БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
5. Класс: бурые водоросли (Phaeophyceae)
Бурые водоросли — группа, представленная самыми разнообразными формами (рис. 11.80, А; см. рис. 5.5; 11.83; 11.85). Это могут быть и маленькие разветвленные нити, разделенные на клетки (гетеротрихальный тип), и псевдопаренхиматозные талломы, и многометровые растения с сильно выраженной дифференциацией на органы и. ткани (тканевые талломы! — см. 5.3.2). Макрофиты из бурых водорослей часто подразделены на органы (филлоид, каулоид, ризоид), которые напоминают лист, стебель и корень высшего растения. Одноклеточные формы отсутствуют, т. е. монадные и коккоидные уровни организации здесь не представлены. Наряду с красными и некоторыми зелеными (Chara)водорослями бурые водоросли относят к наиболее высокоорганизованным формам.
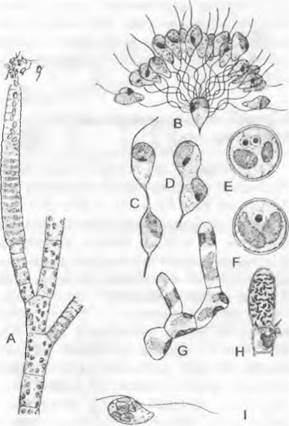
Рис. 11.80. Phaeophyceae, Ectocarpales: А — D — Ectocarpus siliculosus: А — ветвь гаметофита с многогнездным гаметангием (380х), В — D — оплодотворение (В — 1 200х; С, D — 1 600х), Е, F — Asperococcus bullosus, зигота и слияние ядер (2000х); G — Nemacystis divarica- tus, прорастание (780х); Н — J — Ectocarpus: Н — Ectocarpus lucifugus, одногнездный мейоспорангий на диплоидном спорофите (400х), J — Ectocarpus globifer, зооспора, волоски перистого жгутика не изображены
Пластиды. Бурые хроматофоры содержат кроме характерных для всего отдела ассимиляционных пигментов прежде всего фукоксантин в качестве дополнительного пигмента, окраска которого перекрывает окраску всех других компонентов.
Клеточная стенка состоит из твердой и слизистой фракций; первая образована целлюлозными фибриллами и альгинатом, последняя — альгинатом и фукоиданом. Альгинаты — это соли альгиновой кислоты (полимера двух сахарокислот: β-D-маннуроновой и β-L-гулуроновой) с различными катионами (такими как ионы Са2+, Mg2+, Na+). Подвижные стадии (зооспоры и гаметы) имеют форму от грушевидной до веретеновидной и несут чаще всего 2 жгутика неравной длины (рис. 11.80, А, В, I)1.
1Важной особенностью подвижных стадий бурых водорослей считается боковое прикрепление жгутиков. — Примеч.пер.
Вблизи жгутиков находится красно-коричневый глазок в буром хлоропласте (один, изредка несколько). Волоски перистого жгутика закладываются в пузырьках эндоплазматической сети или пузыревидных частях ядерной эндоплазматической сети. Гладкий жгутик в базальной части вздутый; это вздутие расположено вблизи глазка и, возможно, действует как фоторецептор. Жгутики заканчиваются (гладкий всегда, а перистый иногда) тонким волосовидным придатком. Эта особенность кроме бурых и желтозеленых водорослей больше нигде не встречается.
Жизненный циклосуществляется в виде смены поколений, причем мейоспоры всегда образуются в одногнездных спороцистах, а гаметы, — как правило, в многогнездных (= многокамерных) гаметангиях. Гетерофазная смена поколений изоморфная, гетероморфная — до крайне гетероморфной, с (почти) полной редукцией гаплоидного гаметофита. Увеличение значения в жизненном цикле диплоидного спорофита, появившееся среди Ectocarpales, рассматривается как прогрессивная тенденция.
Встречаемость и образ жизни. Большинство из примерно 1500 — 2000 видов бурых водорослей, распределяющихся по 250 родам, обитает в морях, причем наиболее интенсивного развития они достигают в водах умеренных и высоких широт. Это бентосные водоросли (см. бокс 11.7): они живут как литофиты. прочно прирастая к скалам, камням, бревнам и т. д. Некоторые из них при низкой воде остаются на воздухе. Часто бурые водоросли обитают как эпифиты на других водорослях. Они образуют в литоральной зоне скалистых побережий пышные заросли с характерным зональным распределением видов (см. рис. 11.102). Внушительное зрелище представляют собой на тихоокеанском побережье Америки подводные леса, которые образованы бурыми водорослями из родов Lessonia. Macrocystis и Nereocystis с многометровыми талломами. Крошечные нитевидные или дисковидные бурые водоросли хотя и менее заметны, но тоже широко распространены, в том числе на каменистом субстрате, морских желудях, улитках, двустворчатых моллюсках и эпифитно на более крупных бурых водорослях. Мелкие бурые водоросли могут до определенной степени жить эндофитно в более крупных водорослях. В пресных водах встречается лишь немного видов, относящихся примерно к пяти родам.
Класс подразделяется на 11 порядков, из которых следующие, имеющие меньшее значение, дальше рассматриваться не будут: Chordariales (с родами Chordaria и Leptonema; Elachista — эпифит на Fucus с вегетативной диплоидизацией в гаметофитах); Desmarestiales (характеризуются объединением нитей в псевдопаренхиматозную кору; гетероморфная смена поколений), Dictyosiphonales (паренхиматозный таллом), Scytosiphonales (псевдопаренхиматозные корковидные микроталломы (спорофиты) сменяются в жизненном цикле паренхиматозными мегаталломами — гаметофиты?), Sporochnales и Sphacelariales (например, Наlopteris от Ectocarpales и др. отличается наличием верхушечной клетки).
1. Порядок: Есtocarpales. К порядку относится большинство бурых водорослей. Очень распространены виды рода Еctocarpus. Их кустовидные талломы, состоящие из разветвленных нитей, по облику напоминают зеленые водоросли из рода Сladophora (см. рис. 11.90), однако окраска их бурая. Это обитатели мелководий наших морей, которые ползучими нитями прикрепляются к субстрату (каменистому субстрату, более крупным водорослям). Нити растут интеркалярно, верхушечная клетка отсутствует, только часть клеток способна преобразовываться в органы размножения. Жизненный цикл в целом представляет собой изоморфную (или слабогетероморфную) смену поколений.
Гаметофит. Гаплоидный, кустовидно разветвленный нитчатый таллом гаметофита несет по бокам и на концах нитей многогнездные гаметангии, в которых вовсе не каждая клетка способна образовывать гамету. Гаметы освобождаются при разрушении внешних стенок гаметангия, выходя на его вершине. Несмотря на то что морфологически гаметы одинаковы, у многих видов рода Еаctocarpus имеется физиологическая анизогамия: женские («-») гаметы вскоре после выхода из гаметангия останавливаются и отбрасывают жгутики, тогда как мужские («+») гаметы, которые привлекаются феромоном эктокарпеном, группируются возле них. Концами своих более длинных жгутиков мужские гаметы прикрепляются к покоящимся женским, и одна из них сливается с ней (рис. 11.80, В).
Спорофит. После оплодотворения зигота без периода покоя вырастает в диплоидный спорофит, состоящий из нескольких более жестких и менее разветвленных нитей. На нем возникают многочисленные яйцевидные одногнездные спорангии, в которых после мейоза образуется большое количество мейозооспор, дающих начало новому гаметофитному поколению. Определение пола гаплогенотипическое.
Эта обычная — изоморфная и гетерофазная — смена поколений может усложняться многочисленными отклонениями, как правило, трудно объяснимыми: сменяющиеся поколения не всегда связаны с определенной ядерной фазой. Каждая генерация может непосредственно снова воспроизводить себя. Пол четко не фиксирован; организмы могут обходиться и без него.
Если вместилища репродуктивных клеток у Ectocarpus возникают на концах боковых ветвей, то у Pylaiella они дифференцируются интеркалярно.
У некоторых эпифитных видов родов Ectocarpus и Pylaiella гаметофиты и спорофиты встречаются не на тех же самых, а на разных растениях-форофитах (например, у Pylaiella litoralis— спорофит на Fucus, а гаметофит на Ascophyllum).
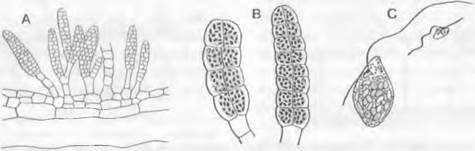
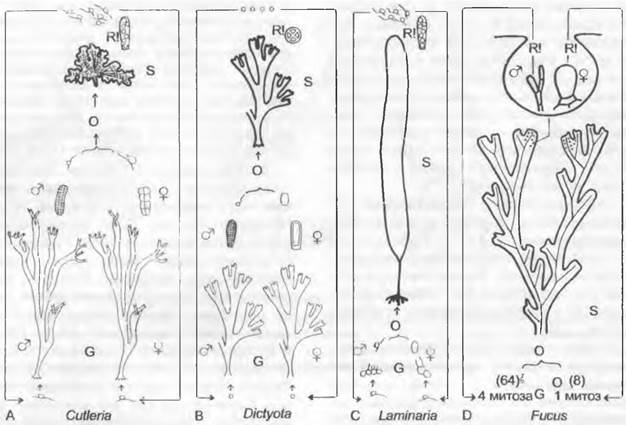
2. Порядок: Cutleriales. Смена поколений у Cutleria гетероморфная, с явно преобладающим гаметофитым поколением (рис. 11.81, А). Гаметофит — прямостоячее, вильчато разветвленное растение, состоящее из лентовидных лопастей, рассеченных на концах. У Cutleria multifida — водоросли из более теплых европейских морей — гаметофит обитает вблизи поверхности воды, имеет размер порядка 40 см и образует на мужских и женских растениях в микро- и мегагаметангиях мелкие (♂) и более крупные (♀) гаметы, снабженные жгутиками (рис. 11.81). Мужские гаметы привлекаются женскими гаметами посредством феромонов мультифидов, после чего происходит копуляция (анизогамия). Ранее описывавшийся как представитель отдельного рода (Aglaozonoia) спорофит заметно меньше (его размер — несколько сантиметров); он плоский, рассеченный на лопасти, распростертый, корковидный; обитает на скалах и раковинах моллюсков на глубине от 8 до 10 м. На верхней стороне паренхиматозного таллома находятся сорусы, состоящие из одногнездных спороцист. После мейоза они высвобождают зооспоры. Zanardinia имеет изоморфную смену поколений.
Рис. 11.81. Phaeophyceae, Cutleriales, Cutleria multifida: А — ♂, В — ♀ многогнездные гаметангии (400х); С — ♂ и ♀ гаметы, волоски перистых жгутиков не показаны (1 200х)
3. Порядок: Dictyotales. Плоские паренхиматозные талломы Dictyota размером с кисть руки многократно дихотомически разветвлены (рис. 11.82). В основе роста и вильчатого ветвления лежит деление крупной верхушечной клетки (см. рис. 5.12, В), которая отчленяет вниз от себя базальные сегменты. Последние делятся дальше на множество клеток, которые образуют ткани (см. рис. 5.12, В — D). Они дифференцируются на периферические ассимиляционные и центральные запасающие клетки (см. рис. 11.82). Время от времени исходная верхушечная клетка разделяется перегородкой, идущей вдоль оси таллома, на две; расположенные рядом клетки, продолжая рост, вызывают дихотомическое ветвление таллома. Смена поколений изоморфная (см. рис. 11 87, В).
Рис. 11.82. Рhaeophусеае, Dictyotales, Dictyota dichotoma: А — поперечный разрез через ♂ таллом с группой спермогониев (окруженной обверткой из покровных стерильных клеток, 200х); В — поперечный разрез ♀ таллома с группой оогониев (200х); С — яйцеклетка с тремя сперматозоидами (400х); D — поперечный разрез таллома с тетраспорангиями (один из них освободился) и «волосками» (200х)
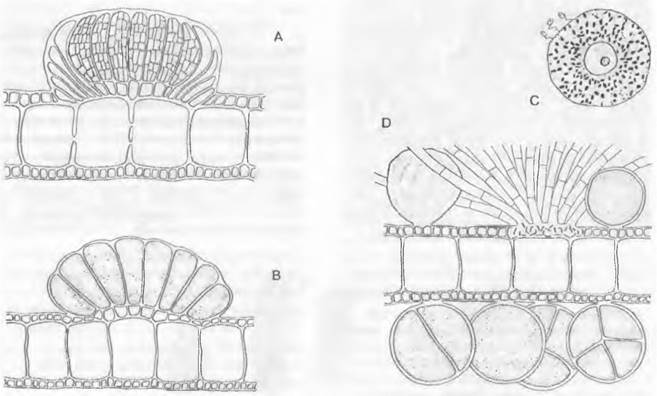
Гаметофит. Половое размножение — оогамия. Многогнездные спермогонии и оогонии расположены на разных растениях и всегда собраны в группы (сорусы) (см. рис. 11.82, А, В).
Каждый оогонии содержит крупную неподвижную коричневую яйцеклетку, которая выталкивается в воду и оплодотворяется сперматозоидом (С). Грушевидные сГ гаметы имеют один сильноредуцированный хлоропласт и только один боковой перистый жгутик; второй, редуцированный, жгутик с собственным базальным тельцем остается внутри клетки в виде крошечной культи. Гаметангии развиваются только в летние месяцы; выход сперматозоидов находится в зависимости от положения Солнца и Луны и происходит только 2 дня в месяц, каждый раз в первый час после рассвета.
Спорофит. Гаплоидные гаметофиты по своему внешнему облику полностью совпадают с диплоидным спорофитом (рис. 11.87, В). Мейоспоры, возникающие по 4 в одногнездных тетраспороцистах спорофитов (рис. 11.82, D), имеют относительно большой размер и лишены жгутиков. Между тетраспороцистами выступают бесцветные волоски.
Часто встречающаяся в более теплых морях вееровидная Раdina растет посредством коровой меристемы. Dictyopteris — за счет группы специальных верхушечных клеток.
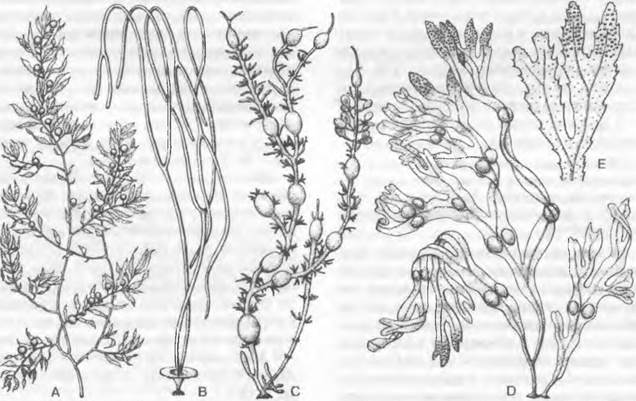
4. Порядок: Laminariles. Смена поколений гетероморфная, с явным преобладанием диплоидного спорофита (см. рис. 11.87, С). Спорофиты морфологически и гистологически очень дифференцированы и достигают иногда значительных размеров (рис. 11.83).
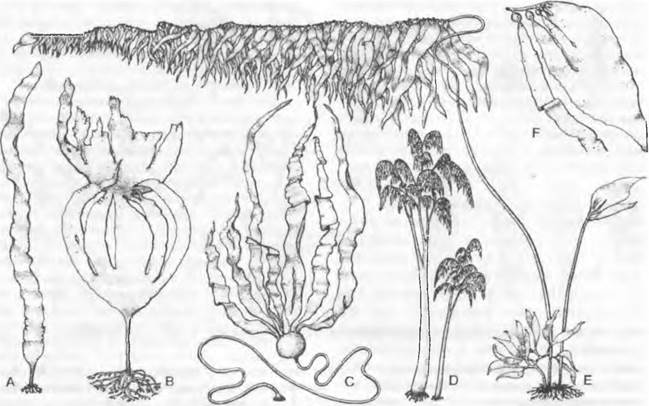
Рис. 11.83. Phaeophyceae, Laminariales: А — Laminaria saccharina (1/40х); В — Laminaria hyperborea, вверху с остатками прошлогодней пластины (1/40х); С — Nereocystis luetkeana(1/200х); D — Lessonia flavicans (1/30x); E — Macrocystis pyrifera (1/250x); F — тот же организм, вершина таллома (1/20х)
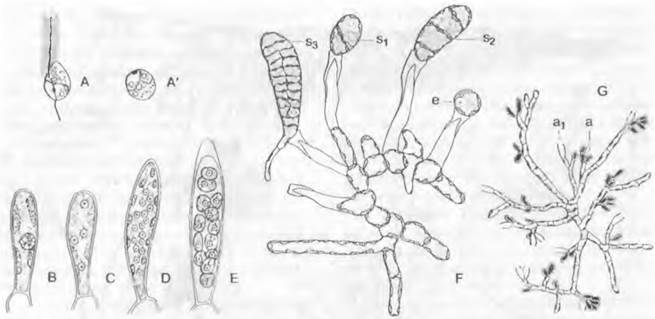
Гаметофиты всех Laminariales, напротив, имеют микроскопически малые размеры. Формы ♂ и ♀ хорошо различаются по строению, т. е. имеются четкие вторичные половые признаки. Мужские гаметофиты относительно сильно разветвлены, быстро растут, состоят из многих, но мелких клеток (рис. 11.84, G) и несут на концах веточек одноклеточные спермогонии с единственным сперматозоидом в каждом. Женские гаметофиты (F) имеют значительно более крупные клетки, но растут медленнее, и клеток у них меньше; в крайнем варианте они состоят только из одной-единственной мешковидной клетки и образуют оогонии с одной яйцеклеткой в каждом. Голая яйцеклетка выходит через отверстие на вершине оогония, где она чаще всего и остается (F: е), а после оплодотворения (оогамии) вырастает в диплоидный спорофит (F: s1— s3).
Рис. 11.84. Phaeophyceae, Laminariales: А — Е — Chorda filum: А — мейозооспоры, (А') округлившиеся перед прорастанием (1 200х); В — Е — развитие одногнездного спорангия (1000х): В — одноядерный, С — 4-ядерный, D — 16-ядерный, Е — с почти зрелыми зооспорами; F, G — Laminaria (300х): F — ♀ гаметофит, G — ♂ гаметофит; а — спермогонии (а1 — освободившийся); е — яйцеклетка; s1—s3 — молодые спорофиты, еще сидящие на освободившемся оогонии
Спорофитное поколение представляет собой макроскопическую фазу в жизненном цикле. На поверхности спорофита образуется кроме вытянутых стерильных клеток (парафиз) обширный слой булавовидных одногнездных спороцист (рис. 11.84, D). В каждой из них после редукционного деления и одновременно генотипического определения пола развивается большое число двухжгутиковых зооспор.
Спорофиты Мacrocysis pyrifera (см. рис. 11.83, Е) в холодных морях Южного полушария достигают в длину свыше 50 м; их таллом, закрепляющийся на глубине 2 — 25 м с помощью когтевидной присоски, несет по одной стороне своих осей (каулоидов, см. 5.3.2) длинные свисающие лопасти (филлоиды, см. 5.3.2), каждая из которых имеет в основании большой воздушный пузырь. Эти пузыри поддерживают водоросль на плаву у поверхности моря. Антарктические виды Lessonia (D), которые имеют стволовидные основные оси толщиной с человеческое бедро и длиной до 5 м, развивают на боковых ветвях длинные свисающие филлодии, внешне напоминают
пальмы. У Nereocystis (побережье Тихого океана от Калифорнии до Аляски) длинный (до 25 м) каулоид, напоминающий канат, несет на конце крупный воздушный пузырь (с высоким содержанием монооксида углерода!), от которого отходит пучок филлоидов (С). Chorda filum («морская струна») имеет шнуровидный неразветвленный таллом, достигающий нескольких метров в длину. Alaria кроме филлоида на вершине имеет еще боковые «листочки» меньшего размера1.
1 Интересно разделение функций этих филлоидов: крупный центральный филлоид с хорошо выраженной срединной жилой стерилен, а сорусы зооспорангиев развиваются только на мелких боковых филлоидах, называемых поэтому также спорофиллами. — Примеч. пер.
Виды Laminaria (ср. рис. 11.102) распространены по побережьям Северной Атлантики и образуют целые подводные заросли («луга») в непосредственной близости от уровня низкой воды. Их талломы длиной до 5 м состоят из многолетней «ножки» с когтевидным ризоидом (см. рис. 11.83. В) и листовидной лопасти (А), сложенной очень многими слоями клеток. Эта лопасть (филлоид) каждый год заменяется за счет того, что располагающаяся у ее основания интеркалярная зона роста лает начало новой «листовой пластинке»; старая пластина при этом отодвигается все дальше и постепенно отмирает (В). Филлоид у L. sacchanna (А) простой, у L. digitata и близких по облику видов (В) — дланевидно-рассеченный. Спорофиты L. hyperborea могут жить 10 — 20 лет.
Дифференциация тканей. На поперечном разрезе через каулоид Laminariales в месте перехода от периферии к центру видна сильная дифференциация. Снаружи видна меристодерма (покровная ткань). Ее клетки, способные делиться в нескольких направлениях, образуют тангенталь- ные, радиальные и горизонтальные стенки. Более глубокие слои меристодермы ответственны прежде всего за рост в толщину. Рост происходит с периодичностью в зависимости от времени года, так что на более старых каулоидах имеются хорошо выраженные годичные кольца. Клетки коры от периферии к центру становятся все крупнее. За счет ослизнения клеточных стенок возникают идущие отчасти продольно, отчасти радиально рыхлые ряды клеток, а в более старых каулоидах — слизистые ходы с широкими просветами. Коровый слой обеспечивает механическую прочность каулоида и в своих наружных частях, состоящих из мелких клеток с хлоропластами, функционирует как ассимиляционная ткань; отчасти и здесь осуществляется рост в толщину. Сердцевина служит для накопления и проведения питательных веществ. Она состоит из многоклеточных нитей (так называемых гиф), которые у поперечных перегородок расширены наподобие раструба. У видов некоторых родов (например, Nereocystis и Macrocysris) поперечные стенки таких нитей перфорированы в виде сита. С помощью радиоактивно меченных соединений углерода удалось установить транспортную функцию этих элементов. Подобные ситовидные трубки сходны по строению и функциям с уже обсуждавшимися ситовидными трубками высших растений (см. рис. 5.10).
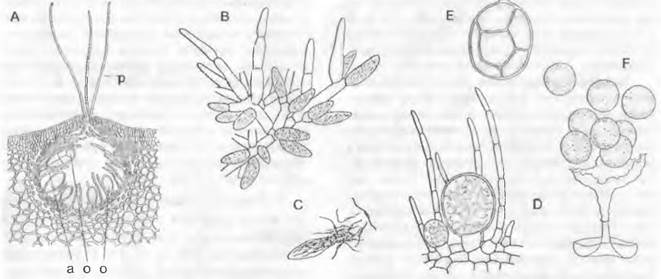
5. Порядок: Fucales. Эти водоросли могут из-за крайней редукции гаметофита рассматриваться практически как чистые диплонты (см. рис. 11.87, D). Размножение осуществляется путем оогамии. Смена ядерных фаз гаметическая, т. е. мейоз происходит при образовании гамет. Диплоидный спорофит (рис. 11.85) образует единственное имеющееся в жизненном цикле вегетативное тело в форме таллома, иногда достигающего в длину 1 м и более. У живущих несколько лет видов Fucus кожистые, лентовидные, дихотомически разветвленные талломы укреплены своего рода «срединной жилкой». Они прикрепляются присоской к каменистому субстрату. Концы ветвей таллома (верхушечная клетка — см. рис. 5.12, В) у некоторых видов Fucus несколько вздуты и несут тесно расположенные углубления в виде кувшинчиков — так называемые концептакулы1 (рис. 11.86, А), в которых между стерильными волосками (парафизами) расположены вместилища ♂и ♀ половых клеток (соответственно спермогонии и оогонии).
1 В русскоязычной литературе широко встречается также другое название концептакулов — скафидии. — Примеч. пер.
Рис. 11.85. Phaeophyceae, Fucales: А — Sargassum bacciferurrr, В — Himanthalia torea; С — Ascophyllum nodosum; D — Fucus vesiculosus; E — Fucus serratos, вершина таллома (A — E — 1/4x)
У некоторых видов спермогонии и оогонии расположены в одних и тех же кониептакулах (однодомность, например, у Fucus spiralis, рис. 11.86, А); другие виды двудомны (например, F. serratus и F. vesiculosus).
Рис. 11.86. Phaeophyceae, Fucales: А — Fucus spiralis, двуполый (однодомный) концептакул с оогониями (о) различного возраста (25х); В — F — Fucus vesiculosus: В — ветвь со спермогониями (а) (200х), С — выход сперматозоидов из спермогония (250х), D — молодые оогонии, Е — содержимое оогония после выхода его из стенки разделяется на восемь яйцеклеток, F — освобождение яйцеклеток (D — F — 120х); р — парафизы
Части таллома с концептакулами каждый год сбрасываются. Во вместилищах репродуктивных клеток происходит разное число митозов. Эти вместилища могут рассматриваться как одногнездные, дифференцированные по полу мейоспороцисты, а первичные продукты мейоза — как мейоспоры. Клетки, образующиеся в этих одногнездных вместилищах в результате митотических делений ядер, представляют собой в определенной степени крайне редуцированные гаметофиты, лишенные уже всякой самостоятельности; они, по-видимому, полностью интегрируются в соответствующие (оогонии или спермогонии) мейоспороцисты (вместилище репродуктивных клеток = мейоспороциста = гаметангий). Из 4 возникших после мейоза гаплоидных клеток в оогонии после первого мейоза возникает 8 яйцеклеток, а в спермогонии после четырех митозов — 64 сперматозоида.
Оогонии (рис. 11.86, А: о, D) представляют собой крупные округлые образования, сидящие на одноклеточной ножке. Стенка оогония состоит из трех слоев. При созревании разрывается сначала только наружный слой стенки, так что 8 яйцеклеток остаются окруженными двумя внутренними слоями, когда они покидают концептакул (Е). В морской воде, наконец, разрывается и самый внутренний слой стенки, так что 8 яйцеклеток (♀) начинают свободно парить в толще воды независимо друг от друга (♂).
Спермогонии — овальные клетки, тесно прижатые к обильно разветвленным коротким нитям (рис. 11.86, А: а; В). Стенка спермогония состоит из двух слоев. Внутрен-
няя стенка сохраняется и охватывает 64 сперматозоида (♂), когда весь пакет при созревании выталкивается из концептакула за счет выделения слизи. Сперматозоиды состоят преимущественно из ядерного вещества и одного-единственного рудиментарного хлоропласта, содержащего глазок; они снабжены двумя жгутиками (в противоположность прочим бурым водорослям направленный вперед перистый жгутик более короткий).
Освободившиеся сперматозоиды (С) привлекаются к яйцеклеткам феромоном фукосерратеном (ср. 8.2.1.1). Первоначально голая зигота окружается содержащей целлюлозу стенкой, прикрепляется к субстрату и, претерпевая клеточные деления, вырастает в новый диплоидный спорофит (см. рис. 7.23).
Fucales — вершина эволюционного ряда, характеризующегося прогрессирующей редукцией гаметофита. Это четко видно при рассмотрении особенностей некоторых Laminariales. Мужской гаметофит может быть редуцирован у них до одной-единственной клетки, в то время как содержимое переходящей к покою мейозооспоры выходит и непосредственно превращается в яйцеклетку. Диплоидное растение Fucus может, таким образом, трактоваться как спорофит, мейоспоры которого непосредственно становятся почти полностью исчезающим гаметофитом.
У многих видов Fucales (как и у большинства более крупных Laminariales) таллом находится в воде также вертикальном положении благодаря воздушным пузырям; они позволяют ему раскачиваться на волнах, не перемещаясь по дну. Виды Fucus образуют в морях Северной Европы низкорослые заросли на мелководье; при низкой воде они временно остаются на воздухе, но защищены выделяемой растениями слизью (фукоидином) и поэтому даже сохраняют способность к фотосинтезу.
Fucus serratus имеет таллом с зубчатыми краями; на талломе F. uesiculosus находятся округлые воздушные пузыри, наполненные газом. Воздушные пузыри имеет и встречающийся в тех же местах Ascophyllum nodosum (см рис. 11.85, С). У Himanthalia (В) таллом состоит из основания, напоминающего по форме юлу; на нем расположены от одной до нескольких ремневидных, вильчато разветвленных вертикальных ветвей. Более обильное ветвление таллома наблюдает
ся у распространенных в теплых морях представителей (250 видов!) рода Sargassum. Некоторые его виды благодаря наличию воздушных пузырей свободно плавают бесчисленными пучками в Саргассовом море Атлантического океана (от Гольфстрима и Вест-Индии до Азорских островов). Они размножаются здесь только вегетативно путем распада талломов (А). Сюда же относятся роды Cystoseira и Halidrys; Pelvetia (см. рис. 11.107): Coccophora и Durvillea (бокс 11.7, с. 199).
У бурых водорослей можно проследить прогрессивный переход от изогамии к оогамии через анизогамию. У исходных форм (Ectocatpus, см. рис. 11.80, А) гаметангии обоих полов многоклеточные, одинакового внешнего вида. У более высокоорганизованных форм (например, Cutlería) с увеличением размера ♀ гамет происходит также увеличение размера гаметангиев с одновременным уменьшением числа гнезд. У Dictyota и Laminaria это приводит к образованию только одной яйцеклетки. Если у Dictyota спермогонии еще многоклеточные, то у Laminaria они уже, как и оогонии, одноклеточные, с одним сперматозоидом. Однако такое развитие признаков не позволяет делать какие-либо выводы о фактической филогении. Смена поколений (рис. 11.87) изоморфная (Isogeneratae) или гетероморфная(Heterogeneratae), причем сходство поколений рассматривается как исходный вариант, тогда как преобладание спорофита (впервые отмеченное у Ectocarpales) — как более подвинутый вариант.
Филогения. Бурые водоросли, по-видимому, произошли с золотистыми от общих предков и уже рано стали эволюционировать разными линиями. Несмотря на свои иногда очень значительные размеры, бурые водоросли в целом сохранились в ископаемом состоянии хуже, чем известковые формы зеленых водорослей. Вполне вероятно, что они существовали уже в силуре и девоне. Некоторые нижнедевонские и силурийские «стволы» (Nematophycus = Prototaxites) толщиной с человеческое бедро, которые состояли из сплетения трубковидных нитей и заканчивались мощными пучками лопастей, напоминающих пластины Laminaria, по-видимому, тоже были бурыми водорослями.
Рис. 11.87. Phaeophyceae. Смена поколений и ядерных фаз у некоторых бурых водорослей (А — D). Гаплоидная фаза показана светлыми линиями, диплоидная — темными; G — гаметофит; S — спорофит; О — зигота; R! — редукционное деление
Обзор Heterokontophyta. В общем виде Heterokontophyta отражается все разнообразие форм организации, имеющихся у водорослей. В этом смысле данный отдел выглядит очень гетерогенным. С другой стороны, очевидны родственные связи различных форм через ряд общих «консервативных» признаков, которые скорее всего либо не играют селективной роли и стабильны, либо абсолютно необходимы для дальнейшего существования вида. На основе структуры подвижных клеток и хлоропластов, а также состава хлорофиллов, дополнительных пигментов и запасных веществ можно охарактеризовать Неterokontophyta как эволюционно единую группу водорослей.
Имея паренхиматозные талломы, бурые водоросли относятся к наиболее высокоразвитым морским растениям. По расчленению вегетативного тела и проводящим элементам, аналогичным ситовидным трубкам, они напоминают строение высших растений. Усложнение строения вегетативного тела в связи с приспособлением к многообразию условии внешней среды — эволюционный принцип, реализуемый в самых разных группах растений. При этом постоянно можно констатировать; одна-единственная эволюционная линия (группа родственных форм) развивается по пути многообразной дифференциации самых разных приспособительных форм, и наоборот — разные эволюционные линии в ходе приспособления к сходным условиям существования могли выработать сходные формы организации (конвергенция!). Пример первого положения — различные уровни организации (Heterokontophyta. Второе положение подтверждается несколькими параллельными эволюционными линиями среди Нeterokontophyta и зеленых водорослей, обсуждаемых дальше.
Отличительная особенность Неterokontophyta — частое использование кремниевой кислоты при образовании и оформлении жесткой клеточной стенки. Пластинки,
чешуйки и отложения из кремниевой кислоты не являются особенностью всех представителей. Так, они полностью отсутствуют, например, у голых клеток Сhloromonadophусеае и иногда в других классах. С другой стороны, как исключение они встречаются в других отделах водорослей (например, Реdiastrum из Сhlorobionta). Однако здесь прослеживается четкая тенденция к образованию этих структур. Целлюлоза явно не служит предпочтительным материалом для построения клеточной стенки. У Сhloromonadophyceae жесткая целлюлозная стенка отсутствует, а у других классов, за исключением бурых водорослей, она широко не представлена.
Приспособление к наземной жизни. Обитателями влажной почвы являются, например, различные диатомеи, а среди желтозеленых водорослей, например, Вatrydium (см. рис. 11.73, С). К тому же классу относится аэрофильная водоросль Сapitulariella, которая распространяется путем рассеивания по воздуху целых спороцист (функционально они могут быть приравнены к апланоспорам) (см. рис. 11.72, С). Мужские репродуктивные клетки Нeterokontophyta обладают (когда имеет место анизогамия или оогамия) хлоропластами, часто сильно редуцированными.
Прогрессивное эволюционное развитие по пути к чистым диплонтампроисходит независимо в разных подвинутых эволюционных линиях. Так, все диатомеи диплонты, а кроме того, Рennales уже не имеют гамет со жгутиками. Среди бурых водорослей прослеживаются усиление в жизненном цикле спорофитного поколения и эволюция в сторону практически чистых диплонтов. У оогамных представителей женские репродуктивные клетки стали неподвижными яйцеклетками.