БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
2. Класс: желтозеленые водоросли (Xanthophyceae)
Желтозеленые водоросли охватывают все уровни организации таллома — от амебоидного и монадного до сифонового. Зеленые хлоропласты окрашиваются НСI в синий цвет и содержат вместо фукоксантина (табл. 11.2) ксантофиллы — гетероксантин и вощериаксантин. Жгутики гетероконтного типа прикреплены сбоку. Желтозеленые водоросли, таким образом, несмотря на зеленую окраску, во многом сходны с остальными классами Heterokontophyta. От зеленых Chlorophyta этот таксон отличают отсутствие хлорофилла b и завершение заднего жгутика тонким волоском (как у бурых водорослей).
У некоторых форм клеточная стенка состоит из двух накладывающихся друг на друга половинок. Они в значительной мере состоят из целлюлозных микрофибрилл и часто пропитаны кремниевой кислотой (однако без панцирей из кремниевой кислоты!). Некоторые виды образуют эндогенные цисты с оболочкой, пропитанной кремниевой кислотой; цисты имеют форму коробки с крышкой.
Размножение. Большинство желтозеленых водорослей размножаются бесполым путем. Только в одном роде (Vaucheria) известно половое размножение в гаплонтном жизненном цикле (зиготическая смена ядерных фаз!); этот случай рассмотрен ниже1.
1 Согласно другим данным, жизненный цикл Vaucheria диплонтный (гаметическая смена ядерных фаз). — Примеч. пер.
При бесполом размноженииVaucheria концы ветвей вздуваются и отделяются от остального таллома перегородкой. Все многоядерное содержимое отделившейся клетки выходит, разрывая ее стенку, в виде одной яйцевидной крупной (размером около 1/10 мм) зооспоры (рис. 11.73, В). Ее поверхность покрыта многочисленными парными жгутиками несколько неравной длины, двигающимися синхронно. В бесцветной периферической части зооспоры позади каждой пары жгутиков расположены по два блефаропласта и одно грушевидно заостренное ядро; от них ближе к центру находятся хлоропласты (рис. 11.73, С); имеются пульсирующие вакуоли. Морфологически это образование соответствует совокупности всех зооспор, образующихся в одной клетке, и, таким образом, представляет собой синзооспору.
Половое размножение. Оогонии и спермогонии Vaucheria возникают на нитях таллома как боковые выросты (рис. 11.73, Е: о, s). Зачаток оогония (о) содержит сначала многочисленные ядра, которые, однако, все, кроме одного — будущего ядра яйцеклетки, вместе с частью хлоропластов мигрируют обратно в несущую нить; только тогда образуется поперечная перегородка. Оставшиеся хлоропласты, капельки масла и ядро отходят в нижнюю часть оогония. тогда как в клювовидном носике скапливается бесцветная плазма, которая при разрыве стенки оогония выступает в виде шарика. Многоядерный спермогоний (s) вместе с несущей его ветвью загнуты в виде рога. Его вершина при созревании ослизняется. Многочисленные крошечные сперматозоиды выходят наружу, внедряются в отверстие оогония и скапливаются перед бесцветным воспринимающим шариком яйцеклетки. Сперматозоиды (рис. 11. 73, F) гетероконтные.
После оплодотворения яйцеклетки одной из ♂ гамет богатая маслом цистозигота окружается многослойной стенкой, переходит в состояние покоя (гипнозигота) и прорастает позже после редукционного деления непосредственно в новую гаплоидную нить.
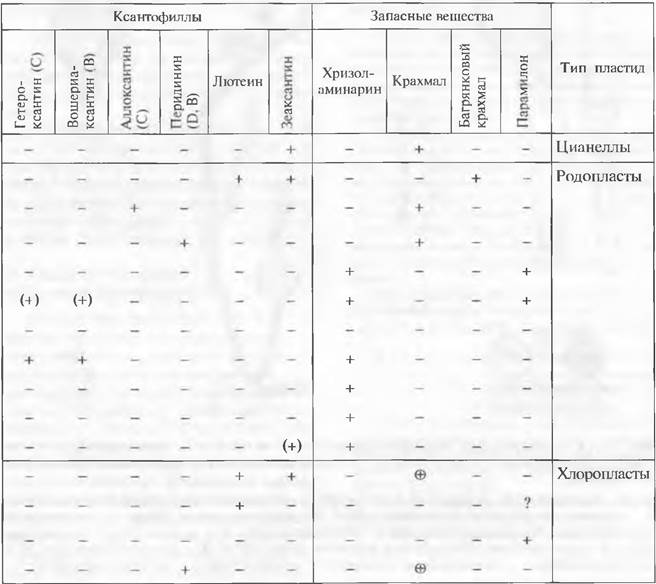
Таблица 11.2. Некоторые химические признаки классов водорослей
Отделы / классы |
Хлорофиллы |
Фикобилины |
Каротины |
Ксантрофиллы |
||||||
a |
b |
с |
α |
β |
Диадино- ксантин (С) |
Диато- ксантин (С) |
Фукоксантин (D, В, А) |
|||
Glaucophyta |
* |
+ |
- |
(•) |
+ |
- |
+ |
- |
- |
- |
Rhodophyta |
* |
+ |
- |
- |
+ |
(•) |
+ |
- |
- |
- |
Cryptophyta |
О |
+ |
- |
+ |
+ |
+ |
(•) |
- |
(+) |
- |
Dinophyta |
∆(O) |
+ |
- |
+ |
- |
- |
+ |
(+) |
(+) |
(•) |
Haptophyta |
∆ |
+ |
- |
+ |
- |
- |
+ |
(+) |
(+) |
+ |
Heterokontophyia |
∆ |
+ |
- |
+ |
- |
- |
+ |
+ |
(+) |
(+) |
Chloromonadophyceae |
∆ |
- |
+ |
- |
- |
+ |
+ |
(+) |
- |
|
Xanthophyceae |
∆ |
+ |
- |
+ |
- |
- |
+ |
+ |
+ |
- |
Chrysophyceae |
∆ |
+ |
- |
+ |
- |
- |
+ |
(+) |
(+) |
+ |
Bacillariophyceae |
∆ |
+ |
- |
+ |
- |
(•) |
f |
+ |
+ |
+ |
Phaeophyceae |
∆ |
+ |
- |
+ |
- |
- |
+ |
(•) |
(•) |
+ |
Chlorohyta |
* |
+ |
+ |
- |
- |
(•) |
+ |
- |
- |
- |
Chlororachniophyta |
О |
+ |
+ |
- |
- |
- |
- |
- |
- |
- |
Euglenophyta |
∆ |
+ |
+ |
- |
- |
- |
+ |
+ |
(+) |
- |
Streptophyta |
* |
+ |
+ |
- |
- |
- |
+ |
- |
- |
- |
Примечание. Условные обозначения — по van den Hoek, обзор ксантофиллов — по Metzner: * — с простыми пластидами (возникшими путем первичного эндоцитобиоза); О — с комплексными пластидами и нуклеоморфой (возникшими путем вторичного эндоцитобиоза); ∆ — с комплексными пластидами без нуклеоморфы (возникшими путем вторичного эндоцитобиоза); + — важный пигмент или запасной полисахарид; (+) — пигмент встречается; (•) — пигмент редок или только в незначительном количестве; пигмент или запасной полисахарид отсутствуют. В случае крахмала: + — вне хлоропласта; © — откладывается внутри хлоропласта; А— 8-кетокаротин, например, фукоксантин или сифоноксантин (последние только у Prasinophyceae и Bryopsidophyceae из зеленых водорослей);
В классе Хаrthophyceae описано примерно 400 видов из 40 родов, которые обитают в пресных водах, отчасти также в морях и на сырой почве.
Монадные формы объединяются в порядок Сhloramoebales (1-й порядок — например, Аnkylonoton, рис. 11.72, D). Капсальный уровень организации представлен в порядке Heterogloeales (2-й порядок); к нему относится морской род Heyerocapsa. Порядок Мischococcales (3-й порядок) объединяет парящие в воде или сидячие формы с жесткой клеточной оболочкой. У разветвленных, обитающих в наземных условиях видов Сарitulariella (рис. 11.73, С) спороцисты отрываются целиком и только потом высвобождают зооспоры (аналогично Тrentepohlia, см. рис. 11.93, С, D). Тribonematales (4-й порядок) представлены в пресной воде и на влажной почве часто встречающимися видами рода Тribonema (рис. 11.72, Е), стенки неразветвленных нитей которых в продольном сечении построены из Н-образных элементов.
В — аллен-каротин, например, вошериаксантин и неоксантин (последний у Euglenophyta, Chlorophyta, Ensligmatophyta, у некоторых Eleterokontophyta, Rhodophyta); С — алкинкаротиноиды; D — каротиноидные эфиры, т. е. ксантофиллы, которые на одной или обеих гидроксильных группах несут остатки жирных кислот. В таблице не учитываются 4-кето-каротины. например эхиненон у Euglenophyta + и Chlorophyta +, у Heterokontophyta (+). Цианеллы — фотосинтезирующие органеллы с пептидоглюкано- вой стенкой. Родопласты — пластиды Rhodophyta и производные от них фотосинтезирущие органеллы. Хлоропласты — пластиды Chlorophyta и производные от них фотосинтезирующие органеллы.
К Неterosiphonales (5-й порядок) относится пузыревидная, растущая на влажном иле водоросль Воrydium (рис. 11.73, в), клетка которой размером около 2 мм закрепляется в иле ризоидами. Пузырь содержит в постенной плазме многочисленные ядра и дисковидные хлоропласты, стенка которых состоит из пектиноподобной фракции и целлюлозы. Когда Воtrydium покрыт водой, то образуются многочисленные гетероконтные зооспоры, которые после разрушения стенки пузыря выходят наружу и на подходящем субстрате вырастают в новый пузырь. При высыхании возникает большое количество многоядерных цист из протопластов, отходящих в ризоид.
Рис. 11.73. Xanthophyceae, Heterosiphonales; А — F — Vaucheria (А — С — V. repens, D, Е — V. Sessilis (F — V. synandra): А — закладка спорангия (150х), В — выходящая из спорангия синзооспора (150х), С — край синзооспоры (500х), D — возникающее из синзооспоры В растение с ризоидом и гаметангиями (70х), Е — участок нити с гаметангиями (150х), F — сперматозоид (700х); G, Н — Botrydium, В. granulatunr. G — целое растение (30х), Н — зооспора (1000х); с — хлоропласты; о — оогоний; s — спермогоний; z — зачаток синзооспоры
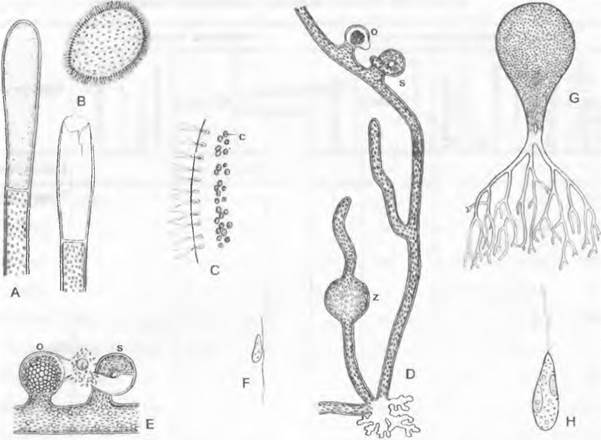
Сифональное строение имеют также широко распространенные виды рода Vaucheria. Они обитают в пресной воде или на влажной почве, прикрепляются к субстрату пучком ризоидов и имеют состоящий из лишенных поперечных перегородок разветвленный таллом, представляющий собой как бы систему нитей (рис. 11.73, D) с многочисленными ядрами и пластидами. Клеточная стенка содержит пектиноподобную массу и целлюлозу.
Стенки некоторых видов инкрустированы известью и могут образовывать известковые туфы.
К желтозеленым водорослям примыкает небольшая группа (рассматриваемая также как самостоятельный отдел Eustigmatophyta, которая отличается от Xanthophyceae рядом ультраструктурных признаков; хлоропласты без периферической опоясывающей ламеллы; пиреноиды только в хлоропластах вегетативных клеток; глазок на переднем конце клетки вне хлоропласта1. В жизненном цикле капсальных и коккоидных организмов могут иметься гетероконтные жгутиковые клетки. У Сhlorobotrys несколько клеток образуют колонию в слизистой обвертке; некоторые виды этого рода широко распространены в мочажинах болот.
1 Эта группа, чаще всего трактуемая как класс, отличается от Хanthophyсеае тем, что у большинства представителей фактически только один направленный вперед перистый жгутик; вместо второго жгутика имеется лишь базальное тело. Только у двух представителей задний гладкий жгутик выступает наружу. Кроме того, из пигментов преобладает виолаксантин, а не диатоксантин. — Примеч. пер.
Для рассматриваемых ниже классов (3 — 5) характерен фукоксантин в качестве дополнительного пигмента в хлоропластах, имеющих окраску от желтой до бурой.
Рассматриваемые далее классы 3 и 4 включают преимущественно одноклеточные или колониальные водоросли и реже — формы, образующие неразветвленные нити. Встречающиеся как исключение тканевые талломы имеют микроскопически малые размеры. Смена поколений неизвестна.