БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
4. Класс: диатомовые, или кремнеземные, водоросли (Ваcillariophyсеае = Diatomеае)
Диатомеи, насчитывающие свыше 10 000 видов из примерно 200 родов, представляют собой группу крайне разнообразных коккоидных одноклеточных водорослей, иногда формирующих колонии в виде лент или звездочек. Бурые хлоропласта, иногда встречающиеся по одному или по два на клетку, содержат в значительной мере те же пигменты, что и золотистые водоросли. Также совпадают и запасные вещества. Продукты ассимиляции откладываются вне хлоропластов: хризоламинарин — в клеточном соке (у золотистых водорослей — в специальных вакуолях), масло — в особых вакуолях. Только мужские гаметы некоторых видов из порядка Centrales имеют один направленный вперед перистый жгутик; другие жгутиковые стадии неизвестны.
Особое положение в системе диатомеи занимают вследствие наличия внутри внешнего плазматического слоя двух кремнеземных половинок панциря, из которых одна (эпитека),как крышка коробки, накладывается на нижнюю (гипотеку) (рис. 11.76, В). К боковым загибам створок примыкает поясок из двух накладывающихся друг на друга ободков (поисковых ободков). Клетка выглядит по-разному в зависимости от того, рассматриваем ли мы ее со створки (А) или с пояска (В). От пояска иногда внутрь клетки могут отходить неполные перегородки — септы (G).
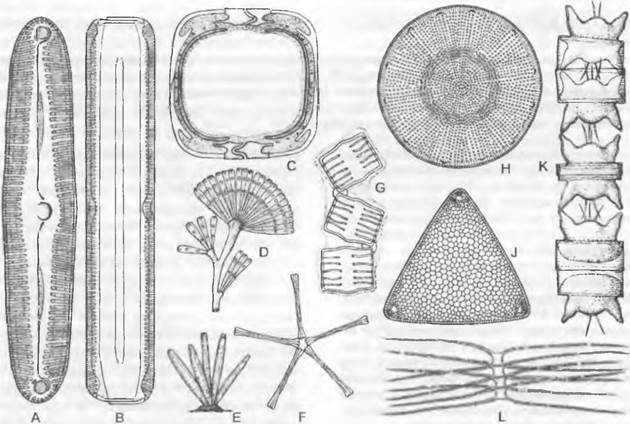
Рис. 11.76. Bacillariophyceae: А — G — Pennales: A — C — Pinnularía viridis, A — вид со створки, со швом (600х), В — вид с пояска (600х), С — поперечный разрез (200х), D — Licmophora flabellata (200х), Е — Synedra gracilis (200х), F — Asterionella formosa (200х), G — Tabellaría flocculosa (400x); H — L — Centrales: H — Coscinodiscus pantocseki (200x), J — Triceratium distinctum (200x), К — Odontella (Biddulphia) aurita (400x), L — Chaetoceros castracanei (250x)
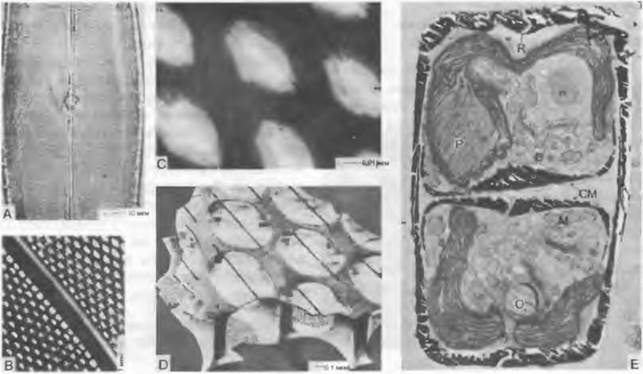
Силикатный (кремниевый) панцирь имеет, особенно на поверхностях створок, чрезвычайно сложно устроенные структуры, часто расположенные рядами; они нередко состоят из крошечных камер, которые могут быть как сверху, так и снизу открытыми или закрытыми и тогда пронизанными тончайшими порами или щелями (рис. 11.77). Силикат является отчасти кристаллическим, отчасти аморфным и поляризационно-оптически изотропным. У створок ископаемых диатомей кристаллическая решетка приобретает аморфное строение. Наряду с силикатом в покрове клеток выявлены также полисахариды («пектины»), белки и жироподобные вещества, однако целлюлоза не обнаружена. Детали для построения панциря образуются в плоских пузырьках под плазмалеммой. Эти пузырьки, возможно, происходят от аппарата Гольджи: очевидно, несколько пузырьков Гольджи сливаются в пузырек, образующий силикат.
Рис. 11.77. Bacillariophyceae, Pennales: A — D — Pleurosigma angulatum, строение кремнеземной створки: А — общий вид средней части створки со швом, В — шов и поры, С — поры, D — реконструкция строения створки по электронномикроскопическому снимку; Е — Gomphonema parvulum, поперечный разрез через клетку в конце деления (10000х); СМ — цитоплазматическая мембрана; D — диктиосомы; М — митохондрия; N — ядрышко; О — капля масла; Р — пиреноид в хлоропласте; R — шов
Вегетативное размножение. Диатомеи размножаются вегетативно путем деления клетки надвое. При этом две половинки панциря (теки) отодвигаются друг от друга за счет увеличения протопласта. Каждая дочерняя клетка, унаследовав от материнской клетки только половину панциря, достраивает недостающую половину, которая всегда бывает гипотекой. Те дочерние клетки, которые наследовали исходную гипотеку (теперь ставшую эпитекой), и образуют соответствующую половину панциря (соответственно новую гипотеку), меньше, чем материнская клетка. Это приводит при дальнейших делениях к прогрессивному уменьшению размеров клеток до определенного минимального размера (примерно половины исходного размера), при котором наступает половое размножение, связанное со значительным увеличением размера зиготы (ауксозиготы). У некоторых видов за счет растяжимости пояска выравниваются размеры эпи- и гипотеки.
Половое размножение. Жизненный цикл диплонтный с гаметической сменой ядерных фаз; клетка диатомовой водоросли содержит, таким образом, диплоидное ядро (в противоположность, например, Zygnematophусеае). При мейозе из диплоидных клеток возникают гаплоидные гаметы.
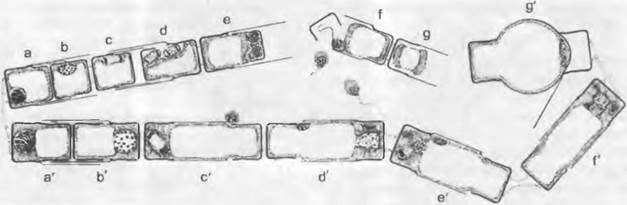
Пример — Centrales (прочие особенности рассмотрены ниже). Определение пола осуществляется модификационно. В клетках, детерминированных как мужские, они чаще всего непосредственно превращаются в спермогоний — возникает 4 сперматозоида (рис. 11.78, d — f) с одним жгутиком каждый. В других клетках, чаще всего более крупных, превращающихся в оогонии, образуются лишенные жгутиков женские гаметы — яйцеклетки. В деталях образование гамет происходит весьма по-разному, и их число может быть разным в зависимости от вида. Сперматозоиды с помощью своего перистого жгутика подплывают к яйцеклеткам. После оплодотворения внутри или вне оогония зигота одевается оболочкой, в которой откладываются силикатные чешуйки. Она вскоре прорастает, причем за счет растяжения стенки вырастает до 2 — 4-кратного размера исходной клетки и превращается в «ауксозиготу»1. Старые половинки панциря отодвигаются друг от друга, и внутри оболочки ауксоспоры образуются новые половинки. При этом возникает диплоидная «первичная клетка», из которой затем, как описывалось выше, при прогрессивном измельчании большей части потомства вегетативным путем возникают новые дочерние поколения.
1 В отечественной литературе для этой структуры применяется название «ауксоспора», хотя даваемое здесь название «ауксозигота», по-видимому, удачнее. — Примеч. пер.
В целом заложение половинок панциря связано с митозами. Даже ядро зиготы при образовании обеих первичных половинок панциря претерпевает митотическое деление, после которого одно из дочерних ядер дегенерирует.
Встречаемость. Диатомеи распространены в пресных водах и морях всех климатических зон. Особенно интенсивно они развиваются весной и осенью, а летом их численность падает. Многие формы живут во влажной почве и на каменистом субстрате. В тропиках они могут обитать совместно с синезелеными водорослями на листьях (эпифильные виды).
Систематика. По симметрии створок Bacillariophyceae подразделяют на два порядка: Centrales и Pennales1. У первой из них створка построена по радиальному плану, у второй — по билатеральному. Помимо строения створок эти два порядка сильно различаются и по типу полового размножения.
1 Согласно современным данным молекулярной систематики, подтверждаемым сведениями по ультраструктуре створок. Centrales представляют собой две эволюционные группы, одна из которых ближе к Pennales, чем к другой из этой группы. — Примеч. пер.
1. Порядок: центрические (Centrales). Контур створки представляет собой окружность или треугольник с закругленными углами (рис. 11.76, Н — L). Элементы орнамента створки имеют при этом радиальное или концентрическое расположение. В противоположность большинству Pennales, вегетативные клетки Centrales неподвижны. Однако мужские гаметы подвижны за счет единственного перистого жгутика (ср. рис. 11.20, А, слева), у которого отсутствуют обе центральные микротрубочки. Половое размножение (см. выше) было особенно детально изучено у Stephanopyxis и Melosira varions.
Распространение. Centrales обитают преимущественно в морях, где составляют существенную часть фитопланктона (первое место среди первичных продуцентов в Мировом океане; см. бокс 11.7). Многие из них имеют особые придатки, способствующие парению (см. рис. 11.76, L), другие объединены слизью в цепочки или соединения иного рода (К).
Виды рода Melosira, образующие нитевидные колонии из коротко-цилиндрических клеток (рис. 11.78), распространены как в морях, так и в пресной воде, многочисленные вилы родов Coscinodiscus и Hemiaulus — только в морях. Для рода Triceratium, также морского (см. рис. 11.76, J), характерны створки с очертаниями от треугольных до многоугольных. Напоминающие обликом консервную банку, клетки Ethmodiscus gazellae (обитателя теплых морей) диаметром почти 2 мм представляют собой самые крупные клетки среди диатомей. Клетки Siephanodiscus снабжены по краю круглых створок венцом из шипов. Род Rhizosolenia распространен преимущественно в морях.
2. Порядок: перистые (Pennales). Клетки имеют форму палочек или лодочек, реже клиновидные (см. рис. 11.76, A — G), поэтому их центр симметрии вытянут по одной линии, от которой перисто отходят элементы орнаментации силикатной стенки. У очень многих форм по продольной оси створки проходит трещина — «шов», тонкое строение которого сильно различается у входящих в этот таксон родов (см. рис. 11.76, А; 11.77). Полагают, что шов играет определенную роль в ползающем движении, встречающемся только у Pennales; у неподвижных (сидячих) форм шов отсутствует. В центре шов прерывается центральным узелком, а у концов створок заканчивается концевыми узелками (см. рис. 11.76, А).
Рис. 11.78. Bacillariophyceae, Centrales, Melosira varians. Половое размножение (схема): а — g — мужской, а' — g' — женский участки нити; а — е, а'—е' — мейоз; f — открывшийся; g — освободившийся спермогоний; d' — мужское ядро, внедрившееся через специальную щель; f' — оплодотворение; g' — молодая ауксозигота
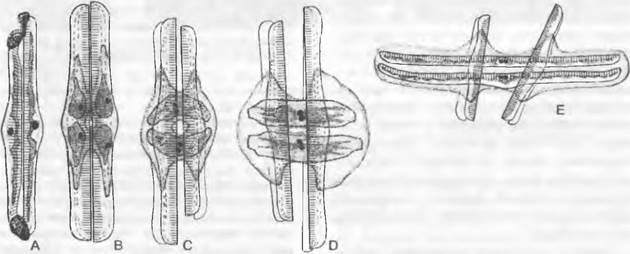
Половое размножение Pennales отличается от «нормального типа» Centrales, поскольку никаких подвижных гамет не бывает. Здесь сливаются изогаметы в форме голых протопластов (единственное исключение — оогамия у Rhabdonema; причем ♂ гаметы все же лишены жгутиков). Для спаривания две вегетативные клетки сползаются вместе и чаще всего обильно выделяют слизь. Ядро каждой клетки делится мейотически на 4 гаплоидных ядра, из которых, однако, 2 дегенерируют, а 2 становятся ядрами гамет. Эпи- и гипотека несколько отходят друг от друга. Через получающийся разрыв между ними гаметы из разных клеток имеют возможность попарно копулировать, так что возникают две зиготы, которые сразу же вырастают в ауксозиготы. Каждая из них выделяет пару силикатных половинок панциря и становится первичной клеткой для последующих клеток разного размера. Первичные и родительские клетки располагаются перпендикулярно (рис. 11.79) или параллельно друг другу. От этого обычного варианта имеется множество отклонений.
Распространение. Большинство подвижных Pennales обитают преимущественно на дне пресных, солоноватых или соленых водоемов (может наблюдаться массовое их развитие), эпифитно на водных растениях или в почве, однако имеются и планктонные формы. Примерно 500 видов рода Navícula, клетки которых напоминают лодочки, распространены в водоемах всех типов; виды рода Pimularia (см. рис. 11.76, А — С) с похожими, но удлиненно-эллиптическими клетками предпочитают пресную воду. Виды Pleumsigma, клетки которых слегка S-образно изогнуты (см. рис. 11.77), могут служить как тест-объект для проверки объективов микроскопов. У видов родов Diatoma, Tabellaría (см. рис. 11.76, G), Frágil aria и др., обитающих преимущественно в стоячих пресных водах, клетки образуют длинные цепочки, у Asterionella (F) — звездчатые, у Meridion — вееровидные (иногда замыкающиеся в кольцо) колонии. Некоторые виды рода Synedra свободно парят в воде, другие прикрепляются слизистыми подошвами к более крупным нитям водорослей (Е). У также ведущих сидячий образ жизни видов Licmophora клетки после деления остаются соединенными друг с другом, так что возникают древовидные колонии из клеток, сидящих на ножках из слизи (D). Можно перечислить еще роды, упоминавшиеся в других разделах: Surirella (см. бокс 11.7), Nitzschia (см 13.8), Gomphonema (см. рис. 11.77, Е) и Rhopaludia (см. рис. 11.79).
Рис. 11.79. Ваcillariophусеае, Реnnales, Rhopalodia gibba, половое размножение: А — две клетки, соединенные слизистым чехлом; В — деление материнских клеток (дегенерировавшие ядра уже исчезли); С — образование зиготы после слияния гамет; D — увеличение размера аук- созиготы (А — D 410х); Е — конечная стадия и образование нового панциря (240х)
Филогения. Centrales, имея снабженные жгутиками сперматозоиды, являются более ранней группой, чем Pennales, у которых жгутики полностью утрачены. Предками диатомей были, возможно, водоросли типа Chrysomonadales, клетки которых уже могут нести кремнеземные чешуйки. Диатомеи в основном остались на коккоидном уровне организации, имея лишь первые намеки на нитчатую организацию.
Древнейшие диатомеи, и именно центрические формы, известны из юры. Большое видовое разнообразие их отмечается с мела.
В третичном и межледниковом периодах массовое развитие диатомей привело даже к образованию соответствующих пород (применение — см. бокс 11.6, с. 247).
В противоположность обоим классам, рассмотренным ранее, у следующего класса отсутствуют простые формы организации (например, нет одноклеточных форм или неразветвленных нитей). Часто образуются макроскопические тканевые талломы с сильно выраженной дифференциацией на органы и ткани. Встречаются различные типы смены поколений В состав клеточной стенки входят только альгинаты и фукоидан.