БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
Шестое подцарство: Rhodobionta
Представители этого подцарства имеют пластиды, которые напоминают пластиды у Glaucobionta. Однако оболочка хлоропласта уже не содержит пептидоглюкан. Клетки, имеющие жгутики, полностью отсутствуют.
Единственный отдел: красные водоросли (Rhodophyta)
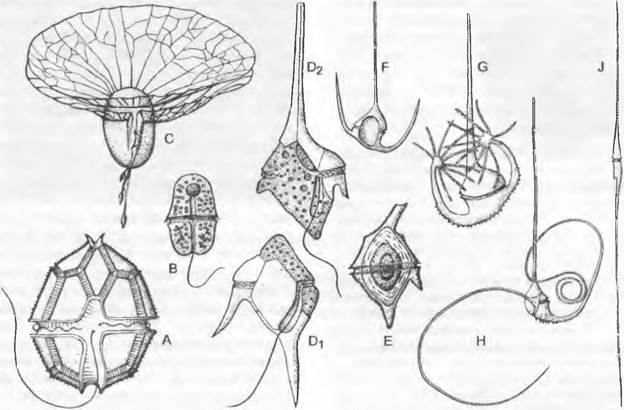
Красные водоросли, обитающие преимущественно в морях, бывают от светлокрасного до фиолетового цвета, изредка от темно-пурпурного, коричнево-красного до почти черного или даже от сине- до оливково-зеленого. Формы или стадии, имеющие жгутики, такие, как монадные виды, зооспоры или сперматозоиды, отсутствуют. Одноклеточные представители встречаются только в достаточно обособленном подклассе Ваngiophycidае. Как в этой небольшой группе, так и среди других красных водорослей преобладают представители с талломами в виде отдельных нитей и сплетения нитей; другие формы имеют псевдопаренхиматозное (ложнотканевое) строение. Настоящие ткани полностью отсутствуют. Сплетения нитей и псевдопаренхимы (= плектенхимы) красных водорослей образуются по одноосевому (центральнонитчатому) или многоосевому (фонтанному) типу (см. 5.3.1; рис. 5.6, 5.7). В клетках, почти без исключения одноядерных, находятся чаще всего многочисленные дисковидные, овальные или лопастные хроматофоры (пластиды), которые, однако, никогда не бывают бокаловидными.
Пластиды. В них хлорофилл а (хлорофиллов b и с нет; наличие хлорофилла d долгое время считалось спорным) и сопутствующие каротиноиды маскируются красными, сильно флуоресцирующими пигментами, которые находятся в так называемых фикобилисомах. Фикобилисомы (они имеются и у прокариотических цианобактерий) представляют собой дисковидные или шаровидные тельца размером 30 — 540 нм. Они располагаются в хроматофорах на тилакоидах и содержат водорастворимые фикобилипротеиды с простетическими фикобилинами, определяющими характер окраски. У красных водорослей она обусловлена преимущественно красным фикоэритрином; в фикобилисомах содержится также фикоцианин (см. рис. 2.89, В, С).
Бокс 11.5. типы организации эукариотных водорослей
Виды Glaucophyta, Rhndobionta и примыкающих к ним групп (например, Heterokontophyta), как и примитивно устроенные представители Chlorobionta (Chlorophyta, Streptophytina), трактуются как водоросли. Эукариотные водоросли (т. е. водоросли, в узком смысле слова, без прокариотных синезеленых водорослей) — это одноклеточные или многоклеточные, по- разному окрашенные, первично фотоавтотрофные растения, чаще всего имеющие талломную организацию и в основном обитающие в воде. Их хлоропласты содержат фотосинтетические пигменты наряду с дополнительными пигментами. Пластиды всех эукариотных водорослей содержат хлорофилл а и большей частью еще и другие формы хлорофилла (табл. 11.3). Для оксигенного фотосинтеза в качестве донора электронов служит вода, при этом высвобождается кислород. Под дополнительными пигментами понимают различные каротиноиды (а также встречающиеся у немногих групп фикобилины, см. 6.4.2). Часто в хлоропластах или вне их имеются пиреноиды (см. 2.2.9.1). Хлоропласты ограничены либо двумя мембранами — простые пластиды, либо 3 — 4 мембранами — комплексные пластиды. У представителей некоторых групп водорослей (Cryptophyta, Chlorarachniophyta) комплексные пластиды содержат нуклеоморфу, которая рассматривается как редуцированное ядро эукариотного эндосимбионта. Согласно теории эндосимбиоза (см. 2.4.2) этот процесс обусловливает появление пластид в клетке хозяина. Предполагается, что простые пластиды (у Glaucophyta, Rhodophyta, Chlorophyta) произошли в результате единственного первичного эцдоцитобиоза от прокариотных водорослей, роль которых отводится Cyanophyta. Первичный эндоцитобиоз был, возможно, уникальным эволюционным событием.
Комплексные пластиды (все другие отделы эукариотных водорослей), напротив, свидетельствуют о том, что они возникли в результате двух этапов эндосимбиоза (см. 2.4). После первичного эндосимбиоза происходили дальнейшие независимые события эндосимбиоза (вторичный эцдоцитобиоз): уже эукариотные водоросли включались в клетки хозяина. Для водорослей с нуклеоморфой в пластидах такой вторичный эндоцитобиоз считается доказанным. Итак, эукариотные водоросли полифилетичны как в отношении клетки- хозяина, так и в отношении приобретения пластид в результате первичного или вторичного эндосимбиоза.
Родство пластид эукариотных водорослей и фотосинтетического аппарата прокариотных водорослей подтвердил анализ последовательностей малых субъединиц рРНК (SSU).
Органы, образующие гаметы и споры, не имеют многоклеточных покровов и чаще всего у них также отсутствуют постгенитальные, т. е. вырастающие позже, покровы. Спорообразуюшие органы (спорангии) всегда, а гаметангии чаще всего одноклеточные. Голые гаметангии водорослей в отличие от снабженных многоклеточными стенками и поэтому, защищенных антеридиев и архегониев мхов и папоротников (см. бокс 11.8) носят названия спермогонии (♂: со сперматозоидами, имеющими жгутики; см. рис. 11.73, Е: s) или же сперматангии (♂: со спермациями, лишенными жгутиков; см. рис. 11.67, D) и оогонии ♀, с яйцеклеткой: см. рис. 11.73: о) или же карпогоны (♀: с особым развитием после оплодотворения: см. рис. 11.67, F — I).
Зиготы никогда не развиваются внутри, женских половых органов в многоклеточные зародыши. В большинстве групп водорослей репродуктивные клетки (гаметы, споры) имеют жгутики, хотя у некоторых более высокоразвитых групп жгутики имеются только у мужских гамет; лишь у немногих групп водорослей вообще не образуется жгутиковых стадий (Реnnales, Rhodophyta, Zygnematophyусеае). Жгутики имеют характерную для эукариот структуру 2 + 9 (см рис. 2.16). Они обычно парные, причем один может быть направлен вперед по ходу движения, а второй — назад. Два жтутика могут быть одинаковой или разной длины, оба гладкие или разные: один гладкий, а другой снабжен перисто-расположенными волосками (перистый жгутик).
Водоросли в ходе эволюции прошли путь прогрессивного развития от одноклеточных форм до сложных ложнотканeвых или тканевых талломов (см. гл. 5). Формы на уровне слоевцовых растений (таллофитов) еще не показывают дифференциации вегетативного тела на «настоящие» листья, стебли и корни. Несколько сходные образования у некоторых высокоразвитых водорослей не имеют проводящих структур, которые можно было бы сравнить с проводящими пучками сосудистых растений (например, у бурых водорослей есть лишь отдельные проводящие элементы, напоминающие ситовидные трубки). По большей части структуры, отдаленно напоминающие основные органы высших растений, не имеютанатомической нифференциации, поэтому их обозначают как филлоиды, каулоиды и ризоиды (см. 5.3.2).
Можно различать следующие морфологические группы разной высоты организации (уровни организации).
• Амебоидный (= ризоподнальиый) уровень. Одноклеточные голые водоросли образуют псевдоподии, с помощью которых они захватывают твердые частицы пищи. Если эти придатки тонкие и нитевидные, их называют ризоподиями (см. рис. 11.74, С). Встречаются также объединения таких клеток.
• Монадный уровень. Одноклеточные водоросли со жгутиками, чаще всего снабженные глазками и пульсирующими вакуолями («флагелляты»; см. рис. 11.103). После клеточных делений они могут образовывать колонии, состоящие из нескольких (до очень многих) клеток (см. рис. 11.95, в; 11.74, F). Пальмеллоидная стадия (стадия пальмеллы) представляет пальмеллоидный (капсальный) уровень: при делении клеток не образуется новых жгутиков, а дочерние клетки погружены в общую слизь (см. рис. 11.75, С).
• Капсальный (= тетрасноральный) уровень. В рудиментарном виде еще сохранились различные признаки монадного уровня. Так, жгутики, если они еще сохранились, негнущиеся или редуцированные, в любом случае не способные к активному движению, могут быть только у репродуктивных клеток. Поскольку клетки после деления погружены в общую слизь, возникают колонии, которые могут быть даже вытянуты в нитевидные структуры (см. рис. 11.75, D). Клеточная стенка тонкая или отсутствует.
• Коккоидный уровень. Вегетативные клетки не имеют жгутиков и покрыты клеточной стенкой. У них нет даже никаких остатков монадной организации. Могут быть одноклеточные формы, ценобии или колонии в более узком смысле (см. рис. 11.76; 11.97, А; 11.98, А).
• Нитчатый (= трихальный) уровень. Одноклеточные (моноэргидные) клетки образуют разветвленные или неразветвленные нити, нарастающие интеркалярно или верхушечно (см. рис. 11.89, А).
• Сифонокладальный уровень. Клетки, образующие нити, содержат по нескольку ядер; многоядерные клетки называют полиэнерги- дами.
• Сифональный уровень. Макроскопический таллом представлен одной -единственной крупной многоядерной клеткой — нитевидной или другого облика. Размеры таллома могут быть достаточно большими (см. рис. 11.73, О; 11.91; 11.92, D).
• Ложнотканевой таллом. Боковые оси (нити) переплетаются друг с другом. Клетки часто слипаются или даже срастаются (см. 5.3.1; рис. 5.7, А).
• Тканевой таллом Клетки, делящиеся в разных направлениях, остаются связанными меж ду собой, образуя ткань (см. 5.3.2; рис. 5.4, G).
Кратко охарактеризованные здесь уровни организации достигались в разных эволюционных линиях водорослей независимо друг от друга, и их развитие происходило неодинаково. Тканевой таллом, например, появился в примитивной форме у Ulvophyсеае и в особенно развитой форме — у Рhaeophусеае, в ложнотканевой форме — у Рhaeophyceae и Rhodophyсеае.
У обоих пигментов имеется несколько вариантов, которые различаются, в частности, абсорбцией и принадлежностью к тем или иным группам (к синезеленым или же красным водорослям). Фикобилисомы у синезеленых и красных водорослей улавливают свет и передают энергию возбуждения на собственно фотосинтезирующий пигмент. Расслоение фикобилинов в фикобилисомах (фиконианины внутри, фикоэритрины снаружи) дает направление переносу энергии. У Сryptophyta, также имеющих фикобилины, фикобилисомы отсутствуют.
В хлоропластах тилакоиды не сгруппированы, а располагаются с равными промежутками между собой (как у цианобактерий и глаукофитов). Двойная мембрана отграничивает хроматофоры от остальной части клетки; эндоплазматическая сеть в этом не участвует.
В качестве запасного вещества откладывается прежде всего багрянковый крахмал в форме округлых, не растворимых в воде, часто слоистых зернышек, дающих с йодом красноватое окрашивание. Это полисахарид, по своим свойствам занимающий промежуточное положение между гликогеном и крахмалом. Зернышки в отличие от крахмала у зеленых водорослей откладываются не внутри хлоропластов, а на их поверхности и в цитоплазме. Встречаются некоторые другие, присущие только красным водорослям вещества («флоридозиды» — соединения галактозы и глицерина), а также капельки масла.
Фибриллярный компонент клеточной стенки состоит преимущественно из целлюлозы, микрофибриллы которой построены из цепочек, но не параллельно расположенных (как у высших растений и некоторых зеленых водорослей), а переплетенных наподобие войлока.
Аморфная часть часто содержит ослизненные галактаны (например, агар; каррагенан = галактансульфат; галактаны — полимеры галактозы).
Жизненный цикл. Для красных водорослей характерна смена трех поколений, при которой за гаплоидным гаметофитом следуют диплоидный карпоспорофит и еще
одно диплоидное спорофитное поколение (преимущественно тетраспорофит).
При диплобионтном типе (три поколения, представленные двумя особями, например, Polysiphonia, рис. 11.65, А) гаметофит представляет собой самостоятельное гаплоидное растение. Он развивает ♀ гаметангий — карпогон. У многих красных водорослей (например, у всех флоридей) его расширенная нижняя часть (брюшко) переходит в трихогину — длинный, чаще всего тонкий воспринимающий орган (рис. 11.66, F: t; 11.67, F: t). На других частях или других особях гаметофитов в сперматангиях (= мужских гаметангиях) возникают лишенные жгутиков мужские (♂) половые клетки — спермации. Они одноядерные, пассивно переносятся токами воды, затем прилипают к трихогине и отправляют в нее свое ♂ половое ядро, после чего это ядро мигрирует к ядру карпогона и сливается с ним (гамето-гаметангиогамия, или нетипичная оогамия).
Рис. 11.65. Rhodobionta, Рhodophyсеае. Смена поколений и ядерных фаз. Трехчленный диплобионтный цикл Роlysiphonia (А); трехчленный гаплобионтный цикл Вtrachospermum (В). Светлые линии — гаплофаза; темные линии — диплофаза; R! — редукционное деление
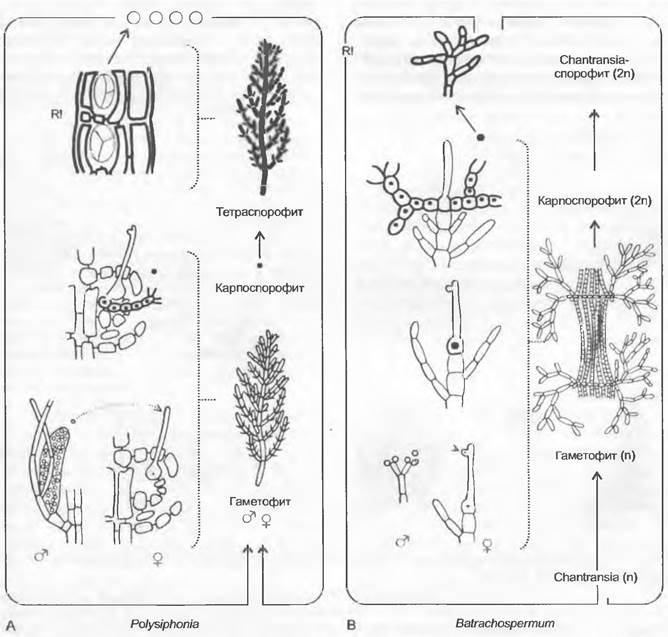
Из оплодотворенной яйцеклетки возникает карпоспорофит в форме диплоидных нитей, разделенных на клетки, которые вырастают из карпогона, при этом, однако, оставаясь соединенными с гаплоидным гаметофитом. Соответственно происходит смена первых двух поколений, разделенных сменой ядерных фаз, на одном и том же растении. Карпоспорофит после исключительно митотических делений дает диплоидные карпоспоры, которые, таким образом, являются митоспорами.
Тетраспорофит. У подавляющего большинства красных водорослей из карпоспоры возникает новое растение, чаще всего морфологически сходное с гаметофитом, однако диплоидное. На нем после редукционного деления из каждой материнской клетки спор образуется 4 гаплоидные тетрамейоспоры (см. рис. 11.65; 11.66, В); это поколение называется тетраспорофит. При переходе от карпоспорофита к тетраспорофиту происходит соответственно без изменения ядерной фазы смена 2-го и 3-го поколений жизненного цикла. Развитие трех поколений осуществляется, таким образом, на двух вегетативных телах (диплобионтное развитие); большинство красных водорослей (за исключением Nemalionales) относится к этому типу.
Рис. 11.66. Rhodophyceae: А, В — Callithamnion corymbosum, образование тетраспор (300х): А — закрытый, В — освободившийся спорангий с 4 мейоспорами; С, D — Bonnemaisoniahamifera, гаметофит и тетраспорофит (5х), С — гаметофит и зачатки цистокарпиев, D — спорофит, также известный как Trailiella intricate; Е — Ceramiales, Platysiphonia miniata,цистокарпий с видными в нем на просвет карпоспорофитами (100х); F, G — Cryptonemiales, Dudresnaya; F — оплодотворенный карпогон, на трихогине (t) которого приклеиваются еще несколько спермациев (s), дает начало разветвленным спорогенным нитям (sf), которые соединяются с шестью ауксилярными клетками (а1—а6), клетки а1—а6 входят в состав ветвей, которые возникают от оси ha (250х), G — зрелое скопление карпоспор
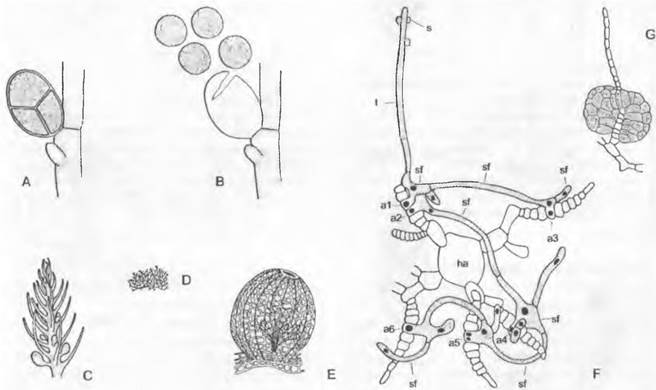
Гаметофит и тетраспорофит имеют чаще всего одинаковый внешний вид, но могут быть и непохожими (см. рис. 11.66, С, D), отчего раньше их относили не только к разным родам, но и даже к далеко отстоящим друг от друга порядкам. Также и паразитирующий на гаметофите карпоспорофит в отдельных случаях выглядит как нечто постороннее в такой степени, что его считали действительно инородным паразитом и давали ему особое название. Гаметофиты бывают однодомными и двудомными. В последнем случае иногда встречаются различия в строении ♂ и ♀ растений. Часто карпоспорофит (= гонимокарп) обрастает специальными покровными нитями гаметофита, в результате чего возникает так называемое плодовое тело — цистокарпий (см. рис. 11.66, Е). Если покров закладывается уже перед оплодотворением карпогона, говорят о прокарпии.
Часто карпоспорофит поддерживается вспомогательными, или ауксилярными клетками, которые, очевидно, имеют значение для физиологии питания. Это богатые цитоплазмой клетки карпоспорофита, расположенные радом с карпогоном, которые воспринимают ядро зиготы (или диплоидное ядро, возникшее после последующих делений) из карпогона, претерпевают последующие митотические деления и наконец продолжают образование карпоспорофита. Отходящие от карпогона соединительные нити (см. рис. 11.66, E: sf) могут достигать многих ауксилярных клеток. При этом диплоидные ядра размножаются и распространяются в гаметофите, так что вслед за одним- единственным актом оплодотворения от гаметофита возникают многочисленные карпоспорофиты, которые могут питаться от него.
Последовательность разных поколений на одном организме (гаплобионтный тип развития) можно проследить на встречающихся в пресной воде видах рода Вatrachospermum (см. рис. 11.65, В). Здесь также происходит трехчастная, гетероморфная и гетерофазная смена поколений, три члена которых, однако, остаются связанными между собой, а именно: 1) диплоидный «Chantransia-спорофит» (предросток); 2) мутовчато разветвленный гаплоидный гаметофит; 3) диплоидный карпоспорофит.
Гаметофит. Однодомный таллом состоит из гаплоидных, мутовчато разветвленных нитей (см. рис. 11.67, А). Многочисленные сперматангии возникают чаще всего по два из конечных клеток, расположенных мутовками боковых ветвей (рис. 11.67, А). Каждый сперматангий состоит всего из одной клетки, все содержимое которой идет на образование одного-единственного округлого бесцветного спермация с большим ядром и очень нежной стенкой (D). Женские органы — карпогоны — сидят между ветвями, несущими сперматангии, также на концах ветвей и представляют собой длинную клетку, которая в нижней части бутылевидно расширена, а в верхней части вытянута в булавовидную трихогину (Е, F). Кар погон с трихогиной глубоко погружен в слизь. Спермаций, пассивно переносимый токами воды, может активно внедряться в эту слизь (механизм неизвестен); при этом он достигает трихогины, в которую выпускает все свое содержимое. Ядро спермация мигрирует в карпогон и сливается с находящимся там женским ядром. После этого базальная часть карпогона с образовавшимся диплоидным ядром отделяется от трихогины слизистой пробкой (G).
Карпоспорофит. Он состоит из разветвленных, разделенных на клетки нитей, которые вырастают из зиготы, однако при этом остаются связанными с гаметофитом (рис. 11.67, Н). Карпоспорофит образует во вздутых конечных клетках по одной округлой митоспоре, содержащей ядро и хлоропласт, — диплоидной карпоспоре. Карпоспоры высвобождаются из остающихся стенок конечных клеток (Iк1, к2) в виде округлых, лишенных жгутиков образований. Они вырастают в Сhantransia-стадию (Сhantransia-спорофит). Это разветвленные диплоидные нити, которые плотно прилегают к субстрату и представляют собой предросток для возникающих позже от них гаметофитов. Сhantransia-стадия, таким образом, еще диплоидна, но собственно гаметофит гаплоиден. Мейоз происходит без образования мейоспор в отдельных клетках нитей Сhantransia. Гаплоидные клетки развиваются затем в мутовчато разветвленные гаметофиты (С).
У некоторых видов в качестве 2-го спорофита образуется маленькое, мутовчато разветвленное растение, которое в определенных местах (после мейоза) непосредственно дает начало гаметофитам; это сближает Сhantransia-стадию со спорофитным поколением.
Рис. 11.67. Rhodophyceae, Nemalionales. Batrachospermum moniliforme:
А — внешний вид (Зх); В — часть таллома гаметофита с мутовкой ветвей (20х); С — диплоидный спорофит (Сhantransia) с двумя сидящими на нем гаметофитами (100х); D — часть ветви гаметофита с 4 сперматангиями, слева вышедший спермаций (540х); Е — зачаток карпогона; F — зрелый карпогон; G — карпогон после оплодотворения спермацием (s), в основании его копуляция половых ядер (о); Н — диплоидный карпоспорофит (g) с гаплоидными покровными нитями (h); I — зрелый карпоспорофит с карпоспорангиями (к); к1, к2 — освободившиеся карпоспорангии; t — трихогина

Распространение и образ жизни. Красные водоросли представлены примерно 4000 видами в составе более чем 500 родов. За немногими исключениями (например, Вatrachospermum, Lemanеа), они обитают в прибрежной зоне морей, особенно более теплых; многие виды очень чувствительны к колебаниям температуры. Часто красные водоросли растут значительно глубже (максимум до 180 м), куда доходит лишь слабый коротковолновый свет, где они не только могут жить как теневыносливые водоросли, но и благодаря наличию дополнительных пигментов (фикобилипротеидов) могут оптимально использовать преобладающий на глубине коротковолновый свет, комплементарный к собственной окраске этих пигментов (см. 6.4.3). Красные водоросли — представители бентоса. Они всегда прочно прирастают специальными нитями или присосками, чаще всего к каменистому субстрату, некоторые растут как эпифиты на более крупных водорослях. Такие эпифиты могут быть очень строго приурочены к определенному роду растения- форофита (например, виды рода Polysiphonia к Аscophyllum). Красные водоросли живут автотрофно; часть из них — бесцветные паразиты, из которых несколько десятков представляют собой очень редуцированные формы, обитающие на других, близко родственных им видах красных водорослей («адельфопаразиты»).
Систематика. Единственный класс красных водорослей — Rhodophyceae. Он подразделяется на подклассы Bangiophycidae и Florideophycidae.
1. Подкласс: Bangiophycidae
Это весьма просто устроенные одноклеточные, нитчатые или листовидные водоросли с ин- теркалярным ростом. Поры обычно отсутствуют. Хлоропласты звездчатые, с пиреноидом.
1. Порядок: Porphyridiales. Объединяет одноклеточные, отчасти колониальные формы. Половое размножение неизвестно. У часто встречающейся наземной водоросли Porphyridium purpureum многочисленные отдельные клетки объединены в слизи.
2. Порядок: Bangiales. К этому порядку относятся нитчатые (например, Bangia) или листовидные (например, Porphyra) формы. Erythrotrichia образует неразветвленные нити, в которых могут возникать вегетативные моноспоронисты, а в них — по одной моноспоре. Сначала моноспоры голые, им свойственно амебоидное движение. Из них вырастают новые нити Erythrotrichia. У Porphyra (пурпурной водоросли) карпогоны чаще всего неотличимы от вегетативных клеток. Оплодотворенная спермациями диплоидная зи гота после нескольких митозов непосредственно делится на некоторое число диплоидных карпоспор; таким образом, карпоспорофитное поколение либо отсутствует, либо сильно редуцировано. Карпоспора прорастает в нитчатое, очевидно. также диплоидное образование, которое внедряется в известковую раковину двустворчатых моллюсков и морских желудей. Начинается развитие так называемой Conchocelis-фазы, которая представляет собой спорофитное поколение. Оно заканчивается образованием конхоспор (гомологичных тетраспорам), после чего, по всей вероятности, происходит редукционное деление. Conchocelis-фаза широко распространена по морским побережьям Европы. К этому же порядку, возможно, относится Cyanidium caldarium — вид, относительно которого дискутируется родственная связь с синезелеными водорослями (цианобактериями)1.
1 Практически во всех современных системах Cyanidium caldarium и некоторые родственные этому виду формы выделяются в отдельный класс Cyanidiophyceae, а он иногда выделяется в самостоятельный отдел Prerhodophyta. — Примеч. пер.
2. Подкласс: Florideophycidae
Представители этого подкласса имеют более сложное строение таллома, основу которого составляют разветвленные нити с верхушечным ростом. Одноклеточные представители не встречаются. Даже простейшие Florideophycidae уже имеют гетеротрихальное строение (т. е. дифференцированы на подошву и вертикальные нити), однако даже наиболее высокоразвитые представители в противоположность бурым водорослям никогда не бывают паренхиматозными (см. 5.3.1), а образуют свой таллом по одноосевому или фонтанному типу (см. рис. 5.6; 5.7). Клетки часто соединены друг с другом «порами». Это отверстия или каналы со структурами в виде пробок внутри; их функция однозначно не выяснена.
1. Порядок: Nemalionales1. Ауксилярные клетки отсутствуют. Этот порядок представлен в Средней Европе видами рода Batrachospermum, растущими преимущественно в быстротекущих, затененных ручьях в форме слизистых масс, напоминающих икру, от буроватого до оливковозеленого цвета. Batrachospermum показывает отклоняющийся от нормального типа гаплобионтный цикл развития. Он также четко отличается от большинства других красных водорослей, тем что обитает в пресной воде, а именно чаще всего в быстротекущих ручьях с высокой чистотой воды.
1 Правильнее назвать этот порядок Nemaliales. — Примеч. пер.
Rhodochorton investiens— красная водоросль, живущая как эпифит на Batrachospermum, имеет, напротив, нормальный диплобионтный цикл развития; гаметофит и тетраспорофит в этом случае в значительной мере сходны между собой. Близкородственный морской вид Rhodochorton purpureum — гаплобионт с двучленной сменой поколений: карпоспорофитное поколение выпадает, а оплодотворенный карпогон непосредственно вырастает в тетраспорофит; последний остается сросшимся с гаметофитом того же облика.
Другой род пресноводных водорослей из этого порядка — Lemanea, тогда как виды родов Nemalion, Bonnemaisonia (см. рис. 11.66, С, D) и Gelidium живут в морях. Систематическое положение последнего рода спорное, так как здесь встречаются клетки, трактуемые как ауксилярные2.
2 В настоящее время роды Ваtrachospermum, Lemanеа и некоторые другие, отличающиеся сходным жизненным циклом (без образования мейоспор), обитанием в пресной воде и отсутствием красной окраски, обычно выделяются в отдельный порядок Ваtrachospermales; род Gelidium выделяется в порядок Gelidiales. — Примеч. пер.
2. Порядок: Cryptonemiales. Ауксилярные клетки закладываются до оплодотворения на специальных пучках ветвей. У Согаllinасеае1 (Согаllinа, Lithothamnion, Lithophyllum) клеточные стенки инкрустированы кристаллами кальцита (образование коралловых рифов); ископаемые представители имеют значение как образователи скальных пород. Сюда относится также Melobesia (см. рис. 5.7, В) с талломом, развивающимся по «фонтанному типу».
1 В большинстве современных систем это отдельный порядок Соrallinales. — Примеч. пер.
3. Порядок Gigartinales. В ауксилярную клетку превращается нормальная интеркалярная клетка таллома. К этому порядку относятся гребневидно-перистый Plocamium, построенная по фонтанному типу Furcellaria (см. рис. 5.7, А) и плоско-вильчатый Chondrus.
4. Порядок: Rhodymeniales. Несущая клетка карпогона отшнуровывает перед оплодотворением дочернюю клетку, а та, в свою очередь, — ауксилярную клетку. Карпогон возникает от прокарпия (состоящего из несущей клетки, дочерней клетки, ауксилярной клетки и карпогонной ветви), который после оплодотворения становится цистокарпием. Сюда относится часто встречающаяся в Атлантике Rhodymenia с листовидным талломом.
5. Порядок: Ceramiales. Ауксилярная клетка обособляется после оплодотворения карпогона от несущей клетки карпогонной оси. Имеются прокарпий (здесь состоящий из несущей клетки, ауксилярной клетки и карпогонной ветви) и цистокарпий, как и у предыдущего порядка. Жизненный цикл соответствует представленной вначале основной схеме (см. рис. 11.65, А). Таллом построен по одноосевому типу и состоит из обильно разветвленных нитей, часто образующих кору.
Особенно сложно расчленен таллом водоросли Delesseria sanguínea, обитающей в Атлантическом океане. Листовидные лопасти ее таллома, отходящие от базального диска, имеют центральную и боковые жилки; осенью пластины отмирают, но основные оси остаются и развивают следующей весной новые пластины. К Сега- miales относятся Grinnellia (см. рис. 5.8, A), Platysiphonia (см. рис. 11.66, Е), а также представленные в Северном и Балтийском морях роды Polysiphonia. Ceramium и Plumaria.
Раньше иногда предполагалось родство красных водорослей с аскомицетами (Муcobionta). Это предположение не было подтверждено данными анализа ДНК. Действительно, обращает особое внимание сходство следующих признаков: ♀ гаметангий (здесь — карпогон, там — аскогон, иногда с трихогиной; здесь — нити карпоспорофита, там — аскогенные гифы); взаимосвязь гаметофитного и спорофитного поколения; отсутствие имеющих жгутики репродуктивных клеток; оплодотворение спермациями (как и у некоторых аскомицетов); слоевища лишайников с порами в септах и способностью к образованию анастомозов; сходство в химическом строении гликогена грибов и багрянкового крахмала; запасной продукт трегалоза. Однако химический состав клеточной стенки различен: здесь целлюлоза, там хитин.
В ископаемом состояниикрасные водоросли встречаются с перми во всех формациях; отнесение Solenopora из ордовика к красным водорослям является спорным.
• Отделы водорослей с комплексными пластидами без хлорофилла b. Последующие отделы водорослей имеют комплексные пластиды, которые возникли в результате вторичного эндосимбиоза. Поскольку эти пластиды не содержат хлорофилла b, но очень часто в качестве дополнительных пигментов присутствуют фикобилины. Предполагается их происхождение от предков, близких к предкам Rhodobionta, что подтверждается сходством состава пластидной ДНК.
1. Приложение к Rhodobionta: отдел криптофиты (Сryptophyta)
Представители этого отдела, за немногими исключениями (пальмеллоидные и нитчатые формы) — жгутиковые организмы (монадный тип организации). Род Biornbergiella отличается нитчатым талломом. Снабженные жгутиками асимметричные клетки, характерные для подавляющего большинства видов, не имеют клеточной стенки, а покрыты только пелликулой, которая состоит из прямоугольных или многоугольных белковых пластинок. На переднем конце выходят два жгутика, несколько различающиеся по длине. Оба жгутика несут перистые волоски: более длинный — в два ряда, более короткий — в один.
Жгутики чаще всего направлены вперед, реже — вдоль тела назад (рис. 11.68, С). Они выходят вместе над глубокой глоткой, которая выложена эджектосомами, сильно преломляющими свет, — это тельца, которые при раздражении выстреливают. По-разному окрашенные (например, синие, сине-зеленоватые, красноватые) хлоропласты содержат хлорофиллы а и с, α- и β-каротин и ксантофилл диатоксантин, а также фикобилины фикоэритрин и фикоцианин. В отличие от красных водорослей и цианобактерий эти пигменты здесь не локализованы в фикобилисомах. Важный запасной продукт — крахмал, который откладывается в пиреноидах. Последние, правда, расположены в складке эндоплазматической сети (= оболочке хлоропласта), однако вне мембраны хлоропласта.
Вегетативное размножение осуществляется путем продольного деления; половое размножение с достоверностью не установлено.
Живущие в морях и пресных водах (примерно поровну) 120 видов распределены по 12 родам.
Хлоропласты можно рассматривать как сильноредуцированные эукариотные эндосимбионты (родственные красным водорослям) на основании наличия сохранившегося недоразвитого клеточного ядра — нуклеоморфы. Она располагается в единственном числе в пиреноиде или у поверхности хлоропласта.
Можно предполагать, что криптофиты исходно имели первичные пластиды типа цианелл (см. Glaucobionta), которые, однако, в ходе дальнейшей эволюции были заменены внедрившимися одноклеточными красными водорослями (путем вторичного эндоцитобиоза).
Отдел содержит единственный класс с единственным порядком — Cryptomonadales. Многочисленные виды рода Cryptomonas (рис. 11.68, А) обитают преимущественно в мезотрофных водоемах.
Рис. 11.68. Сryptophyta: А — Сryptomonas sр.; В — Chilomonas paramaecium; С — Кatablepharis phoenicoston с одним жгутиком, направленным вперед, и другим жгутиком, направленным назад (А — С — 1 200х); с — хлоропласт с несколькими пиреноидами (показан точками); к — ядро; s — глотка; v — вакуоль
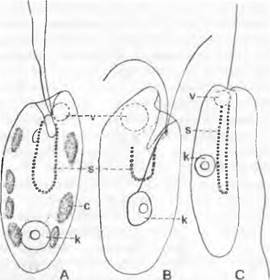
Сапротрофные представители рода Chilomonas (В) бесцветны, однако еще содержат один лейкопласт. Бесцветен и фаготроф Katablepharis (С). Один вид представляет собой сильноредуцированную форму, живущую как эндосимбионт в инфузории Mesodinium rubrum, которая таким образом получила способность к фотосинтезу. Эта симбиотическая водоросль содержит лишь один хлоропласт и немногочисленные митохондрии.
2. Приложение к Rhodobionta: отдел динофиты (Dinophyta, Рyrrhophyta, Dinoflagellata)
В большинстве своем одноклеточные формы, несущие два жгутика с тонкими мастигонемами; известны лишь немногие коккоидные и нитчатые формы. Наряду с вегетативным встречается половое размножение.
Хлоропласты содержат хлорофилл а; у некоторых видов установлено также наличие хлорофилла с. Окраска клеток от желто-бурой до красноватой благодаря наличию дополнительных пигментов, таких, как β-каротин и различные ксантофиллы, из которых наиболее важен перидинин. Оболочка хлоропласта состоит чаще всего из трех мембран, которые не связаны с эндоплазматической сетью ядра. Тилакоиды располагаются в ламеллах по три и не образуют периферической опоясывающей ламеллы (таким образом, картина иная, чем на рис. 11.72, G). Основной продукт ассимиляции — крахмал, который накапливается в виде зернышек вне хло
ропластов. Наряду с ним встречаются также жироподобные вещества. Для некоторых представителей имеются сведения, что исходно имевшаяся простая пластида значительно редуцировалась вплоть до того, что остался один глазок; она стала фотосинтетически неактивной и в результате вторичного эндосимбиоза была функционально заменена комплексной пластидой.
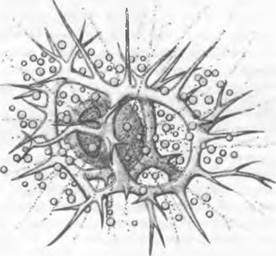
Рис. 11.69. Dinophyta. Зооксантеллы в радиолярии (Eucoronis challengerr, 260х)
Динофиты, возможно, представляют собой гетерогенную группу эукариотных водорослей. Они содержат пластиды очень разного типа. Это взаимосвязано с их способностью к фаготрофии, которая сохранилась и у автотрофных видов. У некоторых видов пластиды не являются постоянными компонентами клеток — это клептохлоропласты. Есть виды, у которых, возможно, имеется нуклеоморфа. Молекулярно-филогенетический анализ ядерного генома дает основание предполагать близкое родство динофитов и относимых к животным жгутиковых (Ciliata, Sporozoa или Apicomplexa). У гетеротрофно живущего возбудителя малярии Plasmodium falciparum уже не выявляются активные остатки комплексных пластид, которые подтверждали бы родство с динофитами.
Клеточная стенка часто имеет тонкие поры, которые выходят в трихоцисты, имеющие форму мешочков. При раздражении трихоцисты выбрасывают белковые нити. Клеточная стенка у многих динофитов построена очень характерным образом из многоугольных целлюлозных пластинок, которые образуют панцирь с поперечной и продольной бороздами. Чешуйки панциря (как и створки у диатомовых водорослей) закладываются в плоских полостях внутри плазмалеммы; поверх панциря сохраняется плазматическая мембрана.
На пересечении продольной и поперечной борозд отходят два жгутика, каждый из которых располагается в одной из борозд (см. рис. 11.70, А). Поперечный жгутик несет один ряд несколько более длинных волосков, продольный жгутик — два ряда более коротких волосков. Боковые волоски значительно тоньше, чем у Нeterokontophytа и Сryptophyta. Жгутик, колеблющийся в поперечной борозде, вызывает вращательное движение клетки вокруг продольной оси, тогда как жгутик, находящийся в продольной оси, обеспечивает движение клетки вперед. Например, клетка Рeridinium, перемещаясь по винтовой линии, за секунду преодолевает расстояние, в несколько раз большее своей длины.
Хромосомы у большинства динофитов (как и у эвгленовых) различимы и в рабочем ядре, так как в интерфазе они остаются в конденсированном состоянии (ср. другие эукариоты; см. рис. 2.27).
При электронной микроскопии хромосомы выглядят как компактно сложенные фибриллы («гирлянды»). Фибриллы имеют диаметр 2,5 нм, что соответствует диаметру двойной спирали ДНК (см. рис. 1.5). Хромосомы других эукариот, напротив, обладают субмикроскопическими фибриллами, имеющими диаметр, примерно в 10 раз больший — 25 — 30 нм (см. рис. 2.21). Эти более толстые хроматиновые спирали из двойной спирали ДНК с центральным нуклеогистоновым тяжем в хромосомах динофитов отсутствуют. В этом имеется некоторое сходство с нуклео- плазмой прокариотных бактерий и синезеленых водорослей.
Вегетативное размножение осуществляется путем косого деления. У имеющих панцирь форм (например, Сеratium) покров, как правило, разрывается под углом к поперечной борозде, и у новых клеток восполняются недостающие половины панциря (см. рис. 2.83, D). Однако у видов некоторых родов (например, Рeridinum) весь панцирь отбрасывается перед делением, так что каждая из возникших дочерних клеток полностью заново строит собственный панцирь. После нескольких таких делений внутри панциря развиваются две клетки, сначала голые, которые покидают материнский покров и покрываются новыми панцирями. При неблагоприятных условиях внутри панциря возникают толстостенные цисты, способные длительно сохранять жизнеспособность.
Половое размножение на данный момент установлено лишь у немногих динофитов. У Сеratium оно анизогамное, с зиготической сменой ядерных фаз (мейоз при прорастании зиготы); у Glenodinium описаны изогаметы, которые возникают в клетках (гаметангиях), выходят наружу и сливаются друг с другом.
Распространение. Большинство динофи- тов из примерно 1000 видов (120 родов) обитает в морях, где они совместно с диатомеями (отдел Нeterokontrophyta) составляют основную часть фитопланктона (важные первичные продуценты в морях). Наибольшее разнообразие форм достигается в более теплых водах, а наиболее массовое развитие — в более холодных водах. В пресных водах обитают лишь немногие Peridiniales, однако иногда в большом количестве; в высокогорных озерах они могут составлять до 50 % биомассы. Многие виды обладают характерными придатками, служащими для парения в воде (рис. 11.70, С, F — J). Noctiluca miliaris (гетеротрофный вид с голыми клетками!), а также видыCeratium, Gonyaulax и Peridinium вызывают свечение морской воды. Массовое развитие динофитов («красные приливы») могут вызывать гибель рыб1. Округлые эндосимбионты разных морских животных известны под общим названием «зооксантеллы» (рис. 11.69). Все кораллы, образующие рифы, живут в симбиозе с такими динофитами. Без эндосимбионтов кораллы выживают, однако теряют способность образовывать известковый скелет. Некоторые виды динофитов живут как паразиты на поверхности и внутри тела морских животных. Среди подобных гетеротрофных форм встречается фаготрофия («заглатывание» бактерий и планктонных водорослей).
1 От «красных приливов» страдают не только рыбы, но и многие беспозвоночные; употребление всех этих животных в пищу может вызывать отравление человека. — Примеч. пер.
Рис. 11.70. Dinophyta (Pyrrhophyceae): А — Peridinium tabulatum (600x); В — Gymnodinium aeruginosum (300х); C — Omithocercus splendidus (125x); D1, D2 — Ceratium hirundinella после деления (350x); E — Ceratium cornutum, циста (150x); F — Ceratium tripos (125x); G — C. palmatum (125x); H — C. reticulatum (65x); J — C. fusus (50x)
Систематика. Единственный класс Dinophyceae подразделяется на 4 порядка. Представители Dinophysiales (1) имеют стенку, две половинки которой (эпикон и гипокон) дополнительно подразделяются продольной бороздой; продольная и поперечная борозды часто окаймлены далеко выступающими пластинчатыми выростами (Ornithocercus, рис. 11.70, С). У Реridiniales (2) клетка или имеет простой покров (Gymnodiniaсеае, В), или одета целлюлозными пластинками (Реridiniасеае с родами Реridinium и Сеratium, А, D — J). У морских видов имеются мешковидные «пузулы», которые выходят узким каналом в борозду, где расположен жгутик; их функция еще не выяснена. Коккоидному уровню организации соответствует порядок Dinococcales (3), нитчатому — Dinotrichales (4), с немногочисленными родами в каждом из них.
В ископаемом состоянии динофиты известны с юры и представлены большим разнообразием форм; в мелу встречаются многочисленные превосходно сохранившиеся представители разных таксонов. Кроме того, так называемые гистрикосферы в отложениях от докембрия до голоцена были идентифицированы как принадлежащие динофитам репродуктивные клетки. Они играют важную роль как «руководящие» микроскопические ископаемые.
3. Приложение к Rhodobionta: отдел Нaptophyta
Этот отдел (называемый также Рrymnesiophyta) включает формы, представляющие монадный, пальмеллоидный, коккоидный и нитчатый уровни организации. Преобладают одноклеточные формы монадного типа. Большинство видов обитает в планктоне морей. Монадные клетки имеют два жгутика, чаще всего равной длины, которые несут не жесткие волоски, а субмикроскопические чешуйки или узелки из органического материала. Вдобавок к этим жгутикам каждая клетка имеет еще дополнительный нитевидный придаток — гаптонему. Она служит не для движения, а для прикрепления. Ее субмикроскопическая структура четко отличается от строения жгутиков. На поперечном срезе гаптонемы можно различить 6 или 7 серповидно расположенных трубочек (структуры 2 + 9 нет!). У некоторых форм эта гаптонема редуцирована до короткой культи. Поверхность клетки снаружи имеет чешуйки разного размера или узелки, которые образуются в пузырьках Гольджи и затем откладываются наружу. Цистерны Гольджи в середине пузыревидно расширены. Желтые, желто-бурые или бурые хлоролласты содержат хлорофиллы а и с, β-каротин и ксантофиллы; в качестве запасного вещества откладываются хризоламинарин, масло и парамилон. Хлоропласта окружены складкой эндоплазматической сети; опоясывающая ла- мелла (см. рис. 11.72, G) отсутствует. Тилакоиды группируются по три. Глазок, состоящий из шариков, располагается в хлоропласте вплотную к его мембране. Базальное расширение жгутика отсутствует.
Систематика. В этом отделе известно около 250 видов, относящихся примерно к 45 ролам. В пресных водах обнаружено лишь немного видов, Haptophyceae — единственный класс этого отдела.
1. Порядок: Prymnesiales. Представители этого порядка отличаются чаще всего очень длинной гаптонемой. Клетки покрыты двумя слоями полисахаридных чешуек, видимых только при электронной микроскопии (рис. 11.71, А). Чешуйки отличаются своей радиальной лучевой структурой; для чешуек наружного слоя характерен к тому же приподнимающийся край. Chrysochromulina имеет очень длинную гаптонему, которая примерно в 5 раз длиннее жгутиков. Наряду с автотрофным питанием возможно также фаготрофное, при котором, например, могут поглощаться целые клетки Chlorella. Клетки могут переходить в амебоидную фазу. Размножение осуществляется путем продольного деления имеющих жгутики или амебоидных клеток на несколько дочерних клеток. Виды рода Prymnesium имеют короткую, виды рода Chrysochromulina — более длинную гаптонему. Prymnesium parvumприкрепляется своей гаптонемой к жабрам рыб и при массовом развитии вызывает в прудах с соленой водой гибель рыб вследствие (как и Chrysochromulina polylepis) выделения токсина.
Рис. 11.71. Haptophyta: А — Prymnesiales, Chrysochromulina chiton, пластинки панциря (10000х); В, С — Coccolithophorales: В — Syracosphaeia pulchra, редуцированная гаптонема между жгутиками (1 500х); С — ископаемый кокколит (Deftandrius sp.), построенный из кальцитовых ромбоэдров; нижний мел (700х)
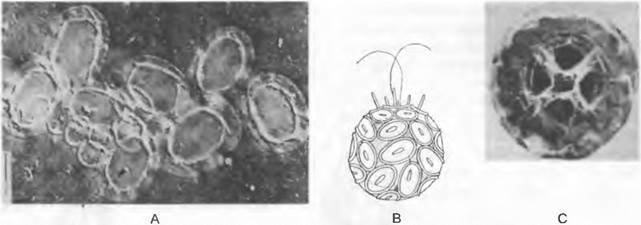
2. Порядок: Coccolithophorales. Гаптонема короткая или полностью отсутствует (см. рис. 11.71, В). Клетки несут поверх плазмалеммы два слоя тонких полисахаридных чешуек. Еще один внешний слой имеет самые разнообразные чешуйки, пластинки или палочки («кокколиты»). Последние также закладываются в пузырьках Гольджи сначала как целлюлозные пластинки, на которые потом накладываются удивительные отложения кальцита, специфичные для каждого вида и поэтому крайне разнообразные. Выдвигающиеся наружу кокколиты образуют правильный панцирь вокруг клетки. В этом порядке известны также виды, ставшие в результате утраты жгутиков неподвижными (CoccoUthus pelagicus) или же имеющие смену жгутиковой и прикрепленной стадий (Syracosphaera). У Hymenomonas carterae происходит гетероморфная смена поколений, причем более развитый по сравнению со спорофитом гаметофит имеет вид разветвленных нитей. Coccolithophorales (или же их кокколиты), известные в ископаемом состоянии с юры (рис. 11.71, С), объединяют характерные ископаемые виды микроорганизмов (128 родов). Они составляют важную долю в определенных известковых отложениях, где представлены в количестве до 800 млн кокколитов на 1 см3 породы (Применение — см. бокс 11.6).
4. Приложение к Rhodobionta: отдел Heterokontophyta (= Chrysophyta, Chromophyta)1
1 В настоящее время для этого отдела наиболее употребительно название охрофиты (Ochrophyta). — Примеч. пер.
Этот отдел при значительных различиях форм таллома отличается однообразием ультрамикроскопических структур (например, гетероконтными подвижными клетками).
Здесь представлены все морфологические ступени — от монадной до сифоновой организации; наивысший уровень организации Нeterokontophyta — морфологически расчлененные и анатомически дифференцированные паренхиматозные талломы.
Убедительно доказано родство автотрофных Неterokontophyta и гетеротрофных, грибоподобных Labyrinthulomycotа и Оomycota. Структурные особенности их клеток, несмотря на наличие пластид, приобретенных путем вторичного эндосимбиоза, настолько совпадают между собой, что Неtегокоnторhуtа и Ооmycota следовало бы объединять в общее подцарство Heterokontobionta (см. Ооmycota). Только структура пластид оправдывает рассмотрение Нeterokontrophyta в приложении к Rhodobionta, поскольку пластиды Нeterokontophyta и Rhodobionta имеют общее происхождение.
Пластиды. Иногда зеленые, однако чаще за счет наличия дополнительных пигментов желтые, желто-коричневые до коричневых, хлоропласты содержат хлорофиллы а и с, β-каротин и различные ксантофиллы (см. табл. 11.3). В дополнение к двум мембранам хлоропласта пластиды окружены еще складкой эндоплазматической сети (рис. 11.72, G).
Рис. 11.72. Xanthophyceae: А — С — Mischococcales, Capitulariella radians (500x): A — зооспора, В — отделившимся зооспорангий, С — таллом с расположенными на концах нитей зачатками зооспорангиев; D — Chloramoebales, Ankylonoton pyreniger в процессе деления (1000х); Е — G — Tribonematales: Е — фрагмент нити Tribonema с характерными Н-образными структурами клеточной стенки, попарно вдвигающимися одна в другую (600х), F — гетероконтная зооспора Tribonema (2 300х), G - хлоропласт Bumilieria (30000х); а — глазок; с — хлоропласт; е — оболочка из складки эндоплазматической сети; g — опоясывающая ламелла из трех периферических тилакоидов; m — две собственные мембраны хлоропласта; р — пульсирующая вакуоль; t — стопка (ламелла) из трех тилакоидов
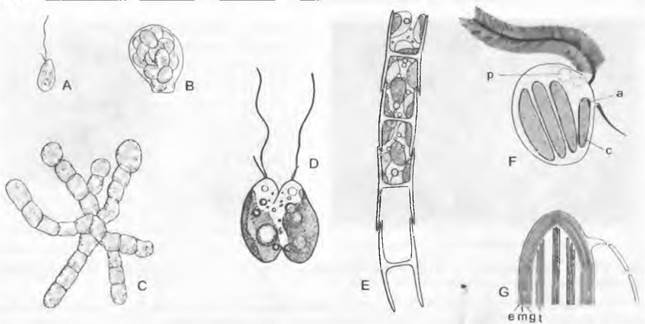
В хлоропластах тилакоиды сгруппированы по три в стопки (см. рис. 2.88) — расположение, наблюдаемое также, например, у Dinophyta и Euglenophyta. Непосредственно под оболочкой хлоропласта, параллельно ее внутренней поверхности, расположены тилакоиды, составляющие опоясывающую ламеллу, которая здесь очень харак терна (ср., например, представителей Нар- tophyta и Heterokontophyta, имеющих многие сходные вещества, с Chlorophyta). Глазки, если они присутствуют, располагаются вблизи основания жгутиков, но внутри хлоропласта.
В качестве запасных полисахаридов образуется хризоламинарин, а иногда также ламинарии и сахароспирт маннит. Они находятся вне хлоропластов, но в основном вблизи пиреноидов. Часто накапливается масло, которое появляется у пиреноидов, однако преимущественно оно находится вторично в вакуолях.
Клеточные стенки очень по-разному усилены дополнительными защитными слоями.
Жгутики. Клетки, имеющие жгутики, гетероконтные. Они несут один длинный, направленный вперед перистый жгутик и один направленный назад гладкий жгутик. Перистый жгутик имеет два ряда волосков, которые образуются в цистернах эндоплазматической сети. Гладкий жгутик (иногда редуцированный) часто имеет расширение у своего основания.
Отдел подразделяется на пять классов. В первых двух классах хлоропласты, цвет которых бывает от зеленого до желто-зеленого, не содержат фукоксантина.