БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
Первый отдел: зеленые водоросли I (Chlorophyta)
У зеленых водорослей встречаются почти все уровни организации. За исключением амебоидных форм (которые, правда, иногда представлены репродуктивными клетками), здесь выработались все типы, даже тканевый и ложнотканевый (соответственно, например, Ulva и Codiurri). Это могут быть микроскопически мелкие одноклеточные организмы, нитчатые водоросли — неразветвленные или разветвленные, часто образующие плотные пучки (см. рис. 11.90), а также сложно устроенные листовидные талломы, которые отчасти напоминают высшие растения.
Образование поперечных перегородок осуществляется преимущественно в фикопластах: здесь микротрубочки во время телофазы собираются в экваториальной плоскости между разделяющимися дочерними ядрами. При этом возникает клеточная пластинка с порами для плазмодесм (см. 2.2.1).
Клетки, имеющие жгутики, чаще всего грушевидные, радиально-симметричные. Жгутики одинаковой длины в количестве 2 или 4 (редко их много) расположены на переднем конце клетки (терминально); эти подвижные клетки изоконтные, снабжены «гладкими» жгутиками. Они нередко содержат сократительные вакуоли (чаще всего 2), а также (в задней по ходу движения части) изогнутый или бокаловидый постенный хлоропласт1 с глазком (стигмой; см. рис. 11.94, А) или без него. Красный глазок состоит из глобул, которые содержат каротины; он не связан (как у Еuglenophyta, Еustigmatophyta и Heterokontophyta) с расширением жгутика.
1 У немногих представителей хлоропласт может занимать и центральное положение. — Примеч. пер.
Понятие «гладкие жгутики» означает, что они не несут трубчатых волосков, однако могут иногда иметь очень тонкие волоски или чешуйки разного типа.
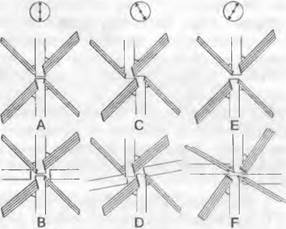
Большое значение для систематики хлорофитов имеет ультраструктура жгутикового аппарата в области прикрепления жгутиков (рис. 11.88). Он состоит из базальных телец (т.е. нижних концов жгутиков в клетке), микротрубочковых корней вместе с ассоциированными с ними структурами, а также ризопласта (или ризопластов). Последние представляют собой соединения между базальными тельцами жгутиков и клеточным ядром. Жгутики чаще всего прикрепляются по крестообразному типу: четыре крестовидно расположенных микротрубочковых корня закрепляют базальные тельца жгутиков в клетке. При расположении «1 час — 7 часов» (сокращенно расположение 1 — 7) два базальных тельца располагаются, если смотреть на клетку сверху, как цифры 1 и 7 на циферблате часов. Аналогично следует понимать расположения «12 часов — 6 часов» и частый вариант «11 часов — 5 часов».
Рис. 11.88. Chlorobionta, Chlorophyta. Микротубулярная корневая система жгутикового аппарата. Вид на базальные тельца с корнями из двух и четырех микротрубочек в крестообразном расположении. Верхний ряд (А, С, Е) — двухжгутиковые; нижний ряд (В, D, F) — четырехжгутиковые клетки. А, В — тип «12 часов — 6 часов»: гипотетический исходный жгутиковый аппарат, у которого базальные тельца (каждая пара находящихся друг против друга жгутиков) расположены на одной линии; С, D — тип «11 часов — 5 часов»: базальные тельца (по сравнению с А, В) слегка сдвинуты относительно друг друга против часовой стрелки; Е, F — тип «1 час — 7 часов»: базальные тельца (по сравнению с А, В) слегка сдвинуты относительно друг друга по часовой стрелке
Бокс 11.8. Применение водорослей
Из золы различных бурых водорослей(Phaeophyceae: Laminariales) можно получать йод; именно таким путем его получали еще до 30-х годов прошлого столетия. Пригодные для этого бурые водоросли могут накапливать в клетках йод (до 0,3 % сухой массы) из морской воды, где его концентрация 0,000005 %. Кроме того, бурые водоросли дают альгинаты,коллоидные свойства которых находят многостороннее применение в текстильной, пищевой промышленности, фотоиндустрии и производстве косметики. Мировая продукция их около 14000 т в год. Они используются, например, при приготовлении мороженого, пудинга, мазей, зубной пасты, диетических продуктов для похудения, капсул для лекарств, клея, красок и т. д. Из бурых водорослей получают также соду и маннит. В Китае и Японии бурые водоросли используются в пищу (комбу).
Из клеточных стенок нескольких видов красных водорослей(Rhodophyta) получают полисахариды для лекарственных и технических целей. Находят применение каррагенан из Chondrus crispus и Gigartina mamillosa с побережья Северного моря (в высушенном виде известен еще под названием «ирландский мох») и агар из различных флоридей Тихого океана (таких, как виды Gelidium и Gracilaria), а в последнее время также из европейских видов. Япония, добывая ежегодно 2 000 т агара, является важнейшим его продуцентом (использование для культур микроорганизмов, в пищевой и фармацевтической промышленности). Рогphyra (культивируемая по морским побережьям Восточной Азии на плантациях, представляющих собой подвешенные у поверхности воды решетки) широко используется в пищу, (нори) особенно в восточноазиатских странах.
Зеленые водоросли (Chlorophyta) имеют меньшее значение, чем красные и бурые. В Западной Сибири проводится массовый сбор нитчатых зеленых водорослей (около 1 000 000 т ежегодно с нескольких тысяч квадратных километров), которые используются для производства бумаги, а также изоляционного и строительного материала (альгилит). Для биотехнологии пригодны коккоидные зеленые водоросли (Chlorella, Scenedesmus) благодаря высокой активности фотосинтеза и возможности массового культивирования. Соответствующие исследования направлены на получение белков и витаминов для питания человека, и животных (максимальные урожаи в культурах под открытым небом в тропиках — 5 т на гектар ежемесячно). С помощью «водорослевых реакторов» можно осуществлять биологический газообмен (СО2 на О2 при фотосинтезе). Испытаны соответствующие приспособления для использования этих водорослей в качестве источников кислорода и питательных веществ (например, для космических кораблей).
Прочие водоросли. Образуемые из диатомей (Bacillariophyceae) породы (диатомовая земля, полировочные сланцы, кизельгур) раньше использовались как материал для строительства, чистки и шлифовки. И поныне они еще используются как фильтрующая масса (например, для очистки воды) или как абсорбент и наполнитель. Порода, состоящая из кокколитов(Haptophyta), раньше находила применение как писчий мел.
При половом размножении у подавляющего большинства образуются гаметы, имеющие жгутики. При этом копулируют 2 гаметы (ср. рис. 11.89, F), которые часто очень сходны с зооспорами (спорами бесполого размножения) и возникают в одноклеточных гаметангиях. Мужские гаметы, как правило, имеют жгутики, женские (яйцеклетки) могут быть также неподвижными (например, см. рис. 11.100, Е). Простой гаплонтный жизненный цикл часто усложняется наличием диплофазы (т. е. становится гаплодиплонтным). Некоторые представители за счет редукции гаметофита стали диплонтами (как Саulerpасеае). Таким образом, исходно зиготическая смена ядерных фаз становится промежуточной, или гаметической. Продукт копуляции — зигота — у пресноводных форм чаще всего представляет собой толстостенную, округлую покоящуюся клетку (цистозиготу), нередко окрашенную каротиноидами в красный цвет.
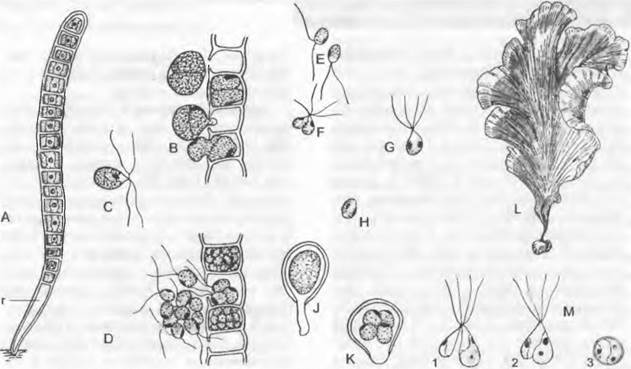
Рис. 11.89. Ulvophyceae: А — К — Ulothrix, U.zonata: А — молодые нити с ризоидальной (r) клеткой (300х); В — фрагмент нити с освобождающимися зооспорами, которые возникают по две в каждой клетке; С — отдельные четырехжгутиковые митозооспоры; D — образование и выход имеющих меньший размер двухжгутиковых гамет из фрагмента нити; Е — гаметы; F — их копуляция; G, Н — зигота; J — зигота, прорастающая после периода покоя; К — образование мейозооспор в зиготе (В — К — 480х); L — Ulva, U. lactuca (морской салат) на камне, клетки по кромке бесцветные из-за выхода зооспор (1/2х); М — Entercmorpha, Е. intestinalis, копуляция анизогамет и зигота (1 800х); 1 — 3 — стадии до образования зиготы
К зеленым водорослям относятся примерно 450 родов и 7 000 видов, большая часть которых (около 90%) обитает в планктоне или бентосе (см. бокс 11.7) пресных вод. Некоторые виды с более крупными талломами встречаются также в морях, обычно у берегов; в морском планктоне, напротив, зеленые водоросли составляют лишь незначительную часть. Некоторые зеленые водоросли живут вне воды: во влажной почве или на ее поверхности, как эпифиты на деревьях и т. д. Определенные виды даже выдерживают значительное высыхание и являются выраженными наземными растениями. Некоторые живут как симбионты в лишайниках или как внутриклеточные эндосимбионты в низших животных («зоохлореллы», например, в гидре). Некоторые представители утратили ассимиляционные пигменты и живут гетеротрофно. Родство их с автотрофными формами зеленых водорослей можно устанавливать по другим признакам. Систематическое положение отдельных родов зеленых водорослей остается пока неясным.
1. Класс: Рrasinophyceae
Представители класса имеют своеобразные чешуйки на поверхности клеток, а также 2 или 4 одинаковых жгутика (изредка жгутик только один). Относящиеся сюда монадные (например, Рyramimonas, Pedinomonas, Рlatymonas), отчасти также капсальные и коккоидные организмы обитают в основном в планктоне морей, лишь немногие виды — в пресных водах. Рlatymonas convolutae — эндосимбионт морского плоского червя.
У последующих классов (2 — 7) жгутики прикрепляются по крестообразному типу и жгутиковый аппарат показывает расположение 11 — 5 своих ультрамикроскопи
ческих структур (у 8-го класса — Сhlorophyceae, напротив, расположение 1 — 7 или 12 — 6).
2. Класс: Ulvophyceae
Одноклеточные (коккоидные) и многоклеточные — нитчатые, паренхиматозные или сифонокладальные — представители этого класса, за исключением репродуктивных клеток, не имеют жгутиков. Жгутиковый аппарат (расположение 11 — 5) показывает четкий сдвиг базальных телец относительно друг друга. Образование клеточной стенки происходит бороздой без участия фикопласта. В поперечных перегородках плазмодесмы отсутствуют. Стенки содержат полисахариды. Большинство видов встречается в морских или солоноватых водах.
Талломы представлены преимущественно неветвящимися нитями, которые удлиняются за счет («диффузного») поперечного деления многих или всех клеток (нитчатый уровень организации). У рода Monostroma более старые нити за счет продольных делений клеток в одной плоскости становятся уплощенными. Крупный листовидный зеленый двухслойный паренхиматозный таллом образуется у обитающей в прибрежной зоне морей водоросли Ulva lactuca (рис. 11.89, L). Enteromorpha, водоросль также прибрежных вод, однако встречающаяся и в соленых внутренних водах, имеет таллом в виде кишки, в сечении округлой или уплощенной. Полярность у некоторых видов выражена лишь слабо. Так, например, у Ulothrix она определяется наличием единственной бесцветной ризоидальной клетки, не способной к делению (рис. 11.89, А). Клетки имеют ядро и по одному постенному поясковидному хлоропласту, имеющему форму цилиндра, закрытого либо с продольными отверстиями или же форму изогнутой пластинки с одним или несколькими пиреноидами. После ядерного деления для разграничения дочерних клеток сразу же втягивается клеточная стенка (ср. Chlorococcales).
Бесполое размножение происходит зооспорами, половое — путем копуляции гамет, имеющих жгутики. Цикл развития осуществляется или как чисто гаплонтный с
зиготической сменой ядерных фаз (Ulothrix)1 либо гаплодиплонтный как гетерофазная смена поколений (Ulva).
1 У Ulothrix зигота непосредственно перед мейозом прорастает в маленькое растеньице грушевидной формы; туда переходит диплоидное ядро, где уже претерпевает мейоз. Это растеньице часто трактуют как диплоидную стадию этой водоросли (Соdiolum-стадия), и тогда уже нельзя говорить о зиготической смене ядерных фаз. — Примеч. пер.
1. Порядок: Соdiolales (Ulotrichales). Часто встречающийся в пресных водах вид (Ulothrix zonata (рис. 11.89, А) образует неразветвленные, интеркалярно нарастающие нити. Их короткие клетки содержат один лентовидный хлоропласт, который в виде незамкнутого с одной стороны кольца прилегает к клеточной стенке. Нити прикрепляются ризоидальной клеткой, чаще всего бесцветной, к камням и другим субстратам. Кроме ризоидальной клетки, любая другая клетка может служить для размножения; при этом в клетках ничего не остается. В жизненном цикле (см. рис. 11.101, А; 11.89, А — К) 4-жгутиковые гаплоидные митозооспоры обеспечивают бесполое размножение. Они имеют глазок и хлоропласт и образуются одновременно путем распада протопласта, становящегося перед этим четырехъядерным, на одноядерные подвижные клетки, которые выходят (В) через боковое отверстие в стенке материнской клетки (= спороцисте) После фазы активного движения они прикрепляются, выделяя слизь, жгутики и глазок исчезают, и клетки вырастают в новую гаплоидную, полярную нить. При неблагоприятных условиях сходным образом, но в значительно большем числе (D, Е) образуются изогаметы; они внешне сходны с зооспорами, но меньше и обладают только 2 жгутиками. Гаметы разных типов скрещивания (+, -) сливаются (F) попарно в зиготу (G). Зигота, сначала плавающая при помощи 4 жгутиков, втягивает жгутики, округляется (Н), одевается прочной стенкой (цистозигота) и окрашивается каротиноидами в красный цвет. Она представляет собой покоящуюся стадию, которая заканчивается мейозом и следующим за ним прорастанием 4 — 16 гаплоидными мейозооспорами (К)2. При этом происходит разделение на два типа скрещивания. Мейозооспоры прикрепляются к субстрату боком с образованием ризоида, так что их продольная ось при начинающемся делении становится поперечной осью. Теперь развиваются гаплоидные нити с генотипическим определением полового знака (А), которые кроме гамет могут давать также митозооспоры. Таким образом, эти растения гаплонт- ны с зиготической сменой ядерных фаз. Зигота (J), образующая палочковидный придаток, может отчасти рассматриваться как крайне недоразвитый спорофит.
2 Структура, в которую прорастает зигота, иногда трактуется как диплоидное поколение (см. предыдущую сноску). — Примеч. пер.
Codiolales обитают как в пресной воде, так и в морях, отчасти также в почве. Талломы водных представителей часто прирастают к субстрату.
У Monostroma grevillei листовидный тканевой (мужской или женский) таллом образует гаметы. Зигота развивается в самостоятельный, внедряющийся в известковые раковины морских животных пузырек (Codiolum). У некоторых видов размер зиготы увеличивается в 20 (и более) раз от исходного диаметра.
2. Порядок: Ulvales с родами Ulva (морской салат) и Enteromorpha (кишечница). Здесь происходит гетерофазная изоморфная смена поколений: вклинивается диплоидная спорофитная фаза.
Эту ситуацию можно кратко рассмотреть на примере Ulva — морского салата. Гаметофит и спорофит имеют внешне одинаковые листовидные тканевые талломы (рис. 11.89, L). Смена поколений аналогична видам рода Cladophora (см. рис. 11.101. В). Гаметофиты генотипически (+ и -) дифференцированы. Между двухжгутиковыми гаметами противоположных типов скрещивания происходит изогамная копуляция. Образующиеся таким образом зиготы прорастают, как у многих морских водорослей (постоянно при благоприятных условиях для роста в противоположность пресноводным формам), сразу в диплоидный спорофит. Он дает четырехжгутиковые зооспоры, с образованием которых связаны как мейоз, так и генотипическое определение пола. Из зооспор вновь возникают гаплоидные гаметофиты, различающиеся по полу. Как гаметофиту, так и спорофиту предшествует нитчатый предросток, который в результате продольных делений клеток становится листовидным тканевым талломом.
Род Enteromorpha отличается от рода Ulva анизогамией. Определение пола гаметофита генотипическое; мужские талломы дают ♂ гаметы меньшего размера, женские — более крупные ♀гаметы с зеленым хлоропластом (ср. рис. 11.89, М). У Hormidium изучали движение хлоропластов (см. 8.2.2).
В родстве с рассмотренными порядками находится и порядок Acrosiphonales. Твердая фракция клеточной стенки состоит из переплетенных в виде войлока целлюлозных фибрилл (тип целлюлозы отличается от кристаллической целлюлозы рассматриваемых ниже Cladophorophyсеае и большинства листостебельных растений). Талломы Spongomorpha(гаметофит) могут быть переплетенными, как войлок. Смена поколений гетероморфная, со спорофитом типа Codiolum. Сюда же относится Urospora (см. рис. 11.102).
3. Класс: Trebouxiophyceae
К этому классу относятся водоросли, обитающие вне воды1, в том числе симбионты лишайников, от коккальной до разветвленно-нитчатой организации. Жгутиковый аппарат (у репродуктивных клеток) со сдвинутыми относительно друг друга базальными тельцами (расположение 11 — 5) показывает особенности, которые встречаются только в этом классе. Дочерние клетки, возникающие в результате схизогонии (быстро следующих друг за другом клеточных делений в материнской клетке), окружаются заново образующейся клеточной стенкой. Этим Trebouxiophyceae сходны с Chlorococcales из Chlorophyceae, однако митоз у них протекает иначе2. Виды родов Тге- bouxia и Chlorella, отличающиеся коккалъным типом таллома (некоторые с агитаноспорами, см. рис. 11.97, J), представлены симбионтами в лишайниках3, а также встречаются в плазме низших животных (Chlorella vulgaris в инфузориях, Chlorohydra и др., ср. 9.2).
1 Виды рода Chlorella очень широко встречаются не только вне воды, но и в пресных водах. — Примеч. пер.
2 Следует уточнить, в чем именно состоит отличие: у Trebouxiophyceae при митозе центриоли располагаются по бокам веретена (т. е. веретено метапентрическое). тогда как у других зеленых водорослей и многих других «протестов» — по полюсам веретена. Это основная особенность Trebouxiophyceae. — Примеч. пер.
3 Виды рода Chlorella в лишайниках не встречаются. — Примеч. пер.
Сюда же примыкают Prasiolaceae. Prasiola stipitata имеет своеобразный жизненный цикл. В чисто вегетативных верхних частях листовидного спорофита происходят мейозы, за которыми следуют митозы. Развивающийся таким путем гаметофит остается на протяжении всей своей жизни сросшимся со спорофитом. Отдельные поля клеток гаметофита производят яйцеклетки, другие — мелкие двухжгутиковые cf гаметы (генотипическое определение пола оогамия). Вышедшие на свободу репродуктивные клетки сливаются затем в зиготу.
Последующие классы стоят весьма изолированно; их родственные связи с другими Chlorophyta еще не выяснены.
4. Класс: Cladophorophyceae
Виды этого класса относятся исключительно к сифонокладальному типу организации; разветвленные, иногда неразветвленные нитчатые талломы образованы многоядерными клетками. Клеточные стенки состоят из целлюлозы в виде фибрилл и с той же структурой, что и у наземных зеленых растений. Подвижные репродуктивные клетки имеют 2 или 4 жгутика; для жгутикового аппарата (расположение 11 — 5) характерно наложение базальных телец. Обитают преимущественно в морях, реже в пресных водах.
Часто обильно разветвленные талломы видов единственного порядка Cladophorales многоклеточные, а каждая клетка многоядерная. Многоядерные клетки встречаются также у видов других порядков (например, Hydrodictyon из Chlorococcales). Виды рода Cladophora имеют чаще всего гетерофазную изоморфную смену поколений (см. рис. 11.101, В). При этом каждое поколение может воспроизводиться бесполым путем. Изогаметы двухжгутиковые, тогда как мейо- зооспоры снабжены 4 жгутиками (у пресноводных видов их 2). Часто образующая в пресных водах пучки длиной до 30 см и более Сladophora glomerata размножается только бесполым путем1.
1 Есть сведения, что у С. glomerata имеется и половой процесс. — Примеч. пер.
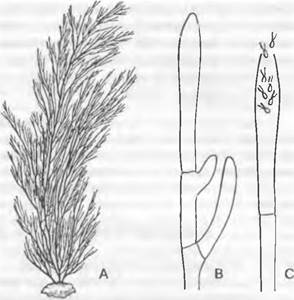
Пучки нитей видов Сladophora, часто встречающиеся на твердом субстрате в пресных водах (в том числе проточных) и в морях на твердом субстрате (рис. 11.90), прикрепляются у основания клеткой типа ризоида и показывают преимущественно верхушечный рост. Ветвление происходит путем выпячивания «стволовой» клетки (каждый раз под центрипетально образованной перегородкой); они продолжают рост при образовании перегородки под острым углом к продольной оси. Постенный хлоропласт сетевидный2. Он содержит пиреноиды с зернышками крахмала. Целлюлозные микрофибриллы клеточной стенки расположены слоями под разными углами, придавая ей большую прочность. Как и у Ulvophусеае. подвижные репродуктивные клетки (зооспоры и изогаметы) возникают в практически неразличимых внешне клетках, однако, как правило, на концах боковых ветвей (Siphonocladus — морская водоросль).
2 На самом деле у Сlodophora не один хлоропласт, а много удлиненных хлоропластов неправильной формы, смыкающихся концами в постенную сеть, так что выглядят они как единая сетчатая пластинка. — Примеч. пер.
Рис. 11.90. Сladophorophусеае, Сladophora: А — внешний вид (1/Зх); В — ветвление; С — гаметангий с гаметами (В, С — 250х)
Примыкающий к Сladophorales порядок Valoniales (рис. 11.91, Е; раньше его относили к Вryyopsidophyceae) отличается особым механизмом деления клеток. Протопласт первоначально не разделенного на клетки мешковидного таллома делится на несколько фрагментов разного размера, которые округляются часто еще внутри исходного таллома и окружаются новыми клеточными стенками. Таким путем может возникнуть многоклеточный псевдопаренхиматозный таллом. Талломы Valonia, содержащие крупную вакуоль, много ядер и многочисленные постенные хлоропласты, представляют собой удобный объект для изучения проницаемости клеточных стенок (см. рис. 2.66, В).
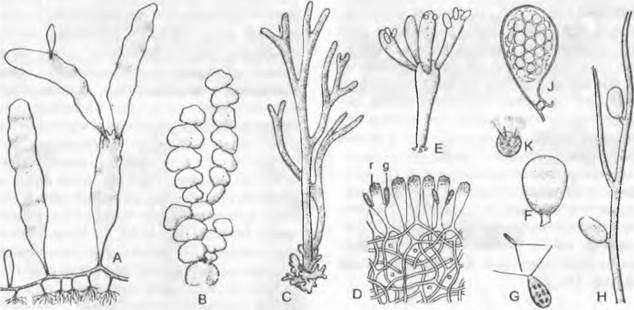
Рис. 11.91. Bryopsidophyceae: A — D — Halimedales: A — Caulerpa proliféra, таллом (12x), B — Halimeda tuna, таллом (1/2x), C, D — Codium tomentosum: С — таллом (1/2x), D — поперечный разрез таллома (15x); E — Valoniales (ныне отнесены к Cladophorophyceae); Valonia utricularis, таллом (11/2x); F — K— Bryopsidales; F, G — Derbesia marina («Halicystis ovalis»); F — гаметофит (3x), G — ♂ и ♀ гаметы (500x); H, J, K — Derbesia marina: H — фрагмент таллома спорофита (30x), J — спорангий (120х), К — зооспора (400х); g — гаметангий; г — кортикальный пузырь (вздутая клетка корового слоя)
5. Класс: Вryopsidophусеае (= сифоновые)
Сифоновые, особенно распространенные в теплых морях, имеют чрезвычайно разнообразный внешний вид. Их таллом не имеет поперечных перегородок, может быть только система распорок. Клеточная стенка (в качестве компонентов которой наряду с целлюлозой встречаются маннан и ксилан) окружает, таким образом, один- единственный полиэргидный протопласт, содержащий многочисленные мелкие дисковидные хлоропласты. Только вместилища репродуктивных клеток отделяются перегородкой (сифональный уровень организации). Талломы некоторых видов представляют собой сложные сплетения нитевидных элементов. К набору пигментов, характерному для зеленых водорослей, у сифоновых добавляются сифоноксантин и сифонеин — отличительные для данного класса дополнительные пигменты.
Половое размножение анизогамное, реже изогамное Репродуктивные клетки имеют 2, 4 или много жгутиков. Жгутиковый аппарат (расположение 11 — 5) показывает наложение базальных телец. Жизненный цикл, согласно новейшим исследованиям, преимущественно гаплонтный, однако возможна и гетероморфная смена поколений с дикариотично-диплоидными спорофитами. Это можно рассмотреть на примере Derbesia-Halicystis. Гаметофит состоит из пузыревидного гаметангия размером 0,5 — 3 см, от которого отходит многолетний ризоид. Из-за незнания связи со спорофитом раньше это растение помещали в отдельный род («Halicystis» = гаметофит Derbesia; рис. 11.91, F). Раздельнополые растения «Наlicystis» дают анизогаметы с двумя жгутиками равной длины (G). Из зиготы вырастает спорофит с разветвленно-нитевидным талломом — Derbesia (Н). В яйцевидных спороцистах этого растения, сначала дикариотичного, затем местами диплоидного, возникают после мейоза мейозооспоры с венцом жгутиков. Смена поколений гетероморфная со слабым преобладанием дикариотично-диплоидного спорофита, достигающего до 10 см в высоту (см. рис. 11.101, С). Состав клеточной стенки у этих поколений, как и у Вгyopsis (см. ниже), различен.
1. Порядок: Вгуopsidales. Виды порядка не имеют амилопластов, служащих для образования крахмала. Виды типового рода Вгyopsis образуют ползучие «корневища», прилегающие к субстрату, и вертикальные ветви, которые в верхней части перисто разветвлены. Жизненный цикл гаплонтный. Клеточная стенка растеньиц, образующих гаплоидные гаметы, состоит преимущественно из ксилана, стенка зиготы — в основном из маннана. Derbesia (Наlicystis) встречается по Атлантическому побережью.
Сodiасеае отличаются талломами длиной до нескольких метров (например, некоторые виды рода Сodium; рис. 11.91, С, D), представляющими собой сплетение разветвленных, лишенных поперечных перегородок нитей, жесткость которым придают кольца на клеточной стенке. Клеточная стенка состоит преимущественно из маннана. В ископаемом виде Сodiасеае известны уже из нижнего палеозоя.
2. Порядок: Наlimedales. Клетки представителей этого порядка имеют как хлоропласты, так и амилопласты. Клеточные стенки не содержат маннана.
Семейство Саuliеграсеае с распространенным в более теплых морях родом Саulerpa, представленным самыми разнообразными формами, отличается талломами, состоящими из бесцветных ползучих основных осей длиной до 1 м с идущими снизу в субстрат ризоидами, а вверх — лопастями различного облика, которые могут достигать размеров в несколько дециметров (А). Крупные растения состоят из одной-единственной многоядерной гигантской клетки, внешняя стенка которой поддерживается только многократно ветвящимися тяжами в виде балок. Клеточная стенка содержит преимущественно ксилан. Происходит ли при образовании гамет (они выходят зелеными облачками, после чего опустевшее растение отмирает) мейоз, остается неясным.
У представителей распространенного в более теплых морях рода Наlimeda (В) таллом состоит из дисковидных элементов, стенки которых инкрустированы известью.
6. Класс: Dasycladophусеае
Dasycladophусеае (Dasycladales) отличаются от типичных сифоновых (предыдущий класс) радиальной симметрией таллома и волосовидными выростами (см. рис. 4.1), которые отчасти могут сбрасываться, оставляя при этом рубцы. Центральная ось несет мутовчато расположенные боковые ветви. Виды этого класса встречаются исключительно в морях.
Клеточная стенка состоит преимущественно из маннана. Таллом образован прикрепленной ризоидами к субстрату «стволовой клеткой» и мутовчато отходящими от нее боковыми ветвями (рис. 11.92, В). Эти ветви могут быть простыми или разветвленными и часто заканчиваются гаметангием.
Рис. 11.92. Dasycladophyceae, Dasycladales. Cymopolia barbata: А — верхняя часть растения (4х); В — продольный разрез через часть таллома, мелкими точками показан известковый покров (40х); С — G — Acetabularía mediterránea: С — взрослые талломы (естественная величина), D — продольный разрез через «зонтик», вверху венец из стерильных выростов, внизу рубцы из отпавших мутовок стерильных выростов (6х), Е — вскрывшаяся циста, из которой выходят гаметы (100х), F — гаметы (300х), G— копуляция (30х)
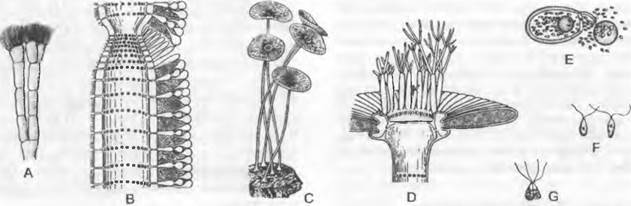
В качестве объекта морфогенетических исследований особенно известна Асеtabularia (рис. 11.92, С — G). Эта водоросль несет на неразделенной ножке «шляпку» в виде зонтика, которая состоит из радиально расположенных, плотно прилегающих друг к другу камер. Над зонтиком и под ним образуется по венцу из коротких клеток. Из верхнего венца возникает дополнительно мутовка тонких, разветвленных вверху ветвей, которые при созревании зонтика быстро отмирают. Таллом имеет сначала лишь одно-единственное первичное ядро, которое долгое время находится без изменений в ризоиде. После развития зонтика оно делится на многочисленные гаплоидные вторичные ядра, которые мигрируют в камеры, после чего начинается образование толстостенных цист. Цисты освобождаются после разрушения зонтика, вскрываются крышечкой и высвобождают гаметы (Е). Зигота (G), возникающая в результате копуляции двух изогамет, прикрепляется к дну и вырастает в новый диплоидный таллом. Согласно недавним исследованиям, Aceta- bularía не диплонт, так как уже первичное ядро, по-видимому, является гаплоидным. Однако согласно другой точке зрения мейоз происходит при образовании вторичных ядер; тогда жизненный цикл был бы диплонтным (с гаметической сменой ядерных фаз).
Внешние слои клеточной стенки у Dasycladaceae очень сильно обызвествляются (В), так что после отмирания таллома остается пронизанная отверстиями известковая трубочка; на этом основана значительная породообразующая роль ископаемых Dasycladaceae, например, в альпийском триасе. Начиная с кембрия известно в целом 120 родов Dasycladophyceae во всех формациях, тогда как в настоящее время — только 10 родов. На основании ископаемых находок мы можем проследить эволюцию от простых форм, у которых ветви отходят беспорядочно от стволовой клетки до высокодифференцированных, таких, как виды рода Acetabularia.
7. Класс: Trentepohliophyceae
Нитчатый таллом часто гетеротрихальный, т. е. разделенный на стелющиеся и восходящие нити, которые могут быть разветвленными (рис. 11.93, С). Клетки одноядерные. У некоторых форм стелющиеся нити склеиваются в плоский диск (Cephaleuros), как и у некоторых представителей Klebsormidiophyceae (Coleochaete, см. рис. 11.106, А). Гетеротрихальным строением таллома Trentepohliophyceae напоминают еще не рассматривавшиеся Chaetophorales из Сhlогоphyceae (Stigeoclonium; см. рис. 11.99, А, В). Исключительно дня Trentepohliophyceae характерны дополнительные структуры в виде колонок в жгутиковом аппарате (ориентация 11 — 5) с налегающими друг на друга базальными тельцами, а также с билатеральными килевидными углублениями в жгутиках.
Рис. 11.93. Trentepohliophyceae: А, В — Pleurococcus, Р. naegelii (600х); С — Е — Trentepohlia: С — Т. аuгеа, участок стелющейся нити с вертикальными ветвями (на вершине правой вертикальной нити одна клетка с зооспорангием, от второй зооспорангий отделился; 500v), D — Т. umbrina, зооспорангий, из которого выходят зооспоры (300х), Е — Т. umbrina,отпадение освободившегося спорангия (300х)
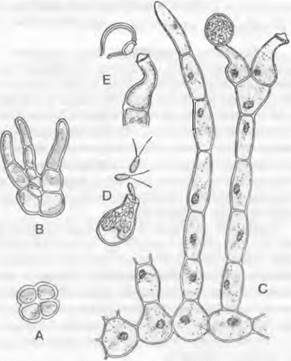
Состоящая из полисахаридов клеточная стенка может дополнительно образовывать слой из спорополленина. При клеточном делении новые клеточные стенки закладываются во фрагмопласте. Большинство видов — наземные водоросли (например, эпифиты на коре деревьев или камне).
Тгentepohlia (рис. 11.93, С) часто встречается как симбионт в лишайниках или как наземная водоросль на скалах (Т. аuгеа — на известковых, а напоминающая запахом фиалку Т. iolithus — на силикатных породах) и стволах деревьев, а в тропиках и на кожистых листьях. Приспособление к наземному образу жизни проявляется также в том, что спороцисты с находящимися в них зооспорами часто отрываются целиком. Двухжгутиковые репродуктивные клетки копулируют как гаметы друг с другом или служат для бесполого размножения (факультативное определение функции). Очень распространенные зеленые налеты на коре деревьев и скалах вызываются водорослями типа «Рleurococcus» (Apatococcus и Desmococus)1; некоторые из этих наземных водорослей больше не образуют подвижных клеток, т. е. они претерпели соответствующую редукцию.
1 Род Desmococcus в большинстве современных систем относится к классу Тгebouxiophусеае (см. выше). — Примеч. пер.
8. Класс: Сhlorophyceae
Эволюционно Сhlorophусеае в противоположность Рrasinophусеае (1), Ulvophyсеае (2) и Тгebouxiophусеае (3), несмотря на их часто примитивную организацию, следует, возможно, рассматривать как позже отделившуюся группу. К этому классу относятся одноклеточные или колониальные формы со жгутиками или без них, а также нитчатые (трихальные и сифональные) виды. Клеточная стенка у монадных видов состоит из гликопротеинов, у неподвижных представителей — из полисахаридов, в том числе из целлюлозы. При клеточном делении новые поперечные перегородки возникают в фикопластах (см. введение к Сhlorophytа); такие стенки часто перфорированы плазмодесмами. Расположение жгутиковотносится к крестообразному типу; жгутиковый аппарат показывает расположение 1 — 7 или 12 — 6 (базальных телец и т. д.; см. рис. 11.88). Виды этого класса обитают преимущественно в пресных водах, незначительная часть их — в солоноватых и морских водах или даже во вневодных условиях.
1. Порядок: Volvocales. К этому порядку относятся имеющие жгутики одноклеточные формы, которые могут соединяться в колонии (см. 5.1). В этом порядке хорошо
прослеживается переход от одноклеточных форм к колониям с различным уровнем дифференциации и увеличивающейся полярностью. Радиально-симметричные клетки имеют 2, 4 или 8 апикальных «гладких» жгутиков равной длины (ср. рис. 11.97, D). Они отходят по обе стороны апикального сосочка.
Размножение одноклеточных видов осуществляется бесполым путем, зооспорами, которые образуются при повторных последовательных продольных делениях содержимого материнской клетки на 2 — 16 протопластов (рис. 11.94, В) и освобождаются при разрыве стенки материнской клетки, ставшей таким образом спороцистой. При половом размножении (у Сhlamydomonas 10% видов) сливаются двухжгутиковые гаметы или яйцеклетки и сперматозоиды.
При изогамии (рис. 11.94, С) копулирующие гаметы имеют полностью одинаковые размер, облик и движение и в целом не отличаются от вегетативных клеток. Они могут при соответствующих обстоятельствах либо произвольно копулировать друг с другом, либо развиваться вегетативно (факультативное определение функции). Таким образом, здесь мы имеем дело с зачатками сексуальности. Гаметы могут при этом относиться к одному типу скрещивания (однодомность) или же при видимом сходстве различаться в генетическом отношении (двудомность с «+» или «-» гаметами, например, Сhlamydomonas геihardiil). Определение функции репродуктивных клеток отчасти зависит от внешних условий. Богатая азотом среда (ионы NН4!) обусловливает исключительно образование вегетативных клеток. Ионы Са способствуют определению функции гамет.
Рис. 11.94. Chlorophyceae, Volvocales. Chlamydomonadaceae:
А — Chlamydomonas angulosa (1 100х); В — то же, четыре дочерние клетки внутри материнской клетки (1 100х); С, D — Chlamydomonas botryoides, копуляция двух изогамет (250х); Е — Chlamydomonas paradoxa, зигота (500х); F — Chlamydomonas monoica покоящаяся цистозигота (500х); G — Stepha- nosphaera pluvialis, прорастающая гипнозигота (300х); Н, J — Chlamydomonas braunli, копуляция анизогамет (400х); К — Haematococcus pluvialis (клетка, окруженная толстым слизистым слоем, 330х): с — хлоропласт; g — жгутик; к — ядро; р — пиреноид; s — глазок; v — пульсирующая вакуоль
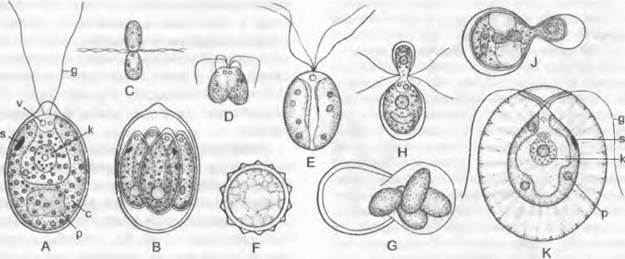
У видов с анизогамией (рис. 11.94, Н, J) ♂ гаметы меньшего размера копулируют с более крупными ♀ гаметами. У Сhlamydomonas suboogama жгутики ♀ гамет не функционируют, и это можно рассматривать как переход к следующей группе видов.
Оогамия. У Сhlorogonium oogamun ♀ гаметы полностью лишены жгутиков. Они амебоидно выходят из материнской клетки (рис. 11.95, D) и становятся яйцеклетками. Яйцеклетка оплодотворяется сперматозоидами, которые возникают в количестве 64 или 128 в виде светло-зеленых двухжгутиковых игловидных образований в ♂ особях путем последовательных делений (рис. 11.95, В). У Сhlamydomonas coccifera размножение осуществляется как гамето-гаметангиогамия, так как вся ♀ клетка, теряя жгутики, становится оогонием и оплодотворяется сперматозоидом.
Рис. 11.95. Chlorophyceae, Volvocales: А — Е — Chlorogonium оодаmum (240х): А — вегетативная клетка, В — ♂ клетка со сперматозоидами, С — ♀ клетка с яйцеклеткой, D — выход яйцеклетки наружу, Е — яйцеклетка, окруженная сперматозоидами; F — Stephanosphaera pluvialis (250х); G — Pandorina morum (160х); Н — то же, образование дочерних колоний (стенки материнских клеток частично уже разрушены, 150х)
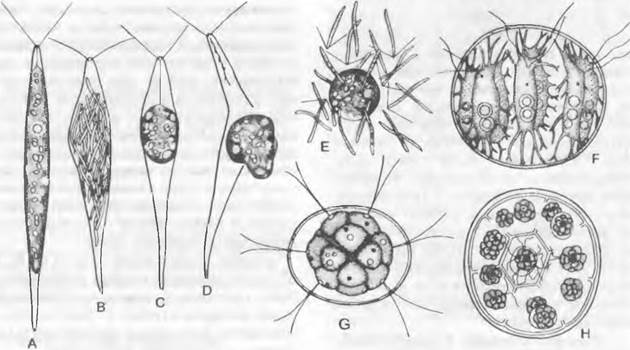
Таким образом, уже у этих одноклеточных форм можно проследить прогрессивное развитие от изогамии через анизогамию и оогамию до слияния ♂ гамет с оогонием.
Имеющие жгутики репродуктивные клетки возникают чаще всего в большом количестве (2 — 64) в материнской клетке путем повторных продольных делений. При изо- и анизогамии они попарно объединяются в зиготы (рис. 11.94, С — Е), причем чаще всего у партнеров соприкасаются сначала вершины жгугиков, которые скручиваются друг с другом (С). При копуляции в качестве гамонов действуют гликопротеины (см. 8.2.1.1), которые привлекают друг к другу гаметы разных типов скрещивания и обеспечивают временное склеивание жгутиков. Зигота имеет 4 Жгутика и поначалу еще подвижна (планозигота). Позже жгутики втягиваются, а становящаяся толстостенной зигота может переходить в состояние покоя (цистозигота; F). Гаметы всегда закладываются голыми, но могут также окружаться стенкой, и тогда при копуляции содержимое должно выходить через эту стенку. При прорастании зиготы (G) происходит мейоз, причем возникающие подвижные клетки распределяются в соотношении 1:1 по двум типам скрещивания (+ и -). Таким образом, подвижные клетки являются мейозооспорами, смена ядерных фаз зиготическая, а жизненный цикл гаплонтный. Для образования гамет в отдельных случаях задействованы целые особи.
Встречаемость. Volvocales — широко распространенные в пресных водах планктонные организмы. Они могут встречаться в таких больших количествах, что вода кажется совершенно зеленой; в морях Volvocales отсутствуют.
У многих видов развитию способствует поглощение органических веществ (миксотрофность; см. 9.1), поэтому они встречаются отчасти в водоемах с органическим загрязнением. Немногие виды (например, Polytoma uvella) живут чисто сапротрофно. Хотя у них хлорофилл отсутствует, имевшийся у предковых форм хлоропласт еще можно различить и здесь как бесцветную пластиду. Вместо тилакоидов она содержит систему беспорядочно расположенных трубочек. Аналогичные пластиды отмечаются у полученных в результате облучения ультрафиолетом желтых, нефотосинтезирующих мутантов Chlamydomonas.
Классификация Volvocales. Исключительно голые представители имеются в небольшом, возможно, исходном семействе Polyblepharidaceae. Если Polyblepharides, насколько известно, размножается только путем продольного деления на две клетки, то у более высокоразвитых форм происходит также половое размножение с фенотипическим или генотипическим определением типа скрещивания (+ и -). Dunaliella salina относится к последней группе, живет в очень соленых водах и окрашена каротиноидами в красный цвет.
Семейство Chlamydomonadaceae отличается от Polyblepharidaceae наличием клеточной стенки. Исходно хлоропласт занимает центральное положение, у большинства видов Chlamydomonas он постенный, у более высокоорганизованных форм сетевидно-продырявленный или даже распадающийся на отдельные мелкие диски. Половое размножение претерпевает прогрессивную эволюцию вплоть до оогамии.
Скорость движения имеющих жгутики клеток, например, у Chlamydomonas, составляет при фототаксисе примерно 10-кратную длину клетки в секунду. Недалеко от выроста, где находятся жгутики, расположены две пульсирующие вакуоли, которые попеременно сокращаются и выталкивают воду. Они поддерживают постоянство осмотического давления клетки. Каждая клетка содержит один бокаловидный хлоропласт, в «донышке» которого обычно находится пиреноид, вокруг которого откладывается крахмал (см. 2.2.9.1 и рис. 11.94, А, ср. 11.96, В), а на переднем конце — глазок (стигма, см. рис. 11.94 А). Образование крахмала в хлоропластах не связано исключительно с пиреноидом. Составляющие глазок пигментные шарики (каротиновые глобулы) вместе образуют 3 — 8 рядов. В образовании клеточной стенки (если она имеется; как, например, у Chlamydomonas) участвуют гликопротеиды (в том числе гидроксипролин и арабиноза, связанная с галактозой) и полисахариды (но не целлюлоза!).
Рис. 11.96. Chlorophyceae, Volvocales, Volvox: А — особь с 6 дочерними особями (50х); В — отдельная клетка с идущими от нее вбок к соседним клеткам плазмодесмами (1000х); С — соединение клеток, общий вид (500х); D — J — развитие и выворачивание наизнанку дочернего шара (D — 250х, Е — F — 350х, G — J — 250х); К — часть однодомной особи с 5 яйцеклетками и 2 пластинками сперматозоидов (200х); L — яйцеклетка, окруженная сперматозоидами (265х); М — сперматозоид (1000х); A — J, М — V. aureus; К, L — V. globator
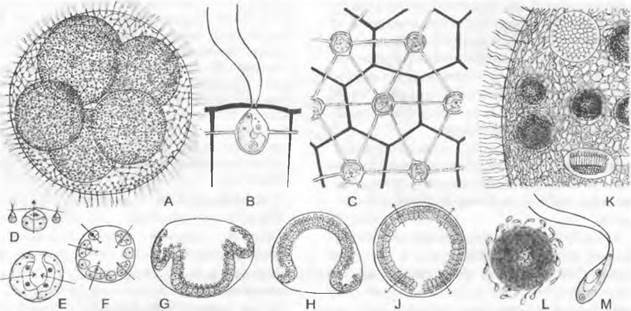
Некоторые виды (Haemalococcus pluvialis, см. рис. 11.94, К) окрашивают лужи после дождя в красный цвет из-за наличия у них каротиноидов. Chlamydomonas nivalis вызывает «красный снег» в высокогорьях и в Арктике. Некоторые Chlamydomonadaceae (и другие жгутиковые) поселяются даже зимой на голом льду и снегу (см. бокс 11.7, с. 199). Виды рода Cartería имеют 4 жгутика.
Семейство Volvocaceae представляет собой результат дальнейшего эволюционного развития по пути образования колоний. Отдельные клетки, часто похожие на Chlamydomonas,оказываются связанными между собой слизью или также плазмодесмами. У Oltmansiella 4 клетки, у Gonium 4 — 16 клеток объединены между собой в плоские таблички, причем все жгутики направлены в одну сторону. Колонии видов Stephanosphaera, живущих в остающихся после дождя лужах (см. рис. 11.95, F), представляют собой венец из 4, 8 или 16 клеток с жесткими придатками; хлоропласты имеют чаще всего по 2 пиреноида. У Pandorina 16 клеток, напоминающих Chlamydomonas, а у Eudorina и Pleodorina соответственно 32 или 128 таких клеток объединены в один шар, полый внутри. У всех колоний жгутики совершают синхронные движения, что обеспечивается плазмодесмами (см. 2.2.7.3). От Раndorina через Еudorina до Pleodorina происходит полярная дифференциация признаков относительно направления движения в воде (размер глазков, размер клеток, способность к размножению и т. д.). Отдельные клетки в конце индивидуального развития не отмирают, а делятся или используются для образования репродуктивных клеток. Наибольшая степень организации, которая оценивается по числу клеток, их дифференцировке и полярности, отмечена у Volvox (рис. 11.96). Колонии Volvox представляют собой видимый невооруженным глазом полый шар размером до нескольких миллиметров, заполненный внутри слизью. Шар образован многочисленными (до нескольких тысяч) клетками; например, у V. globator их насчитывается до 16 000. Каждая клетка имеет 2 жгутика, глазок и хлоропласт. Клетки соединены друг с другом широкими плазмодесмами (В; С). Способны к размножению только некоторые клетки, рассеянно расположенные в задней части шара. Большинство клеток служит для фотосинтеза и движения; однако и они различаются размером стигмы, который постепенно уменьшается при увеличении размера клеток, от переднего к заднему полюсу (полярность!). Передний полюс шара, кроме того, определяется направлением его перемещения в воде. Шар Volvox можно рассматривать уже не как колонию, а как многоклеточную особь. Отдельные клетки уже не тотипотентны. Поскольку к размножению способна только часть клеток, большинство клеток шара отмирает после образования дочерних шаров или гамет («труп» как результат многоклеточности).
При бесполом размножении у Volvox (рис. 11.96, D — J) отдельные, относительно крупные клетки (D) у заднего полюса колонии несколько раз продольно делятся и образуют полую сферу (F), которая в конечном счете превращается в открытый полый шар (G). Возникший таким образом дочерний шар выворачивается наизнанку (Н) и опускается со своими жгутиками, теперь уже ориентированными наружу, внутрь еще заполненного слизью материнского шара. Таким путем возникает несколько дочерних шаров (А), которые освобождаются только после разрушения материнской особи.
Половое размножение у Eudorina и Volvox оогамное. Внутри отдельных более крупных (генеративных) клеток возникают либо зеленые яйцеклетки (по одной в клетке, всего 6 — 8), либо многочисленные мелкие, при освобождении расположенные в виде пластинки, сперматозоиды (рис. 11.96, К, М). Volvoxglobator однодомный вид, V. aureus и V. carteri — двудомные. Развитие возникающих бесполым путем шаров в ♂ или ♀ особи у двудомных видов индуцируется половым гормоном (гликопротеидом). Он образуется ♂ особями (или же их сперматозоидами) и необходим для того, чтобы молодые шары, детерминированные как ♂ или ♀, развивались дальше в особи определенного пола. Если половой гормон отсутствует, то образуются только бесполые шары Volvox. После оплодотворения яйцеклетка превращается в толстостенную покоящуюся зиготу, при прорастании которой совершается мейоз. У всех Volvocaceae, таким образом, клетки колонии всегда происходят от одной-единственной изначальной клетки.
К описанным выше формам примыкают неподвижные одноклеточные или образующие колонии зеленые водоросли с отчасти еще выраженными признаками монадных водорослей, такими, как пульсирующие вакуоли, глазки, жгутиковые стадии. Их иногда рассматривают как отдельный порядок Tetrasporales.
2. Порядок: Chlorococcales (= Protococcales). Клетки с всего одним ядром и одним хлоропластом не имеют в вегетативном состоянии жгутиков и, следовательно, неподвижны. Только при размножении могут возникать двухжгутиковые подвижные клетки (зооспоры, рис. 11.97, D; или гаметы). Последние чаще всего бывают голые и только после плавания в течение некоторого времени одеваются стенкой (инцистируются). У некоторых представителей образуются только лишенные жгутиков «апланоспоры» (К). Половое размножение, известное у очень немногих представителей. изогамное. с помощью имеющих жгутики гамет (например, Pediastrum и Нуdrodictyon', оогамия встречается крайне редко. Зиготы прорастают с редукционным делением, так что жизненный цикл протекает исключительно в гаплофазе. У некоторых видов на основе одноклеточных форм образуются характерного облика колонии (см. 5.1; например, Реdiastrum, рис. 11.98; Scenedesmus, см. рис. 11.97, D). Химизм полисахаридной клеточной стенки в значительной мере неизвестен; у Рediastrum в ней встречается кремниевая кислота, а у некоторых видов откладывается спорополленин.
Рис. 11.97. Chlorophyceae, Chlorococcales. А — G — Chlorococcum (1000х): А — вегетативная клетка с чашевидным хлоропластом (с), имеющим небольшую выемку лишь на переднем конце, с пиреноидом (р), к — слегка просвечивающее ядро, В — деление на 8 дочерних клеток, С — выход зооспор в пузыре, позже расплывающемся, из внутреннего слоя мембраны материнской клетки (m), D — свободно плавающая зооспора с апикальными жгутиками равной длины, Е — она же, перешедшая в состояние покоя; глазок и жгутики еще присутствуют, F, G — развитие до стадии А при потере глазка (а) и вакуолей (v); Н — К — Chlorella, С. vulgaris (500х): Н — вегетативная клетка, J, К — деление на 8 апланоспор; L, М — Scenedesmus, S. acutus (1000х): L — 4-кпеточная колония, М — деление; N — R — Hydrodictyon, Н. utriculatum. N — молодая сетка в клетке материнской сетки (15х), О — ячейка молодой сетки (80х), Р — часть более старой клетки с зооспорами, Q, R — соединение зооспор в новую сетку в постенном протопласте (Р — R — 10х)
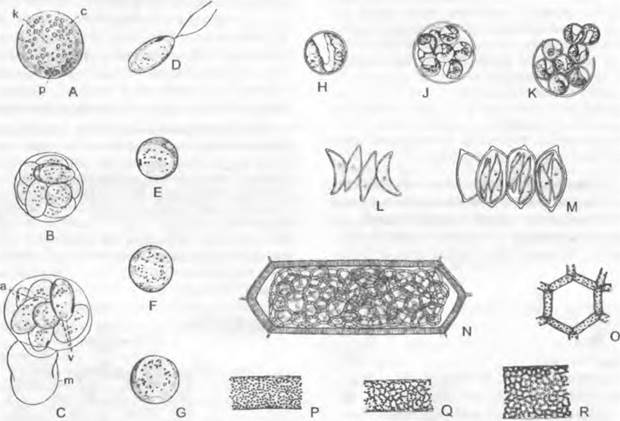
При клеточном делении часто (например, у Сhlorococcum) сначала образуется некоторое число голых дочерних клеток, которые в дальнейшем одновременно одеваются клеточными стенками. Kirchneriella, более детально изученная с помощью электронного микроскопа, отклоняется от этого варианта: септы с материалом клеточной стенки возникают сразу же после делений (последовательно), однако вскоре опять исчезают. Образующиеся 4 дочерние клетки отходят друг от друга и окружаются каждая собственной новой клеточной стенкой, прежде чем они по отдельности покинут материнскую клетку.
Как и у Volvocales, здесь мы встречаем прогрессирующий ряд, но он направлен от одноклеточных видов к колониям (см. 5.1), которые сгруппированы в виде табличек или полых шаров.
Правда, онтогенез Кirchneriella (см. выше) позволяет предположить, что одноклеточные формы этого рода возникли от многоклеточных (колониальных) структур.
Одноклеточные формы, от округлых до эллипсоидных, представлены видами таких родов, как Сhlorococcum (с зооспорами; рис. 11.97, D) и Ооcystis. Группировки расположенных в один ряд клеток простейших форм в количестве чаще всего по 4 (или 8) представлены у широко распространенных в пресных водах видов рода Scenedesmus (рис. 11.97, L, М). Более сложно устроены колонии у также часто встречающегося в пресных водах рода Рediastrum: они имеют вид изящных плоских табличек (рис. 11.98, А), свободно парящих в воде, и несколько напоминают Gonium, только без жгутиков. Взрослая колония Соelastrum трехмерная: клетки образуют полый шар (Е). У водяной сеточки (Hydrodictyon reticulatum), свободно парящей в пресной воде, цилиндрические клетки смыкаются своими концами по 3 — 4 и образуют мешковидную колонию в форме вытянутой замкнутой сети из многих ячеек, полую внутри (см. рис. 11.97, N).
Рис. 11.98. Chlorophyceae, Chlorococcales: А — D — Pediastrum, Р. granulaturrr. А — дисковидная колония из клеток, в трех из которых происходит деление; из четвертой клетки выходит пузырь с 16 подвижными клетками, В — подвижные зооспоры в вышедшем наружу пузыре, С — спустя 4,5 ч началась группировка 16 дочерних особей в колонию, D — то же, вид сбоку (300х); Е — Coelastrum, С. proboscideum (550х)
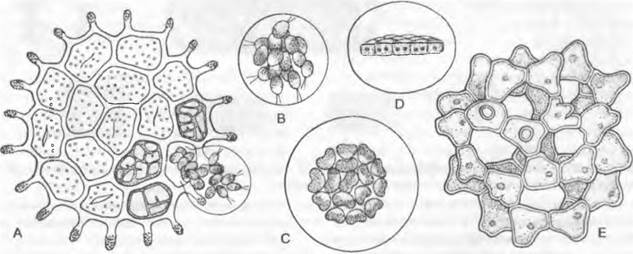
Половое размножение осуществляется изогаметами более мелкими, чем зооспоры. При прорастании зиготы сначала возникают 4 мейо- зооспоры, которые, недолго поплавав, превращаются в толстостенные «полиэдры». Только последние прорастают затем в новые колонии Нуdrodictyon, сначала значительно меньшего размера, чем взрослые. При бесполом размножении у видов всех этих родов образуются подвижные зоо- или неподвижные апланоспоры, которые, однако, освобождаются не поодиночке. Очень рано за счет слипания стенок они связываются между собой, формируя характерную для данного вида (по числу клеток и внешнему виду) колонию (рис. 11.97; 11.98). Объединение может происходить вскоре после выхода из материнской клетки в слизистом пузыре (рис. 11.98, А) или даже в самой материнской клетке, так что после ее разрушения уже имеется колония из соответствующего числа клеток, хотя сначала еше маленькая. Дальнейшие клеточные деления в колониях не происходят (за исключением образования репродуктивных клеток) Вышеупомянутое сходство с соответствующим эволюционным рядом у Volvocales состоит, таким образом, только во внешнем виде, а не в характере развития. У Volvocales колонии возникают путем повторного продольного деления образующих их клеток, за счет чего положение каждой клетки в колонии определяется с самого начала. У Сhlorococcales все клетки «выводка», возникшего в результате деления исходного протопласта, могут сначала (внутри клеток или же слизистых пузырей) свободно перемещаться относительно друг друга, прежде чем вторично соединяться между собой (рис. 11.98, А, В).
Встречаемость. Сhlorococcales обитают преимущественно в пресноводном планктоне; некоторые формы перешли к наземной жизни. Такие виды — обитатели влажной почвы и скал. Почвенная водоросль Spongiochloris устойчива к высоким температурам. Сhlorococcales наряду с другими водорослями являются также обычнейшими компонентами в зеленых налетах на коре деревьев и стенах. Некоторые Сhlorococcales — симбионты в лишайниках (однако ср. Тгеbouxia, Тгеbouxiophусеае). Scenedesmus, Ankislrodesmus и Нуdrodictyonчасто используются в чистой культуре для физиологических экспериментов.
В ископаемом виде формы типа современного Рediastrum описаны уже из перми. Напоминающие Сhlorococcales формы (Сагуоsphaerioides) относятся к древнейшим находкам эукариотных клеток (см. 11.3.2).
3. Порядок; Chaetophorales. Таллом водорослей, относящихся к этому порядку, представлен разветвленными нитями, состоящими из одноядерных клеток с одним хлоропластом. Он обычно гетеротрихальный, т. е. состоит из двух частей: «подошвы» из разветвленных нитей, которые стелются по субстрату, и более или менее обильно разветвленных прямостоячих нитей, несущих органы размножения (рис. 11.99, А). Половое размножение у тех видов, у которых оно было установлено, — изо-, анизо- или оогамия.
У видов некоторых родов гетеротрихальное строение четко не выражено из-за того, что одна из двух частей таллома выражена слабо или вообще практически отсутствует. Некоторые формы представляют собой объединения нескольких особей в колонии, находящиеся в общей слизи. У Stigeoclonium (рис. 11.99, А) наряду с четырехжгутиковыми зооспорами (В) встречаются двухжгутиковые изогаметы. У обитающей на почве водоросли Fritschiella (Индия, Африка; рис. 11.99, С) от подземных ползучих нитей поднимаются в воздушное пространство вертикальные разветвленные нити. Здесь проявляется функциональная дифференциация, сильно выраженная у высших растений: одна часть таллома, выполняет преимущественно поглощающую функцию, другая — ассимиляционную.
Рис. 11.99. Chlorophyceae, Chaetophorales: А — Stigeoclonium tenue (4х); В — Stigeoclonium subspinosum, зооспора (900х); С — Fritschiella tuberosa', а — глазок; b — поверхность почвы; р — пиреноид; ра — первичная вертикальная нить; рг — подземная ползучая нить; г — ризоид; s — подошва (стелющиеся по субстрату нити); sf — вторичные вертикальные нити; v — пульсирующие вакуоли; w — расположенные в толще воды вертикальные нити
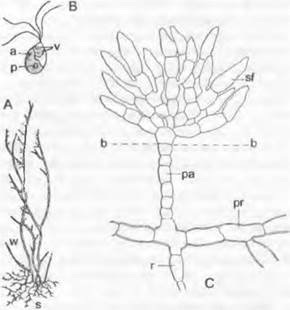
Встречаемость. Большинство видов (часто как эпифиты на водорослях и других водных растениях) — обитатели пресных вод (например, Сhaetophora, Sigeoclonium).
4. Порядок: Оedogoniales. Это еще один порядок с нитчатой организацией таллома. Хотя нити здесь чаще всего неразветвленные, но оогамное размножение, а также уникальная форма деления и растяжения клеток говорят о том, что это сильно подвинутые формы, прошедшие самостоятельный путь развития. Одноядерные клетки содержат по одному постенному сетчатому хлоропласту с многочисленными пиреноидами (рис. 11.100, А).
Уникальные деление и растяжение клеток связаны с образованием «колпачков» у верхнего конца клетки (рис. 11.100, J). Их возникновение сопряжено с началом деления ядра (профазой), когда у верхнего конца клетки образуется кольцевидный валик из сливающихся пузырьков (Гольджи?); он состоит в основном из аморфной, растяжимой фракции клеточной стенки. После завершения клеточного деления между дочерними ядрами внутри фикопласта (см. Chlorophyta, введение) появляется септа, дающая начало клеточной пластинке, сначала еще не фиксированной, — будущей поперечной перегородке. В области верхнего кольцевого валика внешняя стенка клетки в дальнейшем разрывается — образуется кольцевая трещина, после чего кольцевой валик растягивается в цилиндр. У места разрыва остается характерный колпачок. После повторений этого процесса у верхнего конца клетки число колпачков увеличивается, и они как бы вставлены друг в друга (рис. 11.100, С).
Жизненный цикл гаплонтный. Относительно крупные зооспоры возникают по одной, из всего содержимого клетки. Они имеют вблизи переднего конца, лишенного хлоропластов, характерный венец из многочисленных, не сгруппированных попарно жгутиков (рис. 11.100, С). В других местах нити отдельные клетки вздуваются наподобие бочонков в оогонии: их содержимое превращается в одну крупную яйцеклетку (Е), которая остается внутри оогония. Некоторые участки той же или другой нити (модификационное определение пола) в клетках, остающихся относительно короткими, образуют сперматозоиды, чаще всего по два. Они похожи на зооспоры, но мельче и имеют желтоватый цвет.
Иной путь переноса мужских репродуктивных клеток осуществляется через так называемые андроспоры и карликовые мужские растеньица («мужчинки»). В клетках, которые напоминают вышеописанные ♂ гаметангии, вместо сперматозоидов образуются несколько более крупные андроспоры, привлекаемые к оогониям хемотаксисом. Они не могут непосредственно оплодотворять яйцеклетки, а садятся на них или в непосредственной близости от них и прорастают в маленькие, состоящие из немногих клеток растеньица — так называемые «мужчинки» (рис. 11.100, Е, F), верхние клетки которых затем становятся гаметангиями и производят сперматозоиды, способные к оплодотворению. Происходящее в то же время созревание оогониев, очевидно, определяется гормонами, которые выделяются «мужчинками». С другой стороны, сперматозоиды по типу хемотаксиса привлекаются оогониями, которые теперь окружены слизистыми обвертками. Через отверстие в оогонии сперматозоиды проникают к яйцеклетке и один из них сливается с ней. После этого в оого- нии развивается толстостенная красная гипнозигота. При прорастании (G) ее содержимое делится на 4 крупные гаплоидные мейозооспоры (зиготическая смена ядерных фаз), которые выходят наружу и образуют новые нити (D).
Рис. 11.100. Chlorophyceae, Oedogoniales, Oedogonium; А — часть нити (600х); В — D — О. concatenation, выход зооспоры и ее прорастание (300х); Е — G — О. dliatum (350х): Е, F — оплодотворение, G — прорастание зиготы; Н — L — образование колпачка при делении клетки (200х); а — карликовое мужское растеньице (наннандрий); с — хлоропласт; к— ядро: о — оогоний; р — пиреноид; z — зооспора с запасными веществами, скрывающими ядро (см. D)
Обзор Chlorophyta. У зеленых водорослей наблюдается большое морфологическое разнообразие. Переход от одноклеточных форм к многоклеточным талломам идет через ряд уровней организации. Он конвергентно происходит в разных классах, соответственно в разных эволюционных линиях. Это касается не только классов зеленых водорослей, но и проявляется как общий эволюционный принцип в различных отделах водорослей. «Одетая» только одной (отчасти модифицированной) плазмалеммой клетка (например, Роlyblepharides) у зеленых водорослей получила дальнейшее развитие; во всех более высокоразвитых таксонах клетки имеют более или менее толстые клеточные стенки. Благодаря им некоторые формы могут даже жить вне воды в качестве почвенных или аэрофильных водорослей.
Компоненты клеточной стенки еще более разнообразны: в качестве полисахаридного материала уже используется и целлюлоза. Как защитное вещество встречается иногда спорополленин (Тгеntepсеае, Сhlorococcales). Клеточное деление — разделение дочерних клеток поперечной перегородкой — проходит различные эволюционные стадии. В простых случаях вслед за делением ядра происходит разделение плазмы и еще внутри материнской клетки одновременное окружение всех частей клеточными стенками. Идущее от боковых стенок центростремительное разделение материнской клетки за счет врастания перегородки наподобие ирисовой диафрагмы между отодвигающимися друг от друга дочерними ядрами — еще один способ, который можно рассматривать как исходный. В более эволюционно подвинутых случаях в образовании новой клеточной стенки участвует фикопласт, очень редко — уже фрагмопласт (Тгеntepohliophусеае). Исходно бокаловидный хлоропласт в одних случаях разделяется в виде сети или на отдельные диски, в других случаях образуются крупные хлоропласты в виде пластин или лент.
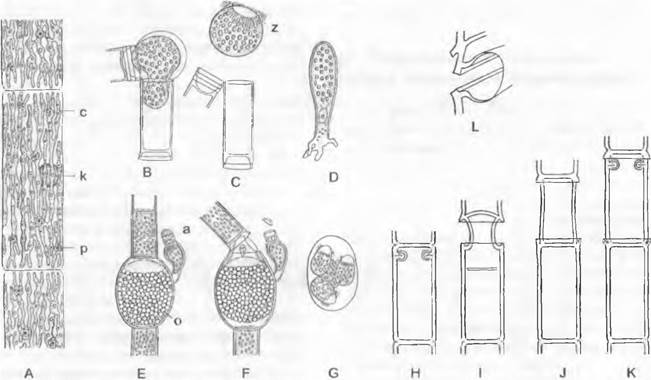
Половой процессэволюционирует от изогамии через анизогамию к примитивной оогамии и наконец к ее наивысшему уровню, когда яйцеклетка уже не выходит наружу, а оплодотворяется в оогонии. Некоторые зеленые водоросли помимо наличия и так уже неподвижных яйцеклеток утратили и жгутики у остальных репродуктивных клеток. Так, некоторые виды Рleurococcus (Аpatococcus, напротив, с зооспорами), приспособившись к жизни вне воды, распространяются лишенными жгутиков апланоспорами. В жизненном цикле можно проследить тенденцию к увеличению доли диплофазы (рис. 11.101). Обычно зеленые водоросли — гаплонты с зиготической сменой ядерных фаз; диплоидны в этом случае только зиготы (А). За счет смещения места мейоза (митотическое ядерное деление вместо мейоза) зигота прорастает в диплоидное вегетативное тело. За счет этого в жизненный цикл вклинивается дополнительно диплоидная фаза, которая заканчивается мейозом, сдвинутым во времени и в пространстве. Таким образом, возникает чередование гаплоидного гаметофита и диплоидного спорофита, т. е. гетерофазная смена поколений.
Рис. 11.101. Chlorophyta, смена поколений и ядерных фаз, схема основных типов: A — Ulothrix; В — Cladophora; С — Halicystis-Derbesia. Дикариотичная фаза — темный пунктир; диплоидная фаза — темные сплошные линии; гаплоидная фаза — светлые линии; G — гаметофит; S — спорофит; О — зигота; R! — редукционное деление
Смена поколений (рис. 11.101) может быть изоморфной (некоторые виды Clado- phora, В) или гетероморфной (с преобладанием спорофита, HalicystisDerbesia, С). Исключительно диплонтный цикл у зеленых водорослей пока неизвестен1. В целом смена поколений осуществляется на разных особях (диплобионтная). Гаплобионтная смена поколений на одной особи (что является закономерной ситуацией у мхов) у зеленых водорослей представляет редчайшее исключение (примеры — Prasiola stipitata, Bryopsis). Смену поколений ни в коей мере нельзя понимать, как закономерную последовательность разных фаз. За счет бесполого размножения каждое поколение может воспроизводить само себя независимо от смены поколений. Зеленые водоросли с простым жизненным циклом (Ulothrix, А) размножаются, как правило, вегетативным и бесполым путем (например, зооспорами), тогда как половое размножение наступает только при определенных внешних условиях.
1 В литературе обычно указывается, что чисто диплонтный цикл имеется у широко распространенного в Европе вида Сladophora glomerata, а также у многих (возможно, большинства) сифоновых зеленых водорослей. — Примеч. пер.
Chlorophyta — несомненно очень древняя группа низших растений. Однако с достоверностью могут быть идентифицированы только устойчивые вследствие обызвествления талломы морских Dasycladales, которые известны с кембрия. Поскольку Dasycladaceae уже в ордовике представлены большим разнообразием форм, они должны были возникнуть еще раньше. Из 120 родов, которые встречались на протяжении более 500 млн лет, в настоящее время сохранилось лишь 10.
• Отделы водорослей с комплексными пластидами, содержащими хлорофиллы а и b. Следующие два отдела, данные в соответствующих приложениях, содержат в своих клетках хлоропласты типа зеленых водорослей. Однако приобретение таких пластид осуществлялось путем вторичного эндоцитобиоза (см. 2.4).
1. Приложение к Chlorophyta: Chlorarachniophyta
Этот очень маленький отдел, в который входит всего два рода с одним видом в каждом1, представляет интерес тем, что их пластиды, окруженные четырьмя мембранами, как и у кринтофитов, содержат нуклеоморфу (см. 2.4). Последняя может рассматриваться как редуцированное ядро фотоавтотрофного эукариотного эндосимбионта. Виды этого отдела живут совместно с сифоновыми морскими водорослями теплых морей как голые амебоидные клетки, которые нитевидными плазматическими выростами соединены в сетевидные плазмодии. Светло-зеленые хлоропласты содержат хлорофиллы a и b. Тилакоиды в пластидах объединены в стопки по 2 — 6. Chlorarachniophyta считают произошедшими от нитчатых амеб путем приобретения пластид.
1 В настоящее времяв этом отделе насчитывается уже порядка 8 видов. — Примеч. пер.
2. Приложение к Chlorophyta: отдел эвгленовые (Euglenophyta)
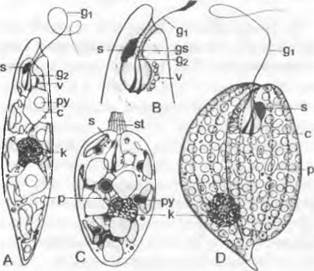
К этому отделу относятся одноклеточные формы, находящиеся на монадном уровне организации. При определенных условиях они иногда могут переходить в капсальное состояние. Размножение осуществляется путем продольного деления, половое размножение неизвестно. Зеленые хлоропласты содержат тот же набор пигментов, что и зеленые водоросли (хлорофиллы a и b, β-каротин, следы α-каротина), однако в них имеется один ксантофилл, неизвестный больше нигде в растительном царстве. В качестве запасного вещества наряду с фосфолипидами в пузырьках в плазме откладывается полисахарид парамилон в виде зернышек или дисков. Это глюкан с β-1,3- связями, который окрашивается йодом в синий цвет. Клетки нередко спирально закручены и имеют почти всегда простую, состоящую преимущественно из белков оболочку — пелликулу, которая непосредственно граничит с плазмалеммой (исключение представляет, например, Тгаchelomonas с домиком, содержащим железо). На переднем конце клетки имеется бутылковидное углубление — ампула, которая подразделяется на брюшную часть и канал. К ампуле прилегает пульсирующая вакуоль, которая окружена несколькими дополнительными пульсирующими вакуолями и служит как органелла, обеспечивающая осморегуляцию. У основания ампулы почти всегда отходят два жгутика, каждый от своего базального тела: один длинный и один короткий, не выходящий наружу из ампулы и срастающийся концом с длинным жгутиком. В месте их слияния находится светочувствительная органелла — фоторецептор. Вблизи ампулы расположен красный «глазок» (рис. 11.102, В). Он состоит из отдельных липидных глобул, каждая из которых окружена собственной элементарной мембраной (относительно роли «глазка» при фототаксисе см. 8.2.1.2). Длинный жгутик, снабженный волосками (см. рис. 11.20, С), описывает при движении поверхность конуса. При одновременном вращении вокруг продольной оси клетка, например, у Euglena, каждую секунду перемещается вперед на расстояние, равное 2 — 3 длинам ее тела.
Рис. 11.102. Euglenophyta: А — Euglena gracilis (600х); В — то же, передний конец (1000х); С — Colacium mucronatum (500х); D — Phacus triqueter (600х); с — хлоропласт; g1 — двигательный жгутик; g2 — второй жгутик; gs — расширение жгутика (фоторецептор); к — ядро; р — свободный парамилон; ру — пиреноид; s — глазок; st — слизистая ножка; v — пульсирующие вакуоли
Ультраструктура эвгленовых показывает следующие особенности. В интерфазном ядре можно видеть хромосомы в конденсированном состоянии. Хлоропласты имеют оболочку из трех мембран, которая никогда не бывает связана эндоплазматической сетью с ядерной оболочкой. В хлоропластах тилакоиды по большей части сгруппированы в ламеллы по три.
Эвгленовые насчитывают более 800 видов, относящихся примерно к 40 родам. В большинстве своем они живут в пресных водах. Виды рода Euglena встречаются в основном в стоячих водоемах, богатых органическими веществами Виды Phacus (рис. 11.102, D), наоборот, предпочитают воды, бедные питательными веществами. Colacium (С) прикрепляется с помощью слизистой ножки к свободно плавающим мелким организмам; только при размножении возможно его свободное движение с помощью жгутиков.
Хотя большинство видов питается фотоавтотрофно, у них имеется тенденция к поглощению органических веществ дополнительно к продуктам фотосинтеза. Ряд бесцветных форм питается полностью гетеротрофно; у некоторых (например. Регаnеmа) есть воронковидное углубление (цитостом), где находятся ловчий аппарат и палочковидный ловчий орган, с помощью которых они способны захватывать такие микроорганизмы, как бактерии, водоросли или клетки дрожжей. Таким образом, границы между растительной и животной организацией здесь еще размыты.
Euglena gracilis в тем новых культурах теряет весь хлорофилл и все тилакоиды. Остающиеся тельца напоминают пропластиды; они сохраняют способность к делению и вовремя темновой фазы, чем поддерживается сохранность пластидома. При последующем освещении сохранившиеся бесцветные пластиды превращаются в хлоропласты с тилакоидами и возобновляется фотосинтез. Наряду с этим имеются разнообразные варианты данного вида, которые вообще не имеют хлоропластов. Такие формы, получаемые при определенных условиях (например, при очень быстрой последовательности делений), уже никогда не могут образовывать хлоропластов.
Эвгленовые были сначала гетеротрофной, вероятно, очень рано обособившейся группой, которая сравнительно поздно приобрела хлоропласты путем эндоцитобиоза с зелеными водорослями.