поселяются на скалах. Виды рода Endocarpon обладают муральными спорами (как на рис. 11.41, Е).
Если во всех ранее названных порядках (1 — 6) плодовые тела развиваются по аскогимениальному типу, в последующих порядках (7 — 8) имеются отклонения от этого распространенного варианта.
7. Порядок: Arthoniales. Лишайники самого разного облика (от накипных до кустистых) с округлыми (апотеции) до штриховидных (гис- теротепии) плодовыми телами. Сумки от булавовидных до яйцевидных, споры от двух- до многоклеточных. Водоросли: преимущественно Trentepohlia. Примерно 500 видов рода Arthoma, обитают в основном в тропиках, имеют тонкие накипные, живущие обычно на коре деревьев слоевища: апотеции, от округлых до звездчато-рассеченных на лопасти, не имеют края и эксципула. Также многочисленные накипные виды рода Opegrapha с округлыми до штриховидных черными плодовыми телами в стерильном эксципуле широко распространены по всему земному шару на коре деревьев, древесине и каменистом субстрате. Виды рода Roccella (рис. 11.63, J) образуют кустистые слоевища и заселяют преимущественно скалы по побережьям теплых морей.
8. Порядок: Dothideales. Этот порядок отличается псевдотециями, которые в своем развитии следуют асколокулярному типу. Гифы гриба рыхло перемешаны с различными водорослями.
II. Basidiolichenes (см. класс Basidiomycetes)
Долгое время были известны лишь немногие тропические представители порядка Porrales, которые были ассоциированы с цианобактериями, например, пантропический эпигейный вид Dictyonema pavonia (= Cora pavonia; см. рис. 11.63, А). К настоящему времени базидиомицетные лишайники найдены как в тропиках, так и в умеренной зоне. Их компонентами являются Clavariaceae или Agaricales (Omphalina) и зеленые водоросли (Соссоmуха и др.).