БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
Второй отдел: Streptophyta
В этом отделе жгутики либо отсутствуют (Zygnematophyceae), либо, если имеются, занимают боковое положение (KJebsor- midiophyceae; Charophyceae). Новые клеточные стенки при делении клеток (так же как у рассмотренных Trentepohliophyceae и сосудистых растений) закладываются во фрагмопластах (микротрубочки располагаются перпендикулярно плоскости будущей перегородки; ср. фикопласт: Chlorophyta, введение). Объем этого отдела, как и его разграничение с уже обсуждавшимися Chlorophyta, подтверждается данными анализа ДНК.
Первый подотдел: Streptophytina (зеленые водоросли II)
В этом подотделе объединены все группы Streptophyta от одноклеточных до многоклеточных водорослей, имеющие простую организацию. Наиболее развитые формы, связанные с водной средой, достигают довольно высокой функциональной дифференциации; у них формируются ткани (например, узлы Characeae) и защищенные органы размножения.
1. Класс: Mesostigmatophyceae
Это монадные одноклеточные организмы (например, Mesostigma) пресных вод. Их клеточный покров выглядит как орнаментированный мелкими точками. Он состоит из трех слоев чешуек. Чешуйки самого внешнего слоя (из органического материала) крупнее чешуек внутренних слоев и имеют вид корзиночек. Два жгутика несколько неравной длины прикреплены не терминально. Chlorokybus нередко образует правильной формы группы из нескольких (до 32) клеток. Из-за наличия чешуек представители этого класса раньше относили к Prasinophyceae (отдел Chlorophyta); принадлежность их к Streptophytina обоснована анализом ДНК. Создается впечатление, что Mesostigmatophyceae — предковая группа, от которой берет начало эволюция зеленых наземных растений.
2. Класс: конъюгаты, или сцеплянки (Zygnematophyceae, Conjugatae)
Конъюгаты не образуют жгутиковых стадий: у них нет ни зооспор, ни гамет со жгутиками. Половое размножение осуществляется путем конъюгации1: два голых протопласта, каждый из которых представляет все содержимое своей клетки, сливаются в зиготу. Последняя прорастает после периода покоя, претерпевая мейоз; смена ядерных фаз зиготическая. Следовательно, конъюгаты — чистые гаплонты, представленные коккоидными или нитчатыми формами. Нити неразветвленные и легко распадаются на отдельные клетки. Каждая клетка содержит ядро, расположенное в ее середине. К конъюгатам относится примерно 4000 — 6000 видов (около 50 родов), обитающих в бентосе и отчасти в планктоне, почти исключительно в пресных водах.
1 Следует помнить, что термин «конъюгация» в биологии очень неудачно используется для самых разных явлений (для инфузорий, бактерий, хромосом и в данном случае). — Примеч. пер.
Mesotaeniaceae — относительно примитивная группа. Водоросль существует либо в виде отдельных клеток, либо образует слизистые колонии (рис. 11.103, А; коккоидный уровень организации). Клеточная стенка сплошная, гладкая.
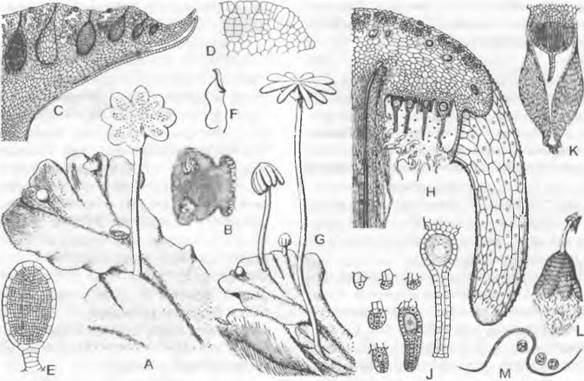
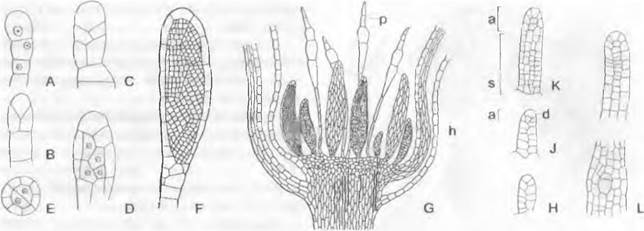
Рис. 11.103. Streptophytina, Zygnematophyceae; Mesotaeniaceae и Desmidiaceae:
А — Mesotaenium braunii (280х); В — Closterium moniliferum (200х); С — Closterium regulare, поперечный разрез через клетку, хлоропласт имеет звездчатый контур (200х); D, Е — Closterium parvulum, копуляция (300х); F — Closterium rostratum, выход зиготы из оболочки (200х); G — Closterium sp., деление зиготы (200х); Н — Cosmarium botrytis (280х); J, К — тоже, деление (280х); L — Micrasterias denticulate (125х); М, N — Oocardium stratum, вид сверху и продольный разрез (320х); О — Desmidium swartzii, часть цепочки из клеток; Р — то же, поперечный разрез через клетку (350х); dk — дегенерировавшее ядро; g — слизистая ножка; к — ядро; kh — известковая обвертка; р — пиреноид; v — вакуоль с кристалликами гипса
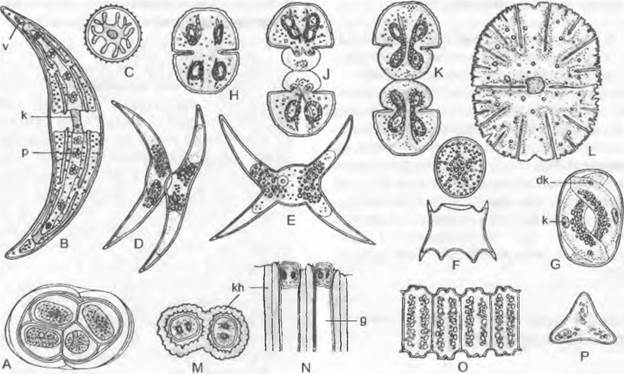
Desmidiaceae2 — как правило, одноклеточные формы (коккоидные). Чаще всего орнаментированные, нередко содержащие железо (поэтому желтоватые), клеточные стенкисостоят из двух равных половинок — полуклеток, которые отделяются друг от друга швом или перетяжкой (перешеек, isthmus). Внутри каждой из двух точно симметричных полуклеток содержится по одному крупному центральному (не постенному) хлоропласту с одним или несколькими пиреноидами (см. рис. 2.87; бокс 4.1, рис. С; рис. 11.103, В, С). В середине клетки находится ядро.
2 Выразительно немецкое название этой группы — Zieralgen (декоративные водоросли). — Примеч. пер.
Вегетативное размножение осуществляется путем деления клетки на две, причем (как у диатомей — см. Bacillariophyceae) у дочерних клеток должна быть восполнена половина клеточной стенки (рис. 11.103, J, К). При этом вновь возникают одноклеточные особи. Однако у представителей некоторых родов дочерние клетки остаются соединенными между собой, образуя цепочки.
При половом размножении две генотипически разные клетки прикладываются друг к другу (рис. 11.102, D) и окружаются слизью. Затем клеточная стенка расходится в середине, протопласты выходят в виде голых гамет в разбухающий, вскоре ослизняющийся копуляционный канал (мостик) и сливаются в зиготу (Е), стенка которой часто несет шипы. Рядом со зрелой гипнозиготой сначала находятся 4 пустые половинки клеточных стенок от обеих слившихся клеток. При прорастании зиготы из 4 гаплоидных ядер, возникших в результате мейоза, у большинства Desmidiaceae 2 погибают, так что возникают только 2 гаплоидных «проростка».
Desmidiaceae, относящиеся к красивейшим водорослям, очень разнообразны по своему облику. Их клетки, например, имеют форму полумесяца (Сlosterium, рис. 11.103, В), бисквита (Сosmarium, Н) или звезды (Мicrasterias, L). Еuastrum (см. бокс 11.7, с. 199) имеет на концах полуклеток выемку, Staurastrum (см. бокс 11.7)— полуклетки с острыми углами. На обоих концах клеток Сlosterium расположены вакуоли с кристалликами гипса, которые находятся в интенсивном броуновском движении (В). Некоторые Desmidiaceae выделяют через поры в мембранах слизистые тяжи, с помощью которых клетки могут медленно передвигаться. Особи видов рода Oocardium, обитающие в богатых известью ручьях, сидят на слизистой ножке, которая инкрустирована известью (М, N). Десмидиевые развиваются преимущественно в бедных питательными веществами водоемах с низким значением pH, например, в торфяных болотах, где имеется большое разнообразие их видов; Pleurotaenium и Staurastrum обитают также в водах с щелочной реакцией.
Семейство Zygnemataceae представлено неразветвленными нитчатыми (формами. Наиболее известен род Spirogyra (рис. 11.104, А). Его многочисленные виды часто встречаются весной в водоемах со стоячей или медленно текущей водой в виде неприкрепленной желтовато-зеленой «тины». Нити растут интеркалярно за счет растяжения в длину и поперечного деления всех клеток; таким образом, все клетки равноценны: отсутствует и полярность нитей. Гладкие, лишенные пор целлюлозные клеточные стенки, снаружи покрываются слизью, из-за чего нити слизистые на ощупь. При митозе ядерная мембрана по большей части сохраняется (внутриядерный митоз). Поперечная перегородка образуется центрипетально по типу ирисовой диафрагмы и дополнительно — как клеточная пластинка во фрагмопласте. Нити могут распадаться по поперечным перегородкам на одно- или многоклеточные фрагменты, которые служат для вегетативного размножения (см. 10. 1.3.3).
Ядро находится посередине клетки и подвешено на протоплазматических тяжах внутри крупной вакуоли. Хорошо различимы один или несколько прилегающих к стенке хлоропластов, всегда в виде левовращающей спирали (см. рис. 2.87; 11.104, А, С: с) с пиреноидами (А: р).
Рис. 11.104. Zygnematophyceae: А — Н — Spirogyra: А — S. jugalis, клетка (250х), В — S. quinina, анизогамная копуляция (240х), С — H — S. longata; С — участок хлоропласта у клеточной стенки (750х), D — Н — молодые и старые зиготы, D — два половых ядра перед копуляцией, Е — после слияния, F — деление ядра зиготы на 4 гаплоидных ядра, G — три мелких ядра дегенерируют (D — G — 250х), Н — одноядерный проросток (180х); J —L — Mougeotia: J, К — М. scalarís, хлоропласт повернут к зрителю широкой стороной («анфас») и узкой стороной («в профиль») (600х), L — М. calospora, изогамная копуляция (450х); с, с1, c2 — хлоропласт(ы); к — ядро; р — пиреноид; s — крахмал; w — клеточная стенка; z — зигота; zw — стенка зиготы
При половом размножении две нити, чаще всего морфологически неразличимые, располагаются параллельно. По линии соприкосновения из клеток выдвигаются сосочки, так что нити снова несколько отодвигаются друг от друга и получается фигура, похожая на лестницу (лестничная конъюгация; см. рис. 11.104, В). Сосочки за счет лизиса стенки в месте соприкосновения превращаются в копуляционный канал между соответствующими клетками («гаметангиями»). Каждая клетка нити может стать «гаметангием». Определение пола модификационное (♂ и ♀ нити). Протопласт из ♂ клетки в виде голой «мигрирующей гаметы» переходит в прилегающую ♀ клетку и сливается с ее протопластом («покоящаяся гамета»). Получающаяся зигота за счет отдачи воды уменьшается в объеме и превращается в гипнозиготу (рис. 11.104, В: z), которая окружается многослойной толстой коричневой стенкой, содержит много крахмала и масла и в таком виде переносит неблагоприятные условия. Хлоропласта (один или несколько) ♂ «гамет» погибают. При прорастании зиготы, связанном с мейозом, 3 ядра (F, G) дегенерируют, так что возникает только один гаплоидный проросток, который вытягивается в длину и, претерпевая клеточные деления, образует новую нить (Н).
Некоторые виды Spirogyra однодомны. У них сливаются протопласты соседних клеток одной и той же нити через боковые копуляционные мостики (боковая конъюгация).
Zygnema и Моugeotia отличаются строением хлоропластов. У Zygnema в каждой клетке имеется два звездчатых хлоропласта, у Mougeotia (рис. 11.104, J, К) — единственный хлоропласт в виде плоской осевой пластинки, реагирующей на свет (ср. 8.2.2). В обоих родах есть виды, у которых зигота образуется в копуляционном канале (L); этот процесс напоминает таковой у представителей предыдущего порядка.
Zygnematophyceae — группа, четко отличающаяся по типу размножения и строению клетки от других зеленых водорослей, от которых она, очевидно, рано отделилась в ходе эволюции, полностью утратив жгутиковые стадии.
3. Класс: Klebsormidiophyceae (Coleochaetophyceae)
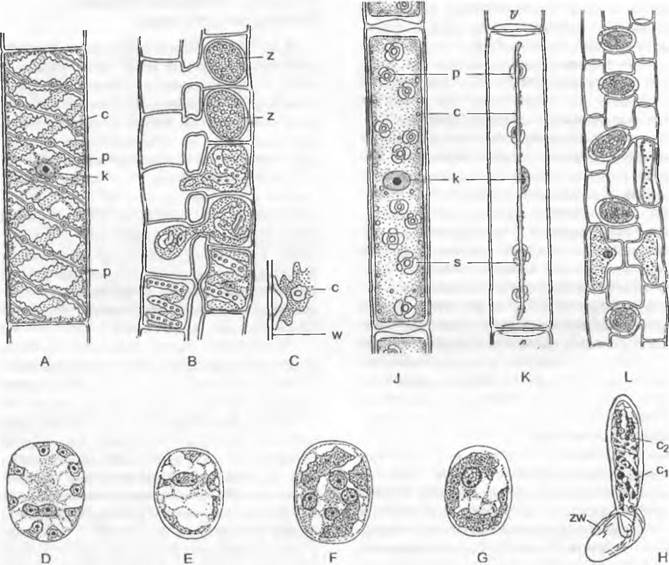
Клеточная стенка нитчатых (неветвящихся и разветвленных) представителей этого класса содержит целлюлозу (фибриллярное расположение). Жгутиковые репродуктивные клетки покрыты ромбовидными чешуйками из органического материала; этим они напоминают клетки Mesostigmatophyceae. Виды этого класса встречаются в пресных водах и во влажных местах вне воды. В порядке Coleochaetales виды представлены разветвленными нитями. Таллом их разделяется на подошву и вертикальные нити (гетеротрихальное строение; рис. 11.105; подобная дифференциация таллома у Trentepohliophyceae и Chaetophorales из Chtorophyceae).
Coleochaete имеет высокий уровень развития среди Chlorophyta (наличие дисковидной подошвы — рис. 11.105, А; по- особому дифференцированные волоски; оогамное размножение). Бутылевидный оогоний с бесцветной шейкой (С), которая открывается на вершине для проникновения бесцветных двухжгутиковых сперматозоидов. После оплодотворения округлая зигота увеличивается в размере и одновременно от несущей ее клетки и соседних клеток вырастают нити, так что в конечном счете зигота одевается однослойной плектенхимой — возникает «плодовое тело» (Е). При прорастании этого покоящегося органа не сразу возникают мейозооспоры, а сначала внутри зиготы после мейоза образуется гаплоидное тельце из 16 — 32 клеток, в каждой из которых развивается по одной гаплоидной зооспоре.
Klebsormidiophyceae названы по роду Klebsormidium, виды которого имеют коккоидное строение или представлены неразветвленными нитями.
Рис. 11.105. Streptophytina, Klebsormidiophyceae: А — Coleochaete scutata, подошва (80х); В — Aphanochaete repens, развитие одетой воротничком щетинки (250х); С — Е — Coleochaete pulvinata, С — оогоний незадолго до вскрытия; D — он же оплодотворенный; Е — зигота за счет обрастания нитями превращается в «плод» (500х); ек — ядро яйцеклетки; sk — ядро сперматозоида
Среди зеленых водорослей Coleochaetophyсеае вместе с еще не рассматривавшимися Charophyceae наиболее близки зеленым наземным растениям. Этот класс водорослей обладает такими общими признаками с мохообразными и папоротникообразными, как расположенные сбоку жгутики, тенденция к образованию тканей, защита зиготы (начало формирования зародыша), фрагмопласт и целлюлоза одного и того же типа. «Плодовые тела» Coleochaete защищены от высыхания и разрушения микробами спорополленином и соединениями типа лигнина.
4. Класс: харовые, или лучицы (Charophyceae)
Харовые представляют собой высокоразвитые зеленые водоросли с талломами, в которых строго последовательно чередуются тканеподобные и нитчатые участки
(соответственно узлы и междоузлия). Органы размножения достигают очень специфичной комплексности, отсутствующей у всех предыдущих групп. Харовые, представленные немногими родами, образуют в прудах и ручьях «подводные луга». Их известно около 300 видов. Обитают они в пресных и солоноватых водах; пресноводные виды часто особенно обильно развиваются в водоемах с высоким значением pH (pH 7 и более; жесткая вода). Characeae — единственное семейство класса, где еще сохранились ныне живущие формы.
Клеточные стенки часто инкрустированы известью, некоторые Characeae относятся к числу важнейших образователен известковых туфов. Они не переносят высоких концентраций фосфатов при загрязнении воды.
Строение. Главные и боковые оси имеют верхушечный рост, нарастая за счет одной апикальной клетки (см. рис. 5.9, В). Она отчленяет вниз от себя по очереди более узкие клетки (будущие узлы) и более вытянутые клетки (будущие междоузлия); последние больше не делятся и растягиваются в длину, претерпевая вакуолизацию, до нескольких сантиметров.
Харовые водоросли характеризуются, таким образом, регулярным подразделением таллома, достигающего нескольких дециметров, на узлы (nodi) и междоузлия (internodia).
Клетки узлов сохраняют способность к делению и развиваются в многоклеточные диски узлов, из которых вырастают расположенные мутовками членистые боковые оси разных порядков. Кроме того, возникают короткие, шиловидные клетки-«прилистники» и клетки коры. Коровые клетки (характерные для рода Chara и отсутствующие у видов рода Nitellaи др.) образуют прилегающий к междоузлиям покров из нитевидных клеток.
Боковые ветви показывают сходное с основной осью подразделение. Они не покрыты корой, простые или несут на своих узлах короткие боковые ветви второго порядка со сходным подразделением на узлы и междоузлия.
В каждой мутовке из «пазухи» коротких «побегов» может возникать длинный «побег», сходный с главной осью (см. рис. 5.9).
Основаниями растения прикрепляются к незакрепленному грунту (илу, песку) посредством бесцветных разветвленных нитевидных ризоидов, возникающих из узлов. Некоторые Сharaceae образуют из нижних частей осей клубеньки, плотно забитые крахмалом, являющиеся органом перезимовки.
Молодые клетки непосредственно после клеточного деления одноядерны. В длинных клетках междоузлий ядро растет эндомитотически и распадается на многочисленные ядерные фрагменты, так что эти клетки становятся многоядерными. Цитоплазма находится чаще всего в интенсивном движении (цитоплазматическое движение, см. 8.2.2). Многочисленные хлоропласта находятся в постенной цитоплазме клеток.
Твердая часть клеточной стенки состоит из целлюлозы, по тонкому строению сходной с целлюлозой высших растений. Новые поперечные перегородки клеток возникают во фрагмопластах.
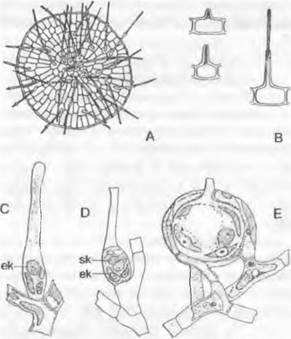
Размножение. Харовые — все оогамные гаплонты с зиготической сменой ядерных фаз. Расположенные вертикально оогонии Chara окружены спирально обвивающими их коровыми нитями. Мужские гаметы возникают в шаровидных вместилищах сложного строения (спермогонии Chara). Двухжгутиковые сперматозоиды закручены в виде штопора, тогда как у всех других зеленых водорослей они радиально-симметричные. Округлые, в зрелом состоянии окрашенные каротиноидами в желто-красный цвет спермогонии и яйцевидные зеленые оогонии (и те, и другие видны невооруженным глазом) образуются у узлов боковых осей.
Рис. 11.106. Streptophytina, Charophyceae: А — вид сбоку на часть таллома со спермогонием (s) и оогонием (о), коровыми нитями и коронкой (с) (50х); В — рукоятка с головкой и сперматогенными нитями; С — клетки сперматогенной нити, содержащие каждая по одному сперматозоиду; D — сперматозоид (540х); Е — продольный разрез через молодой спермогоний; g — жгутики; gz — рукоятка; к — спирально закрученное длинное ядро; kz — головка; р — плазма; w — стенка
Бокс 11.7. Встечаемость и образ жизни водорослей
Преобладающее большинство водорослей — фотоавтотрофы. Им противостоят миксотрофные и гетеротрофные формы. Миксотрофия позволяет фотосинтезирующим организмам дополнительно поглощать органику из богатой питательными веществами окружающей среды. Гетеротрофные водоросли утратили свои фотосинтетические пигменты и питаются готовыми органическими веществами; «фаготрофные» представители этой группы «поедают» твердые частицы пищи, которые поступают в пищеварительные вакуоли. Если фототрофные водоросли — типичные растения, то лишенные пигментов их фаготрофные представители имеют животный образ жизни. Среди монадных форм могут находиться близкородственные виды с автотрофной (растительной) и фаготрофной (животной) организацией. Границы между растениями и животными, таким образом, на этом относительно низком эволюционном уровне еще расплывчаты.
Про- и эукариотные водоросли хотя и встречаются почти во всех биотопах, но большинство видов обитает в воде, где они как «планктон» свободно парят в ее толще или как «бентос» прикрепляются к камням, песку и другому субстрату. Из-за различного содержания соли имеется два типа водного жизненного пространства: моря и пресные воды.
Морские водоросли
Растительный планктон морей представлен прежде всего диатомеями и динофитами (перидинеями), а также крошечными гаптофитами (Соccolithophorales) и золотистыми водорослями (силикофлагеллятами). Представители обеих последних групп не удерживаются ячейками планктонной сетки, и их можно получить только путем центрифугирования («нанопланктон»).
Наибольшая плотность планктона (до 100 000 клеток в литре воды) наблюдается в хорошо освещаемом слое воды. В литре поверхностной воды Атлантики вблизи Фарерских островов было выявлено: 32 000 клеток динофитов, 1 600 диатомей и 54 000 кокколи- тофорид. На глубине ниже 100 м число этих планктонных форм резко падает. Однако и на больших глубинах (4 000 — 5 000 м) еще встречаются Соссоlithophoraceae и «оливково-зеленые клетки», чье систематическое положение до сих пор неясно. Кроме того, наибольшая плотность планктона встречается в более холодных морях и в области холодных морских течений; это объясняется тем, что воды здесь богаче соединениями азота и фосфатами. Данные вещества потребляются в слоях воды, расположенных ближе к поверхности, и накапливаются в более глубоких слоях вследствие опускания туда мертвых клеток. В холодных областях за счет как зимнего, так и ночного охлаждения поверхности моря вода перемешивается лучше, чем в тропиках, что приводит к более интенсивному развитию планктона. Богатство планктона отмечается также там, где холодные глубинные воды, насыщенные соединениями азота и фосфатами, выходят на поверхность благодаря течениям.
Парение планктонных организмов в воде сопровождалось бы более или менее медленным их погружением, если бы не регулировалось специфическим весом и силой трения, а также активным движением жгутиков. Это объясняет многие особенности планктонных водорослей: наличие (образование и разложение) масла как запасного вещества, развитие выростов и выступающих клеточных стенок (см. рис. 11.70), группировка многих клеток в цепочки (см. рис. 11.76). Отмечено, что в теплых водоемах (где вязкость меньше) служащие для парения выросты крупнее, чем в холодных. Минеральные скелеты планктонных водорослей оседают на морское дно. Поскольку известь на глубине более 4000 — 5 000 м растворяется. на больших глубинах мы находим только скелеты диатомей, силикофлагеллят и из животных — радиолярий. На меньших глубинах (2 000 — 5 000 м) происходит также отложение извести (Соссоlithopогасеае, из животных — глобигерины и т. д.). За 1 000 лет откладывается слой мощностью всего 1.5 см. Растительный бентос в морях состоит (не считая морской травы Zosteегасеае) исключительно из водорослей, преимущественно бурых и красных. Чаще всего они с помощью присосок или выростов прикрепляются к твердому субстрату (камню) (см. рис. 11.83, 11.85). Подвижный субстрат (ил, песок) заселяется видами лишь немногих родов, например, Саulerpa (см. рис 11.91, А). Бентосные водоросли встречаются от зоны попадания морских брызг до глубин, где еще возможен фотосинтез (180 м).
В тропических морях водорослевая растительность не достигает такой пышности, как в морях умеренной и холодной зон (ср. причины, указанные для планктонных форм). Бурые водоросли представлены гораздо в меньшей степени, тогда как красные, напротив, достаточно широко, как и некоторые теплолюбивые зеленые водоросли: Caulerpaceae, Dasycladaceae, Codiaceae, Valoniaceae. Богата также растительность коралловых рифов, причем водоросли (Halimeda, см. рис. 11.91, В; Dasycladaceae, см. рис. 11.92, В; Lithothamnion) принимают в образовании извести большее участие, чем сами кораллы. Единственное в своем роде явление представляет собой «Саргассово море», где бурая водоросль Sargassum (см. рис. 11.85, А), плавающая на поверхности воды, образует обильные заросли (сгоняемые к одному месту растения имеют до 5 т фитомассы на квадратную морскую милю).
В умеренно-теплых морях, например, в Средиземном, бентос состоит преимущественно из красных и сравнительно мелких бурых водорослей. Вышеупомянутые Bryopsidophyceae и Dasycladophyceae представлены несколькими видами. Широко представлены виды Lithothamnion. Поскольку интенсивность света зависит от времени года, основное время развития водорослей у поверхности воды — весна, на глубине — лето, а осенью водоросли вообще развиваются меньше.
В холодно-умеренных морях, например, в Северном, явно доминируют бурые водоросли, как по размерам, так и по массе. Время года существенно влияет на многие виды. Так, Desmarestia теряет осенью ассимилирующие волоски, а красная водоросль Delesseria — нежные плоские части таллома, так что зимуют только «жилки».
Крупные ламинарии (см. рис. 11.83) ежегодно обновляют свои филлоиды. На рис. 11.107 на примере скалистого побережья Ла-Манша видна четко выраженная вертикальная зональность водорослевой растительности в зависимости от уровня воды при приливах и отливах. Виды верхних зон (Bangia, Potphyra, Fucus) выдерживают температуры до -20 °С, тогда как никогда не подвергающиеся высыханию обитатели более глубоких зон (Laminaria, Delesseria) вымирают уже при нескольких градусах ниже нуля. Холодные моря имеют бедный видовой состав водорослей, однако именно здесь талломы бурых водорослей достигают наибольших размеров: можно назвать хотя бы Macrocystis (см. рис. 11.83, Е), Lessonia (D) и Nereocystis (С) — все из Laminariales, а также Durvillea из Fucales. Они практически не уступают в размере крупным наземным растениям.
Рис. 11.107. Профиль растительности на побережье Ла-Манша: Chlorophyta: Prasiola, Urospora, Enteromorpha-, Rhodophyta: Bangia, Porphyre, Rhodymenia, известковые корки (например, Lithothamnion); Phaeophyceae: Peivetia, Fucus, Ascophyllum, Alaria, Laminaria, Halidrys, Cystoseira; лишайники: Verrucaria
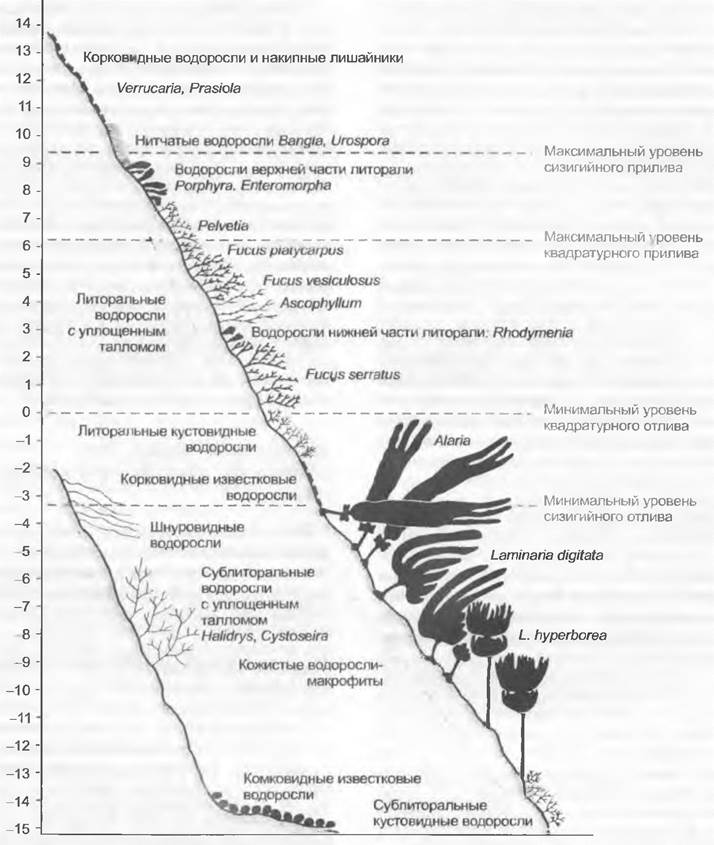
Распространение морских бентосных водорослей зависит от загрязнения и наличия питательных веществ. Например, Ulva растет в очень богатой питательными веществами воде, Padina — в воде с умеренным их содержанием, a Sargassum и Fucus — при малом содержании органики. Между морскими и пресными водами промежуточное положение занимают солоноватые воды. Здесь за счет регулярных приливов и отливов или прибоя перемешиваются пресная и соленая вода. К местообитаниям такого типа, со спецификой планктонных и бентосных водорослей (например. Chaгасеае), относятся также устья проточных вод.
Пресноводные водоросли
В пресной воде видовой состав растительного планктона в значительной степени зависит от наличия питательных веществ; в богатых ими (эвтрофных) водоемах они также поглощают органические вещества (миксотрофность). В районах умеренного климата различия температуры воды, освещенности, значения pH и других показателей в зависимости от времени года существенно влияют на состав планктона. В пресной воде амплитуда колебания температур значительно больше, чем в море. Их диапазон находится в области от температуры таяния (около 0°С) ледников и полярного льда — мест, где обитают определенные, часто окрашенные в красный цвет хламидомонады, Chlorococcales и Mesotaeniaceae, составляющие «криопланктон», до температур горячих источников, в которых еще могут развиваться некоторые диатомеи (до 50 °С) и прокариотные синезеленые водоросли (до 75 °С). В бентосе пресных вод явно доминируют по массе и видовому разнообразию цветковые растения, и только при определенных условиях доминируют водоросли (например, Charaсеае).
Нейстон (сообщество поверхности воды) представлен прежде всего одноклеточными водорослями, например, видами Euglena и Chromulina rosanoffii. Для последнего вида, который придает поверхности воды золотистый налет, отмечалась плотность до 40 000 клеток на 1 мм2. Следует различать обитающий над поверхностной пленкой воды эпинейстон и опускающийся от нее в воду гипонейстон.
Водоросли и качество воды
С усиливающимся эвтрофированием водоема существенно увеличивается образование биомассы, а с ним и потребление кислорода: на дне откладывается (вместо минеральных отложений олиготрофных озер) гниющий ил. Бедные питательными веществами (олиготрофные) водоемы вследствие искусственного удобрения садов и полей, а также общего загрязнения воды приходят в сильный упадок. По наличию видов-индикаторов (планктонных и бентосных) можно судить об уровне загрязнения и соответственно о качестве воды (уровне сапробности, обозначаемом цифрами от I до IV). Наряду с эукариотными водорослями для этого в качестве индикаторов привлекаются также прокариоты (синезеленые водоросли, бактерии) грибы и высшие растения.
Наиболее сильное загрязнение обозначается как IV (полисапробная зона). Здесь из-за недостатка кислорода преобладают процессы гниения. Кислородное истощение чрезвычайно велико. При экстремальных условиях жизни в полисапробной зоне имеет место массовое развитие бактерий, в том числе Beggiatoa, а также прокариотных синезеленых водорослей из родов Spindina и Anabaem. Однако почти полностью отсутствуют эукариотные водоросли и водные высшие растения; единственное исключение составляют виды Euglena и Cartería. Наряду с этими немногими водорослями зеленого цвета встречается также бесцветная гетеротрофная Polytoma. Даже при сильном загрязнении, например, при поступлении неочищенных сточных вод, может происходить в определенной степени самоочищение водотоков.
В сильно загрязненных водах III класса (α-мезосапробная зона) бурно включаются окислительные процессы. Высокая численность различных бактерий характерна и для этого состояния, но при этом наблюдается и массовое развитие водорослей — синезеленых, а также диатомовых и зеленых; начинают хорошо развиваться и некоторые высшие растения. Содержание кислорода может быть значительным и в дневное время превышать значение насыщения, однако ночью оно сильно падает. Из прокариотных водорослей здесь встречаются различные виды Oscillatoria (тогда как в водах IV класса — только О. pulida и О. chlorina) и Phormidium; из диатомей — Stephanodiscus; из конъюгат — Closterium leibleiniiи Cosmarium botrytis; из прочих зеленых водорослей — Chlamydomonas и Gonium. Также характерны грибы сточных вод из родов Leptomitus и Fusarium.
Сильно загрязненные воды II класса качества (β-мезосапробные) характеризуются еще более успешными окислительными процессами; соответственно кислородное истощение относительно незначительное. В этой зоне число живых бактериальных клеток становится еще меньше. Наоборот, наблюдается большое разнообразие диатомовых и зеленых водорослей. Синезеленые водоросли представлены Anabaena flos-aquae, Nostoc и некоторыми видами Oscillatoria. Из диатомей встречаются различные виды родов Melosira, Asterionella и др., из золотистых водорослей — Synura. Из зеленых водорослей можно назвать Pediastrum, Scenedesmus, Chaetophora и Oedogonium. Из десмидиевых особенно распространены различные виды Closterium.
Почти незагрязненные воды классифицируются как I класс качества (олигосапробные). В этой зоне вода, если не считать временного «цветения», прозрачна и богата кислородом. Если эта область расположена ниже по течению от загрязненного участка реки, то органика здесь уже разложена, интенсивность очень быстро протекающих процессов разложения снижается, а процессы окисления завершены Число живых бактериальных клеток снижается до минимальных значений. Из синезеленых водорослей характерны, например, представители рода Наpalosiphon. Диатомеи представлены видами Surirella и Меridion, зеленые водоросли — видами Ulothrix, Сladophora (glomerata), Spirogyra (fluviatilis) и самыми различными видами десмидиевых (из родов Сlosterium, Staurastrum, Euastrum, Micrasterias), желтозеленые — видами Vaucheria. Типично также наличие таких пресноводных водорослей, как Lemanea annulata и Batrachospermum moniliforme.
Аэрофильные водоросли, симбионты, образователи горных пород
Лишь немногие водоросли живут как аэрофильные организмы вне воды, прежде всего на затененной стороне скал и стволов деревьев (например, водоросли типа Pleurococcus и Trentepohlia, см. рис. 11.93, А, С; «чернильные штрихи» из синезеленых водорослей). Наиболее часто они встречаются во влажных тропиках, где поселяются и на листьях. Подходящий известковый субстрат у поверхности (на глубину несколько миллиметров от края) часто пронизан водорослями. Еще шире распространены, но еще мало изучены почвенные водоросли. К «эдафону» — сообществу живых организмов почвы, относятся помимо синезеленых водорослей различные зеленые, желтозеленые и диатомовые водоросли. В 1 г почвы из самого верхнего слоя было обнаружено до 100 000 водорослевых клеток. Особым образом к наземному образу жизни приспособилась зеленая водоросль Eritschiella (см. рис. 11.99, С).
Важную роль в природе играют различные симбиотические водоросли. Также большое значение они часто имеют и как образователи горных пород (например, Dasycladophyceae).
Спермогонии (рис. 11.106, A: s, Е) возникают из материнской клетки, сначала делящейся на 8 клеток — октантов. Каждый октант делится затем тангентальными стенками на 3 клетки (Е). Так возникают в итоге 24 клетки, которые подразделяют круглый спермогоний следующим образом: 8 плоских внешних покровных клеток (щитков), которые из-за возникающих неполных радиальных перегородок приобретают вееровидную орнаментацию; 8 средних клеток (рукояток — manubria), кото-
рые отходят от центров щитков и вытягиваются в длину; 8 внутренних клеток (первичные клетки-головки), которые в конечном счете принимают округлую форму. Из-
за того что 8 щитков растут в ширину более интенсивно, чем в толщину, внутри шара возникает полость. К центру ее от центра каждого щитка направлены клетки-руко-
ятки с сидящими на них по одной первичными клетками-головками. Первичные головки дают начало 3 — 6 вторичным головкам. а из них вырастает по 3 — 5 длинных неветвящихся сперматогенных нитей из одного ряда клеток, заполняющих полость (рис. 11.106, В, С). В каждой из дисковидных клеток образуется по одному спирально изогнутому сперматозоиду (D) с 2 жгутиками и глазком, но без пластид.
Оогоний (см. рис. 11.106, А: о) содержит одну-единственную яйцеклетку, плотно забитую каплями масла и зернами крахмала. Этот оогоний выдается вперед и позже плотно обрастает 5 коровыми нитями, закрученными в виде левовращающей спирали. Их концы отчленяют (путем образования поперечной перегородки) клетки коронки (А: с), между которыми проникает сперматозоид. После оплодотворения зигота окружается прочной бесцветной стенкой. Кроме того, внутренние стенки коровых нитей утолщаются, становятся коричневыми и часто инкрустируются известью, тогда как внешние мягкие стенки вскоре после отделения «ооспоры» (покоящегося органа) исчезают. При прорастании зиготы происходит мейоз; из 4 гаплоидных ядер 3 дегенерируют, так что возникает только один проросток.
Своеобразноe строение таллома, а также спермогонии и оогонии с их особыми защитными покровами, не имеющими аналогов среди других растений, и спирально закрученные сперматозоиды говорят о явно обособленном положении Charophyceae в системе живых организмов. Все эти признаки свидетельствуют об уникальности харовых, однако по составу пигментов и запасных веществ они сходны с другими Chlorophyta и Streptophyta.
Ископаемые харовые (особенно в форме их зигот) известны с силура; из 6 семейств, ранее существовавших на Земле, ныне сохранилось только одно.
Обзор Streptophytina. От исходных стрептофитов взяла начало большая часть ныне живущих наземных зеленых растений. От них возникли уже рассмотренные зеленые водоросли (II; Streptophytina) и пока еще не рассматривавшиеся мхи (см. Bryophytinа) и сосудистые растения (см. Pteridophytina и далее). Эволюционное родство этих групп подтверждается совпадением таких признаков, как: ультраструктура хлоропластов: пигментный состав (хлорофиллы а и b!); расположение и строение пиреноидов; крахмал в качестве запасного питательного вещества; изоконтные подвижные стадии с латерально расположенными жгутиками; целлюлоза в качестве строительного материала клеточной стенки, причем в упорядоченной кристаллической форме и в виде микрофибрилл. Поперечные перегородки внутри клеток возникают во фрагмопластах. У зеленых водорослей (II) уже встречается своего рода «плодообразование», так что оплодотворение индуцирует развитие покровных нитей, обрастающих оогоний (Coleochaete). У Charophyceae покров оогония закладывается до оплодотворения. У некоторых представителей (например, Chara) в ♂ гаметах хлоропласта полностью редуцированы. У Zygnematophyceae отсутствуют клетки, имеющие жгутики. У отдельных видов как защитные вещества встречаются спорополленин и лигниноподобные соединения (Coleochaete).
Однако мохообразные и сосудистые растения, несмотря на сходство по многим признакам с зелеными водорослями (II), претерпели существенное дальнейшее развитие генеративных и вегетативных структур, что свидетельствует о явном эволюционном прогрессе (приспособление к наземной жизни!).
Второй подотдел: мхи (Bryophytina)1
1 Авторы придерживаются далеко не общепринятой версии системы этой группы. — Примеч. ред.
В большинстве своем мхи уже выработали весьма высокоорганизованные тканевые талломы2; этим они отличаются от зеленых водорослей из предыдущего отдела, которые еще приспособлены к жизни в воде.
2 Большинство мхов имеют побеговую организацию. Примеч. ред.
Бокс 11.8. Эмбриофиты
К эмбриофитам относят растения, спорофит которых начинает существование как покоящийся многоклеточный зародыш, питающийся от материнского растения и часто долгое время остающийся в покоящемся состоянии (как у покрытосеменных). Правда, наличие зародыша в зачаточной форме наблюдается уже у некоторых живущих в воде водорослей (Coleochaete) с покоящимися и защищенными гаметофитом зиготами.
Типичные эмбриофиты (мхи и сосудистые растения) — это первично наземные растения с постепенно усложняющимися вегетативными органами, которые служат для закрепления в почве, поглощения воды и питательных солей, фотосинтеза (см. рис. 4.8; 4.1.2 и 5.3.4). Из исходно талломного вегетативного тела в ходе приспособления к наземному образу жизни и в связи с увеличением размеров и разделением функций развились разные органы: у гаметофитов высших мхов — каулоид, филлоид и ризоид (см. 5.3.4), у спорофита сосудистых растений — стебель, лист и корень (см. 4.1.2).
Размножение происходит в ходе гетерофазной и гетероморфной смены поколений, при которой в жизненном цикле преобладает или гаметофит (мхи) или спорофит (папоротники, семенные растения). После оплодотворения зигота развивается в многоклеточный зародыш, питающийся за счет материнского растения. Гаметангии — здесь их называютантеридии (♂) и архегонии (♀) — имеют оболочку (стенку) из стерильных клеток, защищающую гаметоциты. Спорангии эмбриофитов в отличие от спорангиев грибов и водорослей также имеют подобную оболочку из стерильных клеток. Такие защитные образования полностью отсутствуют у грибов и лишь изредка встречаются у гаметангиев водорослей.
Мейоспорангии грибов часто защищены сплетением гиф в плодовых телах, однако у них отсутствует оболочка из слоя стерильных клеток. У водорослей из родов Coleochaete и Charaоогоний (постгенитально) покрывается вырастающими нитями (см. рис. 11.105, 11.106, Е). С конгенитальными многоклеточными стенками антеридиев эмбриофитов можно сопоставить прежде всего стенку спермогония Chara.
Вегетативное тело построено из различных тканей, которые сильно дифференцированы и выполняют различные функции. Транспирация снижена кутикулой и чаще всего регулируется имеющимися устьицами. Транспорт воды и питательных веществ иногда идет по простым проводящим тяжам (мхи) или же по все более структурно усложняющимся проводящим пучкам (папоротники, семенные растения). Пластиды содержат хлорофиллы а и b, а также каротиноиды. Продукт ассимиляции — образующийся в хлоропластах крахмал. Клеточные стенки состоят из целлюлозы1.
1 Из целлюлозы состоит фибриллярный каркас клеточных стенок эмбриофитов, который погружен в «аморфный» матрикс из полисахаридов. — Примеч. ред.
Эмбриофиты подразделяют на мхи (Вгуоphytinal), папоротникообразные (Pteridophytina) и семенные растения (Spermatophyta). У семенных растений антеридии и архегонии сильно редуцированы, так что почти не различимы как таковые2. Семенные растения поэтому уже не относят к архегониатам в узком смысле (= мхам и папоротникообразным). Сборное понятие кормофиты (или также Сormobionta; в противоположность Рrotobionta с более простой организацией как, например, у водорослей и грибов) происходит от понятия «кормус» — вегетативного тела, дифференцированного на стебель, листья и корни (см. рис 4.8), — и включает мхи с иначе дифференцированными спорофитами, поскольку их во всяком случае можно вывести из теломов.
2 У всех современных голосеменных растений и эфедры архегонии легко различимы. — Примеч. ред.
Так, спорогоны мхов при нарушенном развитии, как исключение, могут вильчато ветвиться. Проводящие системы мхов, если они вообще имеются, построены достаточно просто. Однако они проявляют функциональное и даже определенное структурное сходство с проводящими элементами сосудистых растений. Все эмбриофиты близкородственны друг другу, так что их можно рассматривать как группу, имеющую общего предка, представители которой развивались в разных направлениях и достигли разного уровня развития.
В основе цикла развития мхов лежит четкая анизоморфная смена поколений (рис. 11.108, D), при которой зеленый фотоавтотрофный гаметофит чередуется со спорофитом. Гаметофит представлен либо морфологически слабо расчлененным лопастным талломом с ризоидами снизу (талломные мхи), иногда с высокой тканевой дифференциацией (например, на ассимилирующие и запасающие ткани), либо стебельками, варьирующими от лежачих до прямостоячих, покрытыми листиками и ризоидами (листостебельные мхи).Листики, за исключением средней жилки1, обычно однослойные (листья папоротников и семенных растений многослойные). Листостебельные мхи внешне несколько напоминают сосудистые растения, однако отличаются от последних, в частности, тем, что наивысшего морфологического и анатомического уровня дифференциации достиг гаметофит, а не спорофит. Кроме того, у мхов отсутствуют проводящие пучки, а в большинстве случаев и проводящие ткани. Ризоиды у мхов — это одноклеточные или многоклеточные нитчатые структуры, которые ни в коем случае не сопоставимы с высокодифференцированными корнями сосудистых растений (скорее их можно сопоставить с корневыми волосками). Кутикула мхов чаще всего очень нежная, и они чаще всего быстро высыхают при недостатке воды (пойкилогидричные растения, см. 6.10.3.6). Почти у всех гаметофитов отсутствуют устьица (исключение: антоцеротовые), однако для газообмена очень редко служат дыхательные отверстия (Marchantiales, см. рис. 11.100, А).
1 У юнгерманниевых жилки нет и лист целиком однослойный. — Примеч. ред.
Рис. 11.108. Bryophytina: А — сперматозоид достигает яйцеклетки; В — проникновение сперматозоида внутрь яйцеклетки; С — сперматозоид в ядре яйцеклетки, остаток его цитоплазмы остается в цитоплазме яйцеклетки; D — развитие двудомного листостебельного мха (спора, протонема, гаметофит — G, оплодотворение, спорофит — S, редукционное деление, споры). Светлые линии — гаплоидная фаза; темные линии — диплоидная фаза; g — жгутики, к — клеточное ядро р — пластиды, R! — редукционное деление
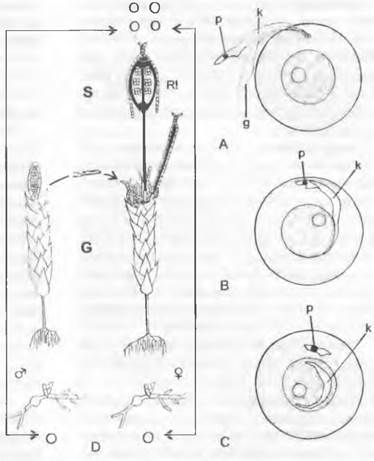
Архегонии мхов (см. рис. 11.111, J) представляют собой бутылевидные органы, части которых — брюшко и шейка — обычно имеют однослойную стенку. Брюшко заключает в себе крупную центральную клетку, которая перед созреванием делится на яйцеклетку и расположенную в основании шейки брюшную канальцевую клетку. К ней в шейке примыкают шейковые канальцевые клетки; таких клеток у мхов всегда несколько, располагающихся в один ряд (см. рис. 11.111, J).
Антеридии (см. рис. 11.111, Е) представляют собой округлые или булавовидные образования на короткой ножке. Развивающиеся в них сперматогенные клетки, окруженные стенкой антеридия, делятся каждая на 2 сперматиды, которые обособляются друг от друга и превращаются каждая в I сперматозоид.
Сперматозоиды всегда короткие, несколько спирально закрученные, нитевидные, основной объем в них занимает клеточное ядро. Вблизи переднего конца они несут 2 длинных гладких жгутика (F), направленные назад под острым углом от места прикрепления, а у некоторых представителей в цитоплазме имеется еще очень маленькая пластида (см. рис. 11.108, А: р). Яйцеклетки могут содержать также немного очень мелких хлоропластов.
Оплодотворение яйцеклетки даже у наземных форм может происходить только в присутствии воды (дождь, роса). Для этого архегоний открывается на вершине, канальцевые клетки ослизняются и выделяют определенные вещества, которые хемотактически привлекают сперматозоиды (см. 8.2.1.1). Из оплодотворенной яйцеклетки возникает диплоидный зародыш (см. рис. 11.111, К), который всегда без периода покоя развивается в спорофит.
Диплоидное спорофитное поколение развивается, таким образом, всегда на доминирующем гаплоидном гаметофите и остается связанным с ним на протяжении всей своей жизни. Изолированные спорофиты не могут полностью развиться, несмотря на наличие у них хлорофилла. Рост спорофита осуществляется, следовательно, в значительной мере за счет гаметофита. Часто встречающееся у растений пигание одного поколения за счет другого (например, у красных водорослей) называют «гонотрофия» (питание за счет производителя). Транспорт веществ в спорофит сокращается или вовсе прекращается, когда он достигает примерно 2/3 своего окончательного размера. Устьица, впервые появляющиеся в ходе эволюции высших растений у мхов, образуются почти исключительно на спорофитах (см. рис. 11.127, G). Спорофит внедряется своей базальной частью (гаусторией, называемой также стопкой, см. рис. 11.127, D; 11.120, С) в расположенную глубже ткань гаметофита, однако растет в основном в сторону вершины архегония, образуя округлый или овальный спорангий (коробочку, см. рис. 11.111, L, 11.123, М), на короткой либо более или менее длинной ножке. Весь спорофит называют спорогонием.
Во внутренней ткани коробочки — археспории — в результате двухкратных мейотических делений материнских клеток спор возникают мейоспоры в группах по четыре (в тетрадах), которые при созревании отделяются друг от друга и округляются. Распространение мейоспор осуществляется по воздуху. Стенка спор состоит из внутреннего нежного эндоспория и внешнего стойкого экзоспория; последний при прорастании споры разрывается. Споры прорастают в гаметофиты с образованием нитчатой или пластинчатой протонемы (см. рис. 11.123, А, В, С; 11.120, D; 11.108, D), которая вскоре переходит в зеленое растение мха1.
1 Это гаметофор. — Примеч. пер.
Наряду с размножением спорами у мхов очень часто встречается вегетативное размножение, например, посредством выводковых телец (см. рис. 11.110, А; 11.114, С, G; см. 10.1.3.3), которые могут различным образом возникать на гаметофитах: на стеблях, листьях или протонеме. Они отрываются и вырастают в новое растение.
Рост тканей осуществляется посредством 2-, 3- или многосторонней апикальной клетки (см. 5.3.4, рис. 5.13), реже — посредством меристемы (например, Riella, Riccia и Anthoceros).
Лигнина у мхов нет, лишь крайне редко встречаются сходные с лигнином соединения (как уже у некоторых зеленых водорослей — Coleochaete).
Систематика. К мхам относят примерно 24 000 видов, которые можно подразделить на четыре эволюционно далекие друг от друга группы: Marchatiopsida, Jungermа nniopsida, Bryopsida и Anthoeerotopsida. Bryopsida называют также листостебельными мхами (всегда олиственные; листья с особыми признаками), тогда как представителей всех остальных классов объединяют под названием печеночники. Однако представители Anthoeerotopsida, очевидно, эволюционно ближе к остальным наземным растениям, чем к любой другой группе мхов, так что они здесь приведены в Приложении к листостебельным мхам.
1. Класс: Маrchantiopsida (= Нераticае р.р., талломные печеночники)
Гаметофит Маrchantiopsida представляет собой уплощенный, чаще всего более или менее вильчато разветвленный, сильно дифференцированный тканевой таллом. С нижней стороны он обычно несет гладкие и так называемые язычковые ризоиды, которые имеют вдающиеся внутрь утолщения стенок. В типичном случае антеридии и архегонии приподняты на особых веточках (гаметангиофорах, или подставках). В клетках содержится по одному или по нескольку окруженных одинарной мембраной «масляных телец» (представляющих собой слившиеся капли терпенов), которые в такой форме отсутствуют у всех других растений (см. рис. 11.110, G: oк). Спорофитдлительное время полностью окружен разрастающейся стенкой архегония — эмбриотекой, которую он только незадолго до созревания прорывает на вершине. В созревающем спорогонии каждая клетка археспория делится на материнскую клетку споры и элатеру, которые, таким образом, являются синхронно возникающими сестринскими клетками.
Из материнской клетки споры после мейоза возникает по 4 споры. Соотношение 4: 1 гаплоидных спор (4) и стерильных диплоидных элатер (1) может в противоположность Аnthocerotopsida сдвигаться в сторону числа спор (например, 8: 1, 128: 1). Материнские клетки спор и элатер отделяются друг от друга продольными стенками, т. е. расположенными параллельно продольной оси спорогония.
Стенки элатер чаще всего со спиральными утолщениями (у Аnthocerotopsida они обычно гладкие). Не исключено конвергентное развитие элатер у Аnthocerotopsida, Маrchantiopsida и Jungermanniopsida1. Элатеры относятся к структурам, способствующим распространению спор (таким, как капиллиций — см., например, Trichia', волокна глебы — см., например, Lycoperdon: гаптеры — см., например, Equisetum), которые следует рассматривать как аналогичные образования.
1 Элатеры Аnthocerotopsida настолько отличаются от элатер печеночников, что их конвергентное возникновение давно не вызывает сомнений. — Примеч. ред.
Антеридий (см. рис. 11.111, Е) возникает из одной клетки эпидермы 1, которая вертикальными, взаимно перпендикулярными стенками делится на 4 клетки, после чего в квадрантах этой башнеобразной структуры происходит разделение тангенциальными стенками периферических покровных клеток и внутренних клеток, дающих начало сперматогенной ткани2.
1 Точнее, протодермы. — Примеч. ред.
2 Это неточное описание развития антеридия в данной группе. — Примеч. ред.
При развитии архегониев (см. рис. 11.111, J) одна клетка эпидермы3, выступающая над соседними клетками, делится периклинальной стенкой на нижнюю клетку, дающую начало ножке и верхнюю клетку — зачаток архегония. Три антиклинальные стенки разделяют его на центральную осевую клетку и на три покровные клетки, образующие тангенциальные секторы. На поперечном срезе через молодой зачаток архегония можно видеть все 4 клетки, на продольном срезе — осевую клетку и только две из трех покровных клеток. Осевая клетка сбоку окружена покровными, а сверху свободна; она расчленяется позже поперечной перегородкой на кроющую и внутреннюю клетки. Из покровных клеток без существенного участия клетки-крышечки образуется стенка шейковой и брюшной частей архегония, а из внутренней (центральной) клетки возникают 4 — 8 шейковых канальцевых клеток, одна брюшная канальцевая клетка и (в основании) яйцеклетка. Иное развитие архегониев у папоротников см. в соответствующем разделе.
3 Протодермы. — Примеч. ред.
Проводящие тяжи в гаметофитах чаще всего не формируются, а в спорофитах всегда отсутствуют полностью. Транспорт воды, например, у Marchantía, происходит через перфорированные клетки в талломе.
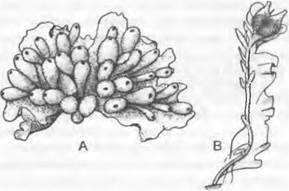
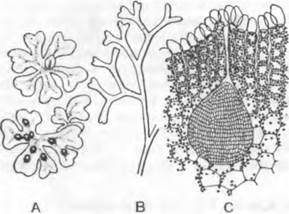
1. Порядок: Sphaerocarpales4. Иногда порядок выделяют даже в отдельный подкласс (Sphaerocarpidae). Таллом5 имеет простое строение: это растущие на почве мелкие розетки (Sphaerocarpos, рис. 11.109, А) или обитающие в воде вертикальные талломы, состоящие из оси (жилки) с отходящим от нее в одну сторону «крылом» (Riella, рис. 11 109, В). Архегонии и антеридии окружены грушевидными обвертками, открывающимися наверху. Стенка спорогония состоит из единственного слоя клеток, разрушающегося при созревании. У Sphaerococarpos (растение, широко используемое в генетических экспериментах) были впервые (1917 г.) в растительном царстве выявлены половые хромосомы. У него, как и у некоторых других мхов, определение пола происходит при мейозе материнских клеток спор.
4 Его целесообразно выделять в особый класс. — Примеч. ред.
5 Современные данные показывают, что это своеобразные побеги, а не талломы. — Примеч. ред.
Рис. 11.109. Marchantiopsida, Sphaocarpales: А — Sphaerocarpos michelii, ♀ таллом с обертками гаметангиев (5х); В — Riella helicophylla, ♀ таллом (2,5х)
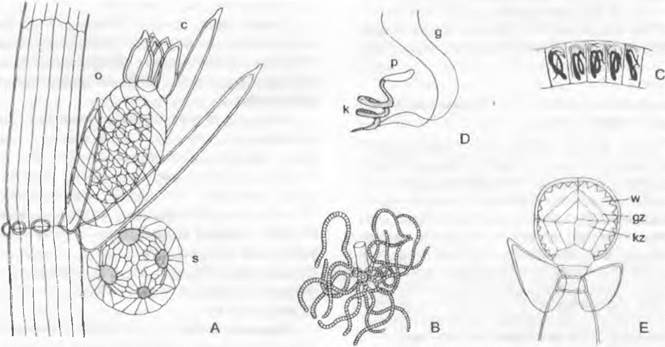
2. Порядок: Мarchantiales. Представители порядка имеют высокодифференцированный таллом. Как пример можно рассмотреть широко распространенную в сырых местах маршанцию обыкновенную (Маrchantia polymorpha, семейство Маrchantiaceae). Она образует лентовидно-уплощенные, несколько мясистые, вильчато разветвленные талломы (см. рис. II. 111, A, G) шириной до 2 см со слабо выраженным срединным ребром. Они нарастают за счет деления группы меристематических клеток на своих концах. На нижней стороне таллома располагаются однослойные брюшные чешуйки (амфигастрии) и отрицательно фототропные одноклеточные ризоиды (рис. 11.110, G), которые закрепляют таллом на субстрате и проводят воду вдоль него (преимущественно капиллярно между ризоидами, как по фитилю; отчасти за счет поглощения ими воды)1.
1 Функции закрепления растения и поглощения воды выполняют простые ризоиды, а ее проведения — язычковые. — Примеч. ред.
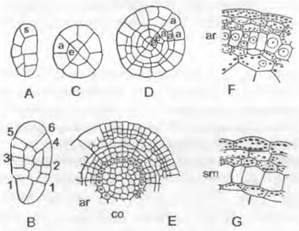
Рис. 11.110. Marchantiales, Marchantía, М. polymorpha: A — F— вегетативное размножение: А — разрез через выводковую корзиночку (12х), В — выводковое тельце в плане (80х), С — в поперечном разрезе; D — F — развитие выводкового тельца (300х); G — поперечный разрез таллома (200х); Н — дыхательное отверстие, вид сверху (200х); J — развитие ассимиляционных камер; а — ассимиляторы; as — место отделения выводкового тельца; о — верхний эпидермис с дыхательным отверстием (устьицем); ок — масляное тельце; óz — масляная клетка; г — ризоид; ri — зачаток ризоида; s — клетка-ножка; и — нижний эпидермис; v — брюшная чешуйка; w — утолщения стенок запасающих воду клеток
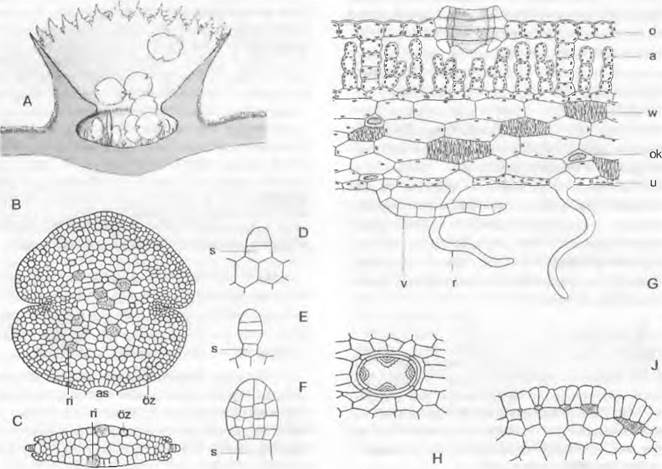
Под эпидермисом верхней стороны, покрытым почти водонепроницаемой кутикулой, расположены обширные межклеточные пространства (см. рис. 11.110, G, J) — «ассимиляционные камеры». Они отделены друг от друга перегородками, состоящими из одного или двух слоев клеток. На поверхности таллома эти перегородки видны как светлые линии, разделяющие темно-зеленые ромбические или шестиугольные участки. Со дна камер поднимаются многочисленные короткие, состоящие из округлых клеток разветвленные ассимиляторы, которые содержат хлоропласты и образуют ассимиляционную ткань 2 (см. рис. 11.110, G). Каждая камера сообщается с окружающим воздухом через бочонкообразное устьице. У Marchantía polymorpha оно состоит из четырех расположенных друг над другом колец, каждое из которых образовано 4 клетками. При недостатке воды устьице может даже немного сужаться, но для регуляции воды это еще не имеет существенного значения. Их строение препятствует проникновению воды в «устьице». Во всем растительном царстве едва ли найдется еще гаметофит с таким совершенным ассимиляционным и транспирационным аппаратом (см. рис. 11.122, А). Крупные, бедные хлорофиллом паренхимные клетки нижней стороны таллома служат как запаса- юшие (например, содержат масляные тельца, рис. 11.110, G: oк).
2 Комплекс ассимиляторов неправомерно называть тканью. — Примеч. ред.
По срединному ребру таллом на верхней стороне, как правило, местами разрастается в бокаловидные структуры с зубчатыми краями — выводковые корзиночки (рис. 11.110, А; 11.111, А) с некоторым числом плоских выводковых телец внутри. Последние возникают, как показано на рис. 11.110, D — F, путем разрастания наружу и дальнейшего деления отдельных поверхностных клеток; они сидят на клетке- ножке (s), от которой позднее отрываются (В: аs). В каждом углублении по бокам они имеют по точке роста и состоят из нескольких слоев клеток (С), из которых некоторые бесцветны и представляют собой зачатки будущих ризоидов (С: ri). Выводковые тельца вырастают в новые талломы и очень успешно обеспечивают вегетативное размножение гаметофитов.
Жизненный цикл. Гаметангии расположены на особых вертикальных ветвях таллома гаметофита (гаметангиофорах, или подставках) (см. рис. 11.111, А, G). В нижней части гаметангиофоры имеют вид трубчатой ножки, а в верхней части повторно вильчато ветвятся, образуя звездчатые «зонтики». Антеридии и архегонии возникают на разных растениях (двудомность). Определение пола осуществляется, как и у многих других мохообразных, гаплогенотипически через половые хромосомы (аналогично тому, как показано на рис. 10,19). В ножке подставки ризоиды, отходящие от ее брюшной стороны, оказываются при завертывании краев ножки внутри трубки (рис. 11.111, В, С); они доходят до нижней стороны таллома и капиллярно всасывают воду, как по фитилю.
Рис. 11.111. Marchantiales, Marchantía, М. polymorpha, половое размножение: А — ♂ растение с выводковыми корзиночками и антеридиофором; точки на поверхности таллома — устьица (1,5х); В — поперечный разрез через ножку антеридиофора немного ниже «зонтика» (13х), справа — спинная сторона с воздушными камерами, слева — брюшная сторона с двумя желобками, где проходят ризоиды; С — продольный разрез через антеридиофор (18х); D — развитие антеридиев (160х); Е — почти зрелый антеридий, продольный разрез (160х) F— сперматозоид (400х); G — ♀ растение с архегониофорами (1,5х); Н — продольный разрез через архегониофор; позади ряда архегониев— псевдоперихеций (25х); J— развитие архегониев (160х); К— продольный разрез через молодой, еще заключенный в стенку архегония спорогон окруженный псевдопериантием (35х); L — освободившийся из покровов спорогон, из которого выступают споры и элатеры; у основания ножки сохранился остаток стенки архегония (1 10х); М — споры и элатера (160х)
Антеридиофоры заканчиваются горизонтальным «зонтиком», который в результате трехкратного дихотомического ветвления становится по краю 8-лопастным (см. рис. 11.111, А). С верхней стороны на нем развиты антеридии, погруженные по одному в бутылевидную полость, которая открывается наружу узким отверстием (С). Эти полости отделены друг от друга ассимиляционными камерами. Вскрывание антеридиев и выход сперматозоидов происходят после дождя в результате того, что стенки антеридия ослизняются и расплываются. Сперматозоиды (рис. 11.111, Я) собираются на антеридиальной поставке в каплях воды (роса или дождь) и задерживаются там несколько загнутым вверх краем подставки.
Архегониофоры (G) в самом начале своего развития очень сходны с антеридиофорами. Архегонии закладываются восемью радиальными лучами, причем расстояние между двумя рядами, находяшимися против спинной стороны ножки, больше, чем между всеми остальными. Край молодого «зонтика» в процессе развития постепенно загибается вниз, так что группы архегониев оказываются на его нижней стороне (за счет чего исходно акропетальное заложение архегониев становится базипетальным). Наконец, находящиеся между группами архегониев ткани разрастаются во вторичные лопасти «зонтика» (общим число девять!); две из них развиваются между наиболее удаленными друг от друга группами архегониев (см. выше).
Оплодотворение осуществляется при дождливой погоде, когда брызги от дождевых капель, содержащие сперматозоиды, попадают с ♂ на ♀ «зонтики». Эпидермальные клетки последних выступают вперед в виде сосочков1 и представляют собой поверхностную капиллярную систему. По ней сперматозоиды проводятся к архегониям, к которым они затем привлекаются по механизму хемотаксиса — вероятно, определенными белками (см. 8.2.1.1).
1 Это не сосочки эпидермальных клеток, а особая однослойная структура псевдоперихеций. — Примеч. ред.
Спорофит. Через несколько дней после оплодотворения зигота начинает развиваться в многоклеточный зародыш, который вырастает в небольшой овальный зеленеющий спорогон с очень короткой ножкой (рис. 11.111, К, L).
Из двух клеток, возникающих после первого деления зиготы, верхняя (обращенная к шейке архегония) дает начало округлой коробочке (экзоскопическое положение зародыша), тогда как нижняя образует гаусторию и ножку (L). Начальные стадии развития не совсем одинаковы у представителей разных родов и семейств. Периклинальными стенками зачаток коробочки разделяется на внутренние и наружные клетки (К); внутренние дают многоклеточную спорогенную ткань (археспорий).
Коробочка у Marchantía имеет стенку из одного слоя клеток с кольцевыми утолщениями стенок. Только на вершине стенка двухслойная. Коробочка вскрывается на верхушке несколькими зубчиками. Зрелая коробочка сначала некоторое время покрыта разрастающейся стенкой архегония (К), которая, однако, при вытягивании ножки разрывается и остается у основания в виде «воротничка». Кроме того, каждая коробочка окружена тонко-кожистой «частной обверткой» (= perichaetium)1с четырьмя или пятью зубцами по краю, которая еще до оплодотворения начинает расти из короткой ножки архегония в виде мешка (Н, К). Наконец, каждый радиальный ряд архегониев окружен еще выростом таллома — «групповой обверткой»(involucrum) в виде изящных зубчиков (Н)2.
1 В отечественной литературе «частную обвертку» называют периантий, а «общую обвертку» — перихеций. — Примеч. пер.
2 Она представляет собой бахромчатую по краю пластинку, а не зубчики. — Примеч. ред.
Коробочка высвобождает несколько сотен тысяч спор (L, М). Между спорами располагаются одноклеточные нитевидные элатеры с тонкими стенками со спираль-
ными утолщениями (М; процесс их возникновения — см. во введении к Marchatiopsida), которые после вскрывания коробочки совершают гигроскопические движения, разрыхляя и рассеивая споровую массу (L). Каждая прорастает в очень короткую хлорофиллоносную нить (протонему), которая сначала с помощью двусторонней верхушечной клетки, а позже более сложным образом вырастает в таллом.
Также растущий на скалах и сырой почве Conocephalum conicum (Conocephalaceae) сходен с Marchantía строением таллома, но имеет более простые устьица и у него отсутствуют выводковые корзиночки. Сперматозоиды из антеридиофора выбрасываются за счет тургорного давления на несколько сантиметров вверх. Lunularia (Lunulariaceae) имеет полулунные выводковые корзиночки и антеридиофоры без ножек. У самого мелкого представителя этой группы — Моnоcarpus sphaerocarpus (Monocaipaceae; Австралия) — сильно редуцированный таллом несет единственный округлый спорогон, который окружен относительно хорошо развитым покровом.
Представители семейства Ricciaceae устроены более просто (рис. 11.112). Вильчатые ветвления таллома посредством двусторонних верхушечных клеток (см. рис. 5.12) чаще всего быстро следуют друг за другом, так что возникают небольшие розетки1 (рис. 11.112, А). У некоторых видов таллом разделен на камеры и имеет простые устьица; однако у большинства видов верхняя часть таллома состоит из вертикальных рядов клеток, каждый из которых заканчивается более крупной бесцветной клеткой (С). Гаметангии, как и не имеющий гаустории и ножки спорофит, погружены в таллом1 (С). Большинство видов Riccia— напочвенные формы (А); лентовидная, дихотомически разветвленная R. fluitans (В) живет в толще воды, а Ricciocarpos natans подобно ряскам плавает по поверхности воды.
1 Они располагаются в углублениях поверхности таллома, а не внутри него. — Примеч. ред.
Рис. 11.112. Marchantiales, Riccia: А — R. glauca, внизу растение со спорогонами (2х); В — R. fluitans, погруженная водная форма (2х); С — R. glauca, поперечный разрез через таллом с антеридием (125х)
2. Класс: Jungermanniopsida (= Нерatiсае р.р., преимущественно листостебельные печеночники)
К этому классу относят талломные и листостебельные формы, между которыми имеются переходы. Первые слабо дифференцированы морфологически и анатомически; с нижней стороны они несут только гладкие ризоиды. Гаметофиты листостебельных форм несут простые листья без средней жилки, расположенные в два ряда2.
2 За редкими исключениями, листорасположение Jungermanniopsida трехрядное. — Примеч. ред.
Представителей класса Jungermanniopsida часто трактуют как прогрессивный ряд от талломных к листостебельным формам. Однако и листостебельные формы можно считать исходными и выводить из них талломные, предположив срастание в ходе эволюции налегающих друг на друга листьев и уплощение стебля.
Гаметангиофоры у гаметофитов отсутствуют. Масляные тела встречаются чаще всего в большом числе во всех клетках, тогда как у Маrchantiopsida локализованы в особых масляных клетках. Коробочка спорогона выносится на длинной ножке (приспособление к распространению спор ветром) и вскрывается чаще всего 4 створками (см. рис. 11.114, А). В предыдущем подклассе у спорогонов ножки отсутствуют или очень короткие. Элатеры сходны по строению и развитию с таковыми у предыдущего класса. Проводящих пучков в спорофите нет, в гаметофите они встречаются редко и тогда очень просты, т. е. сложены одними гидроидами (Symphyogyna, Нaplomitrium).
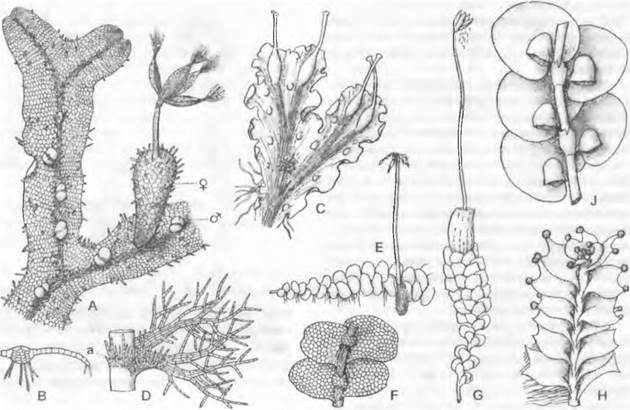
1. Порядок: Меtzgeriales. Таллом, нарастающий за счет одной верхушечной клетки (см. рис. 5.12, В, С) и чаще всего вильчато разветвленный, состоит из одного или нескольких слоев одинаковых клеток; у некоторых видов в нем имеется срединное ребро из удлиненных клеток (см. рис. 11.114, А, В), а у Symphyogyna даже есть примитивный центральный проводящий тяж. Род Fossombronia отличается двумя рядами косо прикрепленных листьев, многорядных в основании. Таким образом, мы имеем здесь переход от талломных к листостебельным формам. Архегонии развиваются под продолжающей дальнейший рост верхушечной клеткой; спорофиты, окруженные псевдоперихецием, сидят на талломе (А) или на коротких боковых ветвях («анакрогинно»). У видов некоторых родов элатеры в виде кисточки прикрепляются к верхним концам створок коробочки (Меtzgerа, А) или в середине ее дна (Реlliа). Большинство из 500 видов этого порядка, например, виды родов Riccardiа, Реlliа, Fossombrontа, обитает на сырой почве. Напротив, Мetzgeria растет на затененных скалах или как эпифит на коре лиственных пород. Род Вlasia (см. рис. 11.114, С) занимает, согласно данным новейших исследований, изолированное систематическое положение между Jungermanniopsida и Маrchantiopsida и поэтому, строго говоря, должен относиться к отдельному классу (Вlasiopsida). У Вlasia таллом имеет бутылевидные вместилища выводковых телец; он разделен по краям на листовидные лопасти и несет снизу мелкие чешуйки.
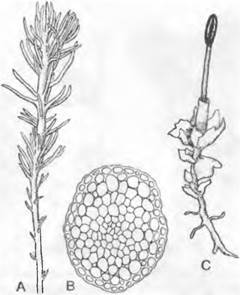
2. Порядок: Сalobryales1. Вертикальные стебельки несут три ряда одинаковых листьев и прикрепляются к субстрату мясистыми разветвленными «корневищами», лишенными ризоидов, но содержащими эндотрофно живущие грибы («микориза»). В частности, встречающийся и в Европе Нaplomitrium (см. рис. 11.113, С) имеет плоские, у основания многослойные листья и центральный тяж в каулоиде.
1 Эту группу с еще большим основанием, чем Blasia, следует выделять в особый класс. — Примеч. ред.
Рис. 11.113. Takakia lepidozioides (А — В) и Haplomitrium hookeri (С): А — внешний вид (6х); В — поперечный срез стебля (100х); С — Jungermanniopsida, Haplomitrium (6х)
3. Порядок: Jungermanniales. Представители порядка — преимущественно тропические растения, чаше все мелкие, живущие на почве и на древесных стволах, а в тропиках и на листьях лесных деревьев. Насчитывая около 9000 видов (в Средней Европе 250), они составляют примерно 90% всех печеночников. Их тело четко дифференцировано на лежачий или приподнимающийся, разветвленный, дорсовентральный стебель и однослойные листья без средней жилки, которые располагаются в два ряда2 по бокам стебелька под углом к нему (рис. 11.114, D — J). В стебле нет никаких проводящих тканей. У Jungermanniales нет ни воздухоносных щелей (как у Маrcantiales), ни настоящих устьиц (как у Аnthocerotales).
2 Листья чаше всего располагаются в три ряда. — Примеч. ред.
Листья, косо прикрепляющиеся к стеблю и поэтому расположенные подобно черепице, по- разному налегают друг на друга и весьма разнообразны по форме. Их называют набегающими, когда нижний край каждого листа покрыт верхним краем расположенного ниже (рис. 11.114, Е, F), и нисбегающими, если нижний край при рассмотрении сверху не закрыт (Н). Листья с закругленной или заостренной верхушкой (G), выемчатые (F), двузубчатые или многозубчатые, двухлопастные (Н) или рассеченные на нитевидные окончания (D). У эпифитной Frullania (J) одна из двух лопастей листа видоизменена в бокаловидную структуру, которая служит для удержания воды («водяной мешок»).
У большинства родов наряду с расположенными в два ряда спинными листьями есть еще расположенный на брюшной стороне ряд листьев меньшего размера и другого вида — это амфигастрии, или брюшные листья (например, у Frullania, Саlypogeiа, рис. 11.114, Е, J). Образование трех рядов листьев обусловлено наличием трехсторонней пирамидальной верхушечной клетки побега, одна сторона которой, однако, дает лишь мелкие листья или — у видов с двумя рядами листьев — вообще не дает листьев. Боковые ветви возникают рядом с листьями.
Расположенные на конце побега (акрогинные) архегонии окружены периантием (рис. 11.114, G), который состоит из трех сросшихся между собой верхних листьев. У некоторых видов клетки гаметофита под оплодотворенным архегонием делятся, так что возникает мешковидное вместилище (марсупий) для защиты молодого спорофита, вырастающего после оплодотворения яйцеклетки. Спорогон бывает уже полностью развитым, когда он вследствие растяжения своей ножки (см. 7.1.1) прорывает стенку архегония, остаюшегося в виде кожистой обвертки у его основания. В коробочке нет колонки. Оболочки клеток многослойной стенки коробочки имеют утолщения в виде колец или балок либо равномерно утолщены (если не считать тонких внешних клеточных стенок); разрыв происходит за счет силы поверхностного натяжения при потере клетками воды (см. 8.4).
Протонема Jungermanniales у разных родов устроена неодинаково, однако обычно состоит всего из нескольких клеток. У Меtzgeriopsis pusillla она, напротив, представляет собой пластинку и является по сути собственно вегетативным телом, на котором сидят растеньица с немногими листьями, служащие лишь для формирования половых органов. У Protocephalozja нитевидная протонема несет антеридии и архегонии1.
1 Как и у Metzgeriopsis, у Рrotocephalozia на долговечной протонеме образуются эфемерные побеги-гаметофоры, образующие половые органы. Непосредственно на протонеме половые органы никогда не образуются. — Примеч. ред.
Вегетативное размножение также широко распространено у Jungermanniales: у одних оно происходит посредством легко отламывающихся выводковых ветвей и выводковых листьев особого облика (часты у тропических эпифильных Lejeuniaceae), у других — посредством выводковых телец, образующихся преимущественно по краям или на конце листьев и состоящих из немногих или всего из одной клетки (см. рис. 11.114, Н).
К Jungermanniales относят также: Scapania, Lophozia и Trichocolea (см. рис. 11.114); Lophocolea (см. 7.1.1.); Cephalozia (см. рис. 8.38).
Рис. 11.114. Jungermanniopsida: А — С — Metzgeriales:
А — Metzgena conjugate (нижняя сторона) с несколькими ♂ и одной ♀ ветвями таллома, на 4 створках коробочки пучки элатер, ножка спорогона окружена псевдоперихецием (15х), В — М. conjugate, поперечный разрез таллома (30х), С — Blasia pusilla, с бутылевидными выводковыми корзиночками и многочисленными ушками на верхней стороне таллома, заселенными Nostoc (4х); D — J — Jungermanniales: D — Trichocolea tomentella, лист и амфигастрий (а) (7х), Е, F — Calypogeia trichomanis: Е — растение, вид сверху, с марсупием и зрелым спорогоном (2х), F — фрагмент с 4 листьями и 2 амфигастриями, вид снизу (6х), G — Scapania undulata с периантием и зрелым спорогоном (2х), Н — Lophozia ventricosa, вид сверху, с группами выводковых корзиночек на концах листьев (10х), J — Frullania dilatata, вид снизу, с «водяными мешками» (25х)
Эволюция. Положение Jungermanniopsida в системе противоречиво. Раньше на основании анализа ДНК предполагали их большую эволюционную близость к Bryopsida, чем к Marchantiopsida. Новейшие исследования подтверждают их самостоятельность как отдельного класса, однако одновременно указывают на более тесную связь с Marchantiopsida, чем с Bryopsida, так что оба класса (с Вlasiopsida, но не с Аnthocerotopsida) образуют таксон более высокого ранга. Согласно этой новой, здесь еще не используемой концепции мхи разделяют на три основные группы: Неpaticophytina (печеночники), Вryophytina (листостебельные мхи) и Аnthocerotophytina (антоцеротовые).
3. Класс: Вгуopsida (= Musci, листостебельные мхи)
Гаметофит листостебельных мхов всегда дифференцирован на стебель и листья. С помощью обычно разветвленных многоклеточных ризоидов с косыми перегородками (у ранее рассмотренных мхов они неразветвленные и почти всегда одноклеточные) он закрепляется в почве или подстилке (см. рис. 11.123, Е). В противоположность олиственным видам Jungermanniales побеги не дорсовентралъные, а радиально-симметричные, а листья чаще всего располагаются по спирали и при рассмотрении сверху выглядят трехрядными или многорядными. Боковые ветви образуются под листом (ср. рис. 5.13; отличие от семенных растений). Листья, нарастающие с помощью двусторонней верхушечной клетки, часто имеют среднюю жилку, а масляных телец у них нет (у листостебельных печеночников часто бывают капельки масла, а листья нарастают с помощью односторонней верхушечной клетки).
У листостебельных мхов с лежачими побегами листья при спиральном расположении отвернуты в одну или в две стороны, так что возникает различие спинной и брюшной сторон, однако иначе, чем у печеночников (например, Fissidens, см. рис. 11.126, С).
Листостебельные мхи отличаются от других мхов также и спорофитом: он чаше всего имеет устьица и развивается как коробочка с колонкой обычно на длинной ножке1. Элатер нет.
1 Как и у других мхов, он имеет еще и гаусторию. — Примеч. ред.
Жизненный цикл. Споры листостебельных мхов прорастают в гаметофиты, представленные вначале обильно разветвленной, положительно фототропичной зеленой нитью — протонемой (см. рис. 11.123, А). Протонемы при массовом развитии видны невооруженным глазом как зеленый войлок. Сначала развиваются богатые хлоропластами нити с перпендикулярными к их оси поперечными перегородками, которые называют хлоронемой. Хлоронема постепенно переходит в стелющуюся по субстрату каулонему, которая беднее хлоропластами и имеет косые поперечные перегородки. При достаточном освещении на ней, а чаще всего на ее коротких боковых ветвях, развиваются почки, дающие начало растеньицам мха1 (см. рис. 11.123, А). Из каулонемы возникают, кроме того, многочисленные, чаще всего направленные вверх боковые ветви типа хлоронем. Почка возникает таким образом, что после отделения одной или двух клеток-ножек вздувающаяся конечная клетка косыми стенками отделяет трехстороннюю пирамидальную верхушечную клетку (В, С), которая, откладывая сегменты, развивает олиственное растеньице мха. Там, где таких почек много, растеньица мха располагаются плотными дерновинками.
1 Протонема представляет собой ювенильную форму растения мха. На ней развиваются не растения мха, а гаметофоры. — Примеч. ред.
Половые органы располагаются у листостебельных мхов группами на концах главного побега или более мелких боковых ветвей. Они окружены самыми верхними листьями, которые часто видоизменены в особую «обвертку» — перихециальные листья2 (рис. 11.115).
2 Перихециальные листья окружают только собрания архегониев. Вокруг собраний антеридиев находятся перигониальные листья. — Примеч. ред.
Рис. 11.115. Bryopsida): А — G — развитие антеридия у Fuñaría hygrnmetríca~. А— поперечное деление зачатка антеридия, В — образование и С — деление верхушечной клетки, D — разделение на покров и зачаток сперматогенной ткани, Е — то же, на поперечном срезе (А — Е — 650х), F — почти зрелый антеридий (300х), G — продольный срез собрания антеридиев Mnium hornum, часть антеридиев видна с поверхности, часть дана в разрезе (100х); Н — L — развитие архегония у Mnium undulatum (250х): Н — ножка еще без зачатка архегония, J — архегоний (а) закладывается путем образования центральной клетки (отмечена точками), кроющей клетки (d) и клеток стенки, К — центральная клетка разделилась на яйцеклетку и брюшную канальцевую клетку, L — многочисленные шейковые канальцевые клетки отделяются от кроющей клетки; h — кроющие листья; р — парафизы; s — ножка
Листостебельные мхи могут быть обоеполыми (гермафродитными), однодомными или двудомными соответственно в зависимости от того, развиваются ли антеридии и архегонии на одном или на разных побегах того же растения или на разных растениях.
Между половыми органами обычно располагается некоторое число многоклеточных, часто имеющих округлые конечные клетки парафиз.
Антеридии и архегонии листостебельных мхов расположены на ножках и в отличие от остальных мхов (и архегониат в целом) (бокс 11.8, с. 204) их тело формируется из сегментов, откладываемых верхушечной клеткой (см. рис. 11.115).
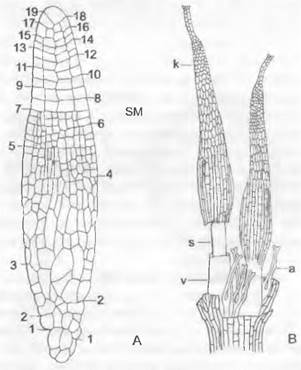
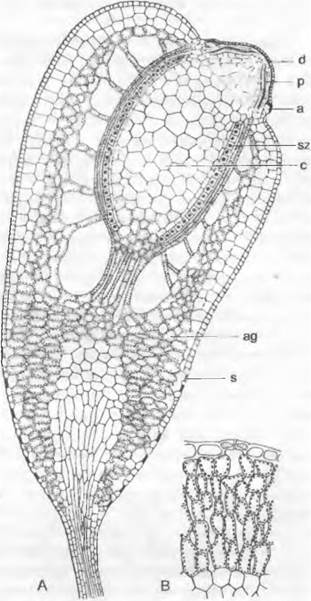
Спорофит. После оплодотворения яйцеклетки хемотаксически (см. 8.2.1.1) привлекаемым сперматозоидом зигота сначала многократно делится поперечными перегородками, и развивается сегментированный зародыш, вытянутый в длину. При типичном развитии в самой верхней его клетке возникают косые перегородки, которые отделяют клиновидную двустороннюю верхушечную клетку (рис. 11.116; 11.117, А, В; 11.118). Она отделяет в обе стороны сегменты, которые делятся дальше. В клетках сегментов, образующих коробочку, перегородка появляется по вертикальной радиальной стенке, так что на поперечном срезе зародыша видны 4 квадранта (см. рис. 11.117, С). Из них при делении периклинальными стенками возникают внешние (амфитеций) и внутренние (эндотеций) клетки (рис. 11.117, С, D: а, е). Самый внешний слой эндотеция чаще всего дифференцируется в археспорий (Е, F: аг), который целиком делится на материнские клетки спор (G; sm). Каждая материнская клетка мейотически делится на 4 гаплоидные споры. В противоположность Маrchantiopsidaвнутренние клетки эндотеция не участвуют в образовании археспория, а чаще всего образуют тяж из стерильной ткани — колонку (рис. 11.117, Е: со; 11.119, А: с), окруженную археспорием (рис. 11.119, А: sz).
Рис. 11.116. Bryopsida, Buxbaumiales: А — С — Buxbaumia aphylla: А — ♀, В — ♂ гаметофиты (А, В — 35х), С — спорофит; D — Diphyscium sessile
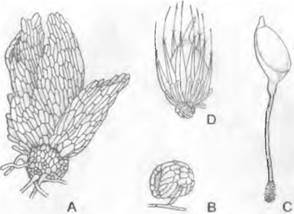
Рис. 11.117. Bryopsida, развитие спорогона Fuñaría hygrometríca: А, В — продольный разрез, первые деления зиготы; С — Е — поперечный разрез: С — деления в эндотеции (е) и амфитеции (a), D — дальнейшие деления, Е — более зрелый спорогон; в эндотеции самый наружный слой клеток — археспорий (аг) отделяется от колонки (со) (А — Е — 300х); F, G — поперечный разрез через археспорий (аr) и возникающие из него, еще не изолированные материнские клетки спор (sm) (250х); s — верхушечная клетка
Колонка служит для подведения питательных веществ и накопления воды для образующихся спор; питательные вещества спорам поставляют также богатые содержимым клетки вокруг археспория. Нижняя часть зародыша (рис. 11.118, А) — гаустория — закрепляется в ткани гаметофита.
Молодой спорофит (зародыш) сначала окружен оболочкой (эмбриотекой), которая образуется как из брюшка архегония, так и из ткани его ножки и даже из ткани стебля. По мере роста спорофита эмбриотека, не выдерживая натяжения, разрывается поперечно. При этом верхняя часть выносится на верхушке спорофита в виде колпачка, тогда как нижняя остается в виде влагалища (рис. 11.118, В).
Рис. 11.118. Bryopsida:
А — продольный срез молодого спорофита листостебельного мха Pogonatum urnigerum (150х); цифры указывают следующие друг за другом сегменты; сегменты 1 — 7 образуют ножку спорофита; В — Pottiales, Pottia lanceolata (40х), верхняя часть побега, листья удалены, два архегония оплодотворены: зародыш слева вследствие вытягивания ножки (s) поднял верхнюю часть эмбриотеки в виде колпачка (к), оставив нижнюю как влагалище (v); эмбриотека справа еще целая; а — неоплодотворенный архегоний; SM — начало меристемы ножки
На верхнем конце зрелой коробочки образуется своеобразная структура — колечко, предназначенное для вскрывания коробочки и рассеивания спор. Ножка выносит коробочку вверх, так что ветер может легко распространять споры. В молодом спорогоне снаружи от споровой массы находится деятельная ассимиляционная ткань, которая покрыта эпидермой1.
1 Ассимиляционная ткань находится значительно глубже эпидермы. К эпидерме она непосредственно примыкает в апофизе — части спорогона между коробочкой и ножкой. — Примеч. ред.
Рис. 11.119. Bryopsida, Bryidae: А — продольный разрез через спорогон листостебельного мха Fuñaría hygrometríca (25х); В — ассимиляционная ткань с устьицем (90х); а — колечко; аg — ассимиляционная ткань; с — колонка; d — крышечка; р — перистом; s — устьице; sz — спорогенные клетки
На спорогоне встречаются устьица Mnium-типа (рис. 11.119, А, В), свойственного также папоротникам. В разных семействах они значительно различаются по числу (от 3 до 300 на одну коробочку), форме и размеру.
Проводящие тяжи. Некоторые виды имеют просто устроенные проводящие ткани. Они могут быть и в гаметофитах, и в спорофитах. Как и в проводящих пучках сосудистых растений, проведение веществ осуществляет цепочка разных клеток. Для транспорта воды и солей служат гидроиды — вытянутые мертвые клетки, в полностью развитом состоянии, без ядра и цитоплазмы, с утолщенными боковыми стенками и косыми поперечными. В отличие от трахеид сосудистых растений их клеточные стенки не бывают ни одревесневшими, ни укрепленными кольцевыми или спиральными утолщениями. Продукты ассимиляции проводят другие, также вытянутые клетки — лептоиды, по развитию и строению напоминающие ситовидные элементы сосудистых растений. Боковые стенки их часто утолщены и в меньшей степени, чем иногда косые поперечные стенки, пронизаны ситовидными отверстиями с плазмодесмами. Вплоть до отмирания лептоиды содержат в своей цитоплазме ядра и пластиды. Обычно гидроиды располагаются внутри, а лептоиды снаружи. часто они перемешаны с другими элементами в центральном тяже. Последний окружен тканями из тонкостенных (внутренняя кора) и толстостенных (внешняя кора) клеток. Стереиды — это живые, имеющие ядро и пластиды, вытянутые в длину клетки, соседние с гидроидами. Их утолщенные, но не одревесневшие стенки служат для придания механической прочности (сравнимы с клетками колленхимы). Граничащие с лептоидами паренхимные клетки при большом структурном и функциональном сходстве с ними отличаются отсутствием ситовидных пор в поперечных стенках. От такого типа тяжей имеются различные отклонения — от упрощения строения (например, в случае отсутствия лептоидов) до полной редукции тяжей.
Тяжи листовых следов (см. рис. 11.123, Н), присоединяющиеся к центральному тяжу или слепо заканчивающиеся в коровой паренхиме, продолжаются в листья как проводящая ткань их средней жилки. Только при непосредственном соединении между гидроидами листа и центрального тяжа каулоида, т. е. при наличии настоящих листовых следов, возникают предпосылки для непрерывной системы проведения воды. Часто встречаются упрощения проводящей системы листьев: так, в коровой паренхиме каулоида гидроидов может быть очень мало или их нет совсем. В проводящих тяжах листьев для проведения ассимилятов, по-видимому, служат широкопросветные паренхимные клетки, тогда как типичные лептоиды здесь редки (Рolytrichales).
Для питания спорофита большое значение имеет его прикрепление к гаметофиту. Служащая как всасывающий орган гаустория внедряется в сильно разрастающуюся ткань ножки архегония, а в некоторых случаях (Роlytrichum) — даже глубоко в ткань стебля до его центрального тяжа. Гидроиды ножки спорогона тесно примыкают к гидроидам стебля гаметофита. На гаустории спорогона иногда встречаются придатки типа ризоидов, внедряющиеся в ткань гаметофита, но чаще — передаточные клетки, которые примыкают к клеткам паренхимы или лептоидам и отличаются увеличивающими поверхность утолщениями стенки в виде шишек или горбов. Цитоплазматические связи (через плазмодесмы) между спорофитом и гаметофитом, напротив, отсутствуют. Часто сочленение тканей между гаметофитом и спорофитом довольно несовершенно.
Вегетативное размножение. Мхи обладают чрезвычайно высокой регенеративной способностью. Так, обломившиеся стебли и листья могут непосредственно или через стадию протонемы вырастать в новые растения. У некоторых видов в пазухах листьев и на верхушках побегов вырастают группы клеток, которые могут опадать как «выводковые тельца» (см. рис. 11.126, О).
Листостебельные мхи подразделяют на три подкласса (Sphagnidае, Аndreaeidае, Вгyidае). В первых двух подклассах коробочка спорогона выносится вверх с помощью гаилоидной ложной ножки (удлиненной ножки архегония1).
1 Ложная ножка имеет более сложную природу, включая часть тканей стебля. — Примеч. ред.
1. Подкласс: Торфяные мхи (Sphagnidae)
Подкласс включает только одно семейство — Sphagnaceae, с единственным, но очень богатым видами (свыше 200!) родом Sphagnum. Сфагнум обитает в болотистых, преимущественно бедных известью местах, часто с низкими значениями pH, где образует большие подушки и ковры, из года в год нарастающие вверх, тогда как нижняя их часть отмирает и постепенно превращается в торф. В клеточных стенках откладываются лигниноподобные вещества.
Тетраэдрические споры прорастают в присутствии определенных микоризных грибов в протонему; сначала она нитевидная, а затем представляет собой маленький лопастной однослойный таллом с нитча
тыми ризоидами. Таллом образует чаще всего только один гаметофит с пучком ризоидов у основания (рис. 11.120, D).
Рис. 11.120. Bryopsida: А — Н — Sphagnidae, Sphagnum:
А — S. acutifolium, растение со спорогонами (2/Зх), В — S. squrrosum, зрелый спорогон на конце веточки (10х), С — S. acutifolium, молодой спорогон в продольном разрезе (17х), D — S. acutifolium, протонема с молодым растеньицем (100х), Е — S. molluscum, участок ветви с бутылевидными клетками, накапливающими воду; листья удалены (10x), F — то же, на поперечном срезе (10х), G — S acutifolium, участок однослойного листа; видны крупные водоносные клетки с кольцевыми утолщениями и отверстиями, а между ними — мелкие хлорофиллоносные клетки (300х), Н — то же, на поперечном срезе (300х), J — К— Andreaeidae, Andreaea mpestris: J — целое растение (8х), К — продольный разрез через молодой спорофит (40х); ah — шейка архегония; aw — эмбриотека; с — колонка; d — крышечка; f — гаустория: к — колпачок; ка — коробочка; pb — листья перихеция; ps— ложная ножка; s— споры; sf — гаустория спорогона; sg — спорогенная ткань; w — стенка коробочки
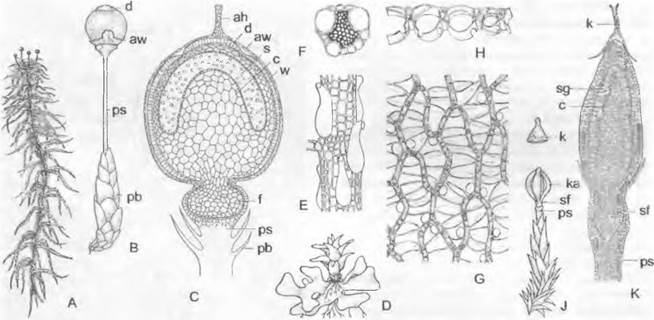
Вертикальные, лишенные ризоидов стебли почти всегда находятся в плотных дернинках и несут через регулярные промежутки пучки боковых ветвей. В каждом пучке часть ветвей отходит вверх, а часть — вниз, плотно прилегая к стеблю (рис. 11.120. А). На верхушке побега ветви образуют плотную розетку. Некоторые виды Sphagnum (особенно обитатели верховых болот) благодаря пигментам клеточных стенок окрашены в коричневый или ярко-красный цвет. Каждый год одна из ветвей на верхушке побега развивается так же мощно, как и соответствующий прирост материнского побега, так что происходит ложное вильчатое ветвление (ложная дихотомия). По мере того как стебли снизу постепенно отмирают, дочерние ветви становятся самостоятельными растениями.
Кора стеблей состоит из одно- или многослойной ткани из мертвых, пустых клеток, которые капиллярно всасывают воду; их боковые и поперечные стенки часто имеют округлые отверстия (рис. 11.120, Е). В листьях также находятся клетки с перфорированными стенками, усиленными кольцевыми или спиральными утолщениями (рис. 11.120, G); они располагаются поодиночке в ячейках однослойной сети из удлиненных живых клеток, содержащих хлоропласты. Эти своеобразные структуры служат для поглощения воды и минеральных веществ; растения могут таким образом удерживать воду, масса которой превышает сухую массу примерно в 20 раз. Листья не имеют средней жилки, а стебли — центрального тяжа.
Размножение. Отдельные ветви розетки имеют особый внешний вид и окраску. Они производят половые органы. Мужские ветви образуют в пазухах листьев округлые антеридии с длинными ножками (выходящие из них гаметы были первыми открытыми у растений сперматозоидами); женские ветви несут на конце архегонии, которые растут в отличие от остальных листостебельных мхов без верхушечной клетки — как у печеночников. Спорогоны развивают лишь очень короткую ножку с расширенной гаусторией. Они долгое время заключены в эмбриотеку, которую разрывают на вершине, оставляя ее у своего основания в виде оторочки (рис. 11.120, В: аw). В шаровидной коробочке спорообразующая ткань (С: s) куполообразно покрывает полушаровидную колонку. Археспорий возникает не из эндотеция, а из самого внутреннего слоя амфитеция. Спорогон своей расширенной гаусторией погружен во вздутый конец несущего его небольшого побега; после формирования спорогона этот конец вытягивается в ложную ножку значительной длины и выносит спорогон вверх (В: рs). За счет избыточного давления воздуха в коробочке крышечка с хорошо различимым шумом сбрасывается, и споры вылетают на расстояние более 20 см.
2. Подкласс: Аndreaeidae
Единственное семейство подкласса Аndrеаcaсеае представлено тремя родами. Виды рода Аndreaea (их насчитывается 120) образуют мелкие плотные темно-коричневые дернинки на неизвестковых скалах в высокогорьях, в Арктике и Антарктике. Спорогон, как и у Sphagnum, выносится вверх на ложной ножке, которая образуется из ножки архегония. Коробочка, сначала покрытая, как шапкой, колпачком, раскрывается четырьмя продольными щелями, причем получающиеся четыре створки остаются соединенными между собой на верхушке и у основания (рис. 11.120, J). Как и у Sphagnum, спорогенная ткань наподобие колокола окружает колонку (К). Протонема пластинчатая, разветвленная. У рода Andreaeobryum ложной ножки нет.
Ранее относимые к печеночникам Takakiales, вероятно, лучше трактовать как листостебельные мхи и включать в Andreaeidae1. Сюда относят азиатский род Takakia (см. рис. 11.113, А) с листьями, рассеченными до основания на 2 — 4 цилиндрических сегмента, и центральным тяжем в стебле.
1 Еще лучше рассматривать их как класс Таkakiopsida. — Примеч. ред.
3. Подкласс: Brydae
У представителей этого подкласса коробочка выносится вверх не принадлежащей гаметофиту ложной ножкой (как в обоих предыдущих подклассах), а образуемой спорофитом ножкой (диплоидной).
Гаметофит в этом подклассе достигает наибольшего разнообразия и наивысшей дифференциации среди листостебельных мхов; однако в немногих случаях он почти ограничен стадией протонемы (например, Ephemeropsis tjibodensis, Viridivellus pulchellum). Побеги или растут вертикально и несут на верхушке архегонии, а позже коробочку на ножке (акрокарпные мхи; см. рис. 11.123, Е), или они плагиотропные и одновременно чаще всего перисто-разветвленные, образующие архегонии, а позже коробочки на коротких боковых ветвях (плеврокарпные мхи; см. рис. 11.126, R). В стебле обычно проходит один центральный тяж (см. рис. 11.123, Н), который у наиболее высокоразвитых форм (Polytrichum)достигает значительной гистологической дифференциации (см.: Систематика, с. 233).
Листья состоят преимущественно из одного слоя клеток. Краевые клетки листовой пластинки часто образуют особую кайму (см. рис. 11.123, К: наружный слой) или зубчики. Клетки листа у акрокарпных мхов обычно паренхимные (изодиаметрические, см. рис. 11.123, К), у плеврокарпных, напротив, они в основном прозенхимные (вытянутые, как на рис. 11.120, G). Верхушечная клетка листа у акрокарпных форм дает несколько дочерних клеток, которые затем делятся более или менее перпендикулярными друг к другу перегородками, так что возникает сеть из изодиаметрических клеток. У плеврокарпных видов дочерние клетки, отчленяемые верхушечной клеткой косыми перегородками, быстро делятся дальше на ромбические клетки, боковые углы которых вытягиваются, в результате возникает сеть из прозенхимных клеток. Листья (особенно с сетью прозенхимных клеток) часто имеют среднюю жилку (см. рис. 11.123, J — L).
Коробочка у Bryidae также достигает наибольшего совершенства. Спорогон состоит из тонкой упругой ножки (см. рис. 11.123, Е; 11.126) и радиальной (см. рис. 11.123, Е) или дорсовентральной (см. рис. 11.116, С) коробочки, сначала покрытой колпачком (верхней частью эмбриотеки: см. рис. 11.118, В; 11.123, Е, М), позднее отпадающим1. Шейка архегония вскоре засыхает и остается как шпиль на колпачке. Колпачок, таким образом, состоит не из диплоидной ткани спорофита, а из гаплоидной ткани гаметофита (см. рис. 11.108, D). Самую верхнюю часть ножки под коробочкой называют апофизой; здесь обильнее всего развиваются устьица. По всей длине коробочки проходит колонка, вокруг которой в виде полого цилиндра располагается спорогенная ткань (см. рис. 11.119, A: sz). Колонка и спорогенная ткань, кроме того, окружены крупными межклетниками (см. рис. 11.119), которые образуются в амфитеции и особенно сильно увеличиваются при созревании. Более или менее шаровидные мейоспоры чаще всего содержат многочисленные хлоропласта (см. рис. 11.123, О).
1 В состав спорогона входит также гаустория. — Примеч. ред.
Верхняя часть стенки коробочки развивается как крышечка (см. рис. 11.119, A: d; 11.123, М). Под краем крышечки часто находится узкая зона в виде венчика — колечко (см. рис. 11.119, А: а; 11.124). Его клетки содержат расплывающуюся слизь и обеспечивают таким образом отбрасывание крышечки при созревании спор (колпачок отпадает еще раньше). По краю остающейся после опадения крышечки урночки у большинства листостебельных мхов находится перистом (см. рис. 11.119, А: р; 11.123, N), как правило, сложенный зубцами и изначально прикрытый крышечкой. У остальных мхов перистома нет. Строение перистома очень разнообразно.
Рис. 11.121. Вгуорsida, Вгуdaeае, Leucobryum glaucum: А — гаметофит со спорофитами (естественная величина); В — строение листа: два слоя лишенных цитоплазмы клеток, связанных друг с другом крупными отверстиями в стенках; между ними мелкие, вытянутые в длину клетки, содержащие хлоропласты (300х)
Рис. 11.122. Вгуidaeае, Роlytrichales: А — строение листа Роlytrichum formosum, на верхней стороне видны пластинчатые выросты из хлорофиллоносных клеток (250х); В — Роlуtrichumjuniperinum, поперечный срез стебля (120х); аг — внешняя кора; h — гидроиды; i — межклетники; ir — внутренняя кора; I — лепто- иды; z — центральный тяж
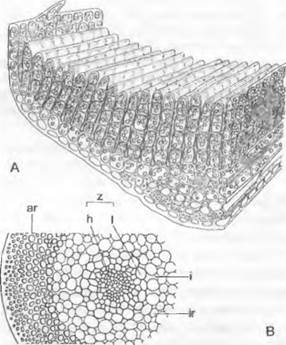
Рис. 11.123. Вгуidае, Вгуаles, Мnium puncatatum: А — протонема с почкой (20х); В — возникновение почки на протонеме; хлоропласты в верхних клетках не показаны (80х); С — заложение трехсторонней верхушечной клетки (85х); D — ♂ растение (естественная величина); Е — ♀ растение со спорофитом (естественная величина); F — собрание антеридиев в продольном разрезе (15х); G — собрание архегониев в продольном разрезе (15х); Н — поперечный срез стебля с центральным тяжом и тремя тяжами листовых следов (40х); J — лист (4х); К — кончик листа (25х); L — поперечный срез нижней части листа (50х); М — зрелая коробочка, рядом крышечка и колпачок (4х); N — перистом; слева наружный перистом удален; один из трех зубцов наружного перистома в сухом состоянии отогнут (30х); О — спора (100х)
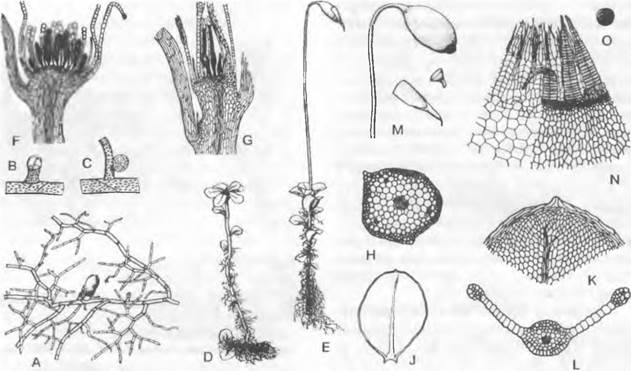
У немногих мхов (Рolytrichales, Тetraphidales, см. рис. 11.126, N) зубцы перистома состоят из рядов целых клеток. Однако у всех остальных мхов перистом образуется под крышечкой из утолщенных частей клеточных оболочек трех самых внутренних слоев амфитеция.
Возникновение такого перистома можно проследить на поперечном (рис. 11.124, В) и продольном (рис. 11.124, А) срезах верхней части коробочки. Тангенциальные стенки между первым и вторым слоем клеток утолщаются особенно сильно, а стенки между вторым и третьим слоем утолщаются слабее; радиальные, а также неутолщенные части тангенциальных стенок трех слоев клеток в конце концов разрушаются, так что сохраняются только утолщенные части тангенциальных стенок клеток. Они и представляют собой перистом, который, таким образом, является двойным (рис. 11.123, N) и состоит не из целых клеток, а только из сохраняющихся тангенциальных стенок. Наружный перистом состоит из 16 поперечнополосатых зубцов (рис. 11.123, N), расположенных по внутреннему краю стенки урночки; внутренний перистом («реснички») плотно прилегает к наружному и сложен узкими пластинками и нитями, которые имеют поперечные утолщения на внутренней поверхности1 и в своей нижней части сливаются в общую мембрану (рис. 11.123, N; 11.124, А; ip). Между каждыми двумя зубцами внешнего перистома располагается по две реснички внутреннего перистома2 (группа диплолепидных мхов в противоположность гаплолепидным только с одним кольцом зубцов перистома).
1 На внешней. — Примеч. ред.
2 Взаимное расположение элементов наружного и внутреннего перистомов разнообразно. — Примеч. ред.
Рис. 11.124. Bryidae:
А — Fuñaría hygrometrica, продольный срез верхней части коробочки листостебельного мха перед вскрыванием (200х), В — Mnium punctatum, поперечный срез коробочки в области перистома (120х); ар — наружный перистом; ад — ассимиляционная ткань; с — колонка; d — зона разрыва; ed — эпидермис крышечки; ек — эпидермис урночки; ip — внутренний перистом; s — материнские клетки спор; w — толстостенные клетки, соединяющие зубцы наружного перистома с эпидермисом; I — III — клетки кольца; 1 — 3 — три самых внутренних слоя амфитеция
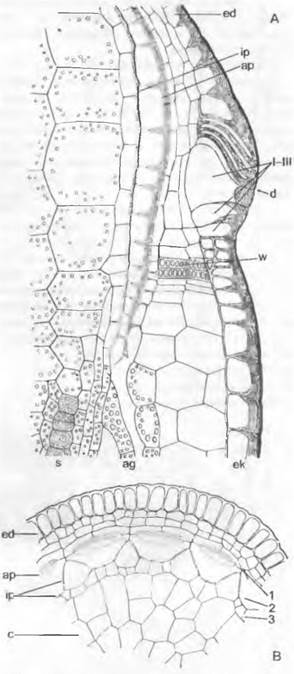
Наружные зубцы перистома совершают гигроскопические движения (см. рис. 8.36; см. 8.4), закрывая или открывая коробочку (рис. 11.123, N) в зависимости от погоды (в сухую погоду чаще всего изгибаются наружу), и таким путем способствуют постепенному рассеиванию спор. Наклоненные спорогоны и спорогоны с широким отверстием урночки чаще всего имеют хорошо развитый перистом, тогда как у родов с вертикально расположенным спорогоном, имеющим узкое отверстие урночки, перистом часто редуцирован (рис. 11.125, D).
Рис. 11.125. Bryidae, Schiatostegales, Schistostega pennata: A — несущее коробочку растеньице (103x); В — стерильное растеньице (10х); С— фрагмент предыдущего (50х); D — открытая коробочка (25х); Е— протонема («светящийся мох»), вид сбоку; стрелкой показано направление падения световых лучей (150х); F — то же, вид сверху (150х); G— выводковое тельце на протонеме (150х); Н — ход световых лучей в клетке протонемы
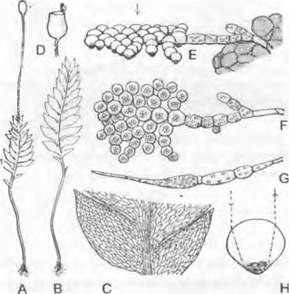
Поскольку можно заставить молодые спорофиты регенерировать протонему, то соответственно имеется и возможность получить диплоидные гаметофиты, которые в свою очередь могут образовывать тетраплоидные спорофиты. Неоднократно повторяя этот процесс, удалось получить гаметофиты с 16-кратным набором хромосом. Подсчеты числа хромосом у многих видов показали, что гаметофиты мхов часто содержат от двойных до четверных наборов хромосом на ядро и, таким образом, часто полиплоидны; и в этих случаях спорофит содержит вдвое больше хромосом, чем гаметофит.
Систематика. В основе систематического подразделения Bryidae, насчитывающих примерно 15 000 видов, лежат признаки как гаметофи та, так и спорофита (особенно перистома).
1. Надпорядок: Polytrichanae. Перистом состоит из целых подковообразных или вытянутых волокнистых клеток. Побеги акрокарпные, с высокодифференцированной проводящей тканью (непрерывная система проведения воды). Подземные «корневища» отличаются от надземных частей гаметофита приблизительно радиальным расположением проводящих воду гидроидов и напоминают этим корни двудольных цветковых растений. Листья сверху с продольными пластинчатыми выростами из богатых хлоропластами клеток («ассимиляционные пластинки», см. рис. 11 122).
1.1. Порядок: Dawsoniales. Дерновинки, состоящие из стройных растений высотой до 70 см, встречаются в Австралии и других областях Южного полушария.
1.2. Порядок: Роlytrichales. Отверстие урноч- ки в отличие от видов предыдущего порядка сначала закрыто тонкой пластинкой (эпифраг- мой). Рolytrichum (рис. 11.126, Т) с игловидными листьями, достигающий высотой 40 см, растет на почве в лесах и болотах. У Pogonatum протонема долговечная. Аtrichum undulatum с волнистыми листьями в виде язычков — мох, часто встречающийся на почве в лесах.
2. Надпорядок: Dicrananae. Перистом, если он не полностью редуцирован, простой (гаплолепидный). Преимущественно акрокарпные мхи.
2.1. Порядок: Dicranales. Перистом обычно состоит из 16 попарно соединенных зубцов. Виды Dicranum, отличающиеся серповидными листьями, часто обитают на почве в лесах. У Leucobryum жилка, почти полностью занимающая весь лист, состоит их клеток двух типов: зеленых живых и мертвых, накапливающих воду (см. рис. 11.121). Обитает на кислом лесном гумусе. Сюда относят также Аrchidium (см. рис. 11.126, А) со спорогоном без ножки, крышечки и перистома. Коробочка раскрывается беспорядочно за счет сгнивания (клейстокарпия).
2.2. Порядок: Fissidentales. Листья расположены в два ряда, со спинным крылом. В Германии встречаются виды родов Fissidens (рис. 11.126, С, D) и Octodiceras.
2.3. Порядок: Pottiales. Клетки листьев с шишковидными утолщениями наружных стенок («папиллами»). Tortula с длинным спирально закрученным перистомом (рис. 11.126, F). Eucladium (см.: Встречаемость и образ жизни мхов, с. 228) образует чаше всего подушки, инкрустированные известняком.
2.4. Порядок: Grimmiales. Преимущественно подушковидные обитатели каменистых субстратов не имеют признаков двух предыдущих порядков. Листья, например, у Grimmia (рис. 11.126, Н) и Rhacomitrium, часто вытянуты в бесцветные волоски или стекловидные окончания.
Последующие надпорядки отличаются двойным перистомом (диплолепидным).
3. Надпорядок: Bartramiales включает акрокарпные мхи. Коробочки, например, у Bartramia и Timmia, обычно наклонные, булавовидные и почти округлые, бороздчатые.
4. Надпорядок: Funariales. Представлен широко распространенным по всему земному шару видом Funaria hygrometrica (рис. 11.126, J), часто встречающимся на пожарищах. Это растущий на почве акрокарпный мох с крупными гладкими клетками листьев1. Splachnum luteum (рис. 11.126, К), растущий, как и другие виды этого рода, на помете травоядных животных — двудомный вид. Он отличается ярко окрашенной дисковидной апофизой спорогона. Женские растеньица (♀) приземистые, с более крупными листьями по сравнению с мужскими (♂); их архегонии окружены прилегающими листьями обвертки (у ♂ они оттопырены), так что возникает своего рода почка. Уже протонемы показывают половой диморфизм. У Ephemerum протонема сохраняющаяся.
1 Интересно происхождение видового названия этого мха — hygrometr¡ca (гигрометрическая): гигроскопичная ножка спорогония спирально скручивается в сухих условиях и раскручивается во влажных. — Примеч. пер.
5. Надпорядок: Bryales. Внутренние зубцы двойного перистома высокодифференцированы (см. рис. 11.123, N). Имеются переходы от акрокарпных к плеврокарпным формам, а также между листьями из паренхимных и прозенхимных клеток.
Сюда относят Bryum, Rhodobtyunt (см. рис. 11.126, L) с крупным пучком листьев и частые на почве в лесах виды рода Mnium (см. рис. 11.123), а также Mittenia plumula (Австралия, Тасмания, Новая Зеландия), протонема которой отражает свет (ср. «светящийся мох»).
Рис. 11.126. Bryopsida, Bryidae: А — Archidium phascoides, целое растение (5х) и коробочка (20х); В — Dicranum scoparium, трехлетнее растение (естественная величина); С — Fissidens bryoides (4х); D — то же, лист (15х); Е — G — Tortula muralis (4х): F — перистом (30х), G — лист со стекловидным волоском (10х); Н — Crimmia pulvinata (в натуральную величину); J — Fuñaría hygrometrica (2х); К — Splachnum luteum (в натуральную величину); L — Rhodobryum roseum (в натуральную величину), М — Tetraphis pellucida (2х); N — перистом; О — выводковая корзиночка (8х); Р — Climacium dendroides (в натуральную величину); Q — Hylocomium splendens, четырехлетнее растение (1/2х); R — Cratoneuron commutatum (1/2х); S — Раpillaria deppei (1/2х); Т — Polytrichum commune, справа молодой, покрытый колпачком спорогон (1/2х)

6. Надпорядок: Нypnanae. Здесь преобладает плеврокарпия. Клетки в листьях прозенхимные, жилка (если имеется) в отличие от видов предыдущих групп гомогенная, т. е. состоит из клеток одного типа.
6.1. Порядок: Neckerales. Внутренний перистом прямой коробочки обычно недоразвит. Сlimacium (рис. 11.126, Р) разветвлен наподобие маленького деревца.
Часто встречающиеся в тропиках эпифитные виды Рарillaria (S) и близких родов образуют «повислые формы». Fontinalis (см. с. 228) приспособился к жизни в воде. Виды Маcromitriumобразуют в коробочках более крупные ♀ и мелкие ♂ споры. Сюда же можно отнести и Orthotrichum.
6.2. Порядок: Ноokeriales. Перистом редуцирован; листья широкие, состоят из крупных клеток; пример — Нооkeria lucens, обитающая на влажной почве в лесах.
6.3. Порядок: Нурnales. Внутренний перистом высокодифференцирован. Длинные ножки заканчиваются обычно наклоненными коробочками. Вгachythecium, Нурnum, Нуlocomium(рис. 11.126, Q), Рleurozium и Р!аgiothecium с многочисленными видами, часто встречающимися на почве в лесах: к роду Cratoneuron (R) относят важные образователи известкового туфа.
Систематическое положение следующих порядков до сих пор неясно.
7. Порядок: Вuxbaumiales1. Протонема долговечная, богатая хлорофиллом, в значительной мере обеспечивающая питание гаметофита2 и спорофита. У Вихbaumia сильно редуцированный гаметофит3 (половой диморфизм; см. рис. 11.116, А — С) мало участвует в питании физиологически самостоятельного, снабженного обильной ассимиляционной тканью спорофита. У Diphyscium (D) лишенная ножки коробочка сидит в розетке из зеленых листьев.
1 Этот и следующий порядки, вероятнее всего, надо выделять в отдельные классы. — Примеч. ред.
2 Протонема — это стадия развития гаметофита, а не отдельное растение/поколение. — Примеч. ред.
3 На самом деле — гаметофор. — Примеч. ред.
8. Порядок: Теtraphidales. Четыре зубца перистома состоят каждый из нескольких продольных рядов клеток. Теtraphis pellucida (см. рис. 11 126, М — О) часто встречается на гнилой древесине.
9. Порядок: Schistostegales. Единственный вид порядка— «светящийся мох» Schistostega реnnata (см. рис. 11.125): отличается вторично двухрядными листьями, отсутствием перистома и сохраняющейся протонемой, которая размножается многоклеточными выводковыми тельцами. Протонема растет в расселинах скал и ямах в земле. Она образует шаровидные клетки, которые улавливают падающий на них свет и частично отражают его (Е, Н). Листья гаметофита сначала прикрепляются косо и расположены по спирали, однако в ходе развития принимают вертикальное положение перпендикулярно лучам падающего на них света (А. В).
4. Класс: антоцеротовые (Аnthocerotopsida)
Гаметофиты антоцеротовых талломные, однако отличаются от уже рассмотренных печеночников (классы 1 и 2) рядом признаков. В клетках гаметофитов нет масляных телец (у печеночников они встречаются часто) и имеется только по одному хлоропласту с пиреноидом. У гаметофита есть устьица (у печеночника Marchantía — напротив, только дыхательные отверстия). Спорофиты растут посредством интеркалярной меристемы, поэтому их рост не ограничен. Способ прикрепления спорофита к гаметофиту (передаточная ткань) сходен с тем, что мы наблюдаем у Psilotopsida из па поротникообразных, но не известен ни у одного из остальных мхов. Коробочка также имеет устьица и вскрывается двумя створками. Элатеры антоцеротовых отличаются способом возникновения и внешним видом от элатер печеночников (у листостебельных мхов элатер нет).
Антеридии с самого начала закладываются эндогенно (у печеночников они сначала экзогенные, но могут потом обрастать тканью гаметофита); архегонии также погружены в таллом.
Антоцеротовые составляют небольшую группу из примерно 100 видов, которые объединяют в единственный доживший до настоящего времени порядок.
Порядок: Anthocerotales. Гаметофит представляет собой дисковидный лопастной очень просто устроенный таллом длиной несколько сантиметров, прикрепляющийся к почве ризоидами (рис. 11.127, А). Клетки более или менее однородной паренхимы содержат в противоположность всем другим мхам только по одному крупному блюдцевидному хлоропласту с пиреноидом1. Эпидерма нижней стороны таллома имеет устьица с двумя бобовидными замыкающими клетками. Расположенная за ними подустьичная полость заполнена слизью и чаще всего заселена синезеленой водорослью Nostoc (В). Листьев и чешуек на нижней стороне таллома (брюшных чешуек), как и масляных телец в клетках, нет. Ризоиды гладкостенные.
1 У некоторых видов хлоропластов в клетке несколько. — Примеч. ред.
Рис. 11.127. Anthocerotopsida, Anthocerotales:
А — Phaeoceros laevis, таллом с молодыми и вскрывшимися спорогонами (2х); В — Anthoceros vincentianus, устьице на нижней стороне таллома, подустьичная полость заселена Nostoc (270х); С — Anthoceros punctatus, продольный срез молодого спорогона (130х); D — Dendroceros crispus, продольный срез почти зрелого спорогона (80х); Е — Anthocerospunctatus, неравные клеточные деления в археспории (100х); F — Anthoceros husnoti, поперечный срез спорогона с тетрадами спор и колонкой; стрелки — места разрыва стенки спорогона (100х); G — Anthoceros pearsoni, устьице спорогона (125х); а — археспорий; с — колонка; е — элатеры; f — гаустория спорогона; s — споры; sm — материнские клетки спор; w — стенка коробочки.
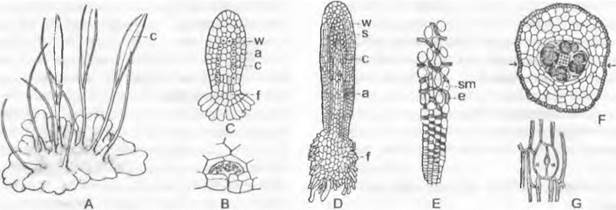
Антеридии и архегонии погружены в тал лом на его спинной стороне. Оплодотворенная яйцеклетка делится поперечной перегородкой на две клетки, из которых
верхняя, т. е. обращенная к шейке архегония, после дальнейших делений становится спорогоном, а нижняя — более толстой гаусторией, внедряющейся в таллом клетками типа ризоидов (рис. 11.127, D)1.
1 Противопоставление спорогона и гаустории ошибочно, так как гаустория — это орган спорогона. — Примеч. ред.
Спорогон представляет собой лишенную ножки щетинковидную коробочку 2 длиной от одного до нескольких сантиметров и вскрывающуюся, как стручок, двумя продольными створками (рис. 11.127. А). Она характеризуется (в отличие от коробочек Маrchantiopsida) сложной тканевой дифференциацией. Вдоль оси коробочки располагается стерильная колонка (С, D: с), состоящая из нескольких рядов клеток. Колонка окружена тонким слоем спорогенных клеток (археспорием, а), которые помимо мейоспор образуют также стерильные клетки — элатеры. Диплоидные материнские клетки спор и клетки, становящиеся элатерами, являются сестринскими; на каждую материнскую клетку спор (соответственно тетраду спор после мейоза) приходится одна зрелая элатера или одна еще способная к делению стерильная клетка, в результате последовательных митотических делений которой окончательное число элатер может увеличиться многократно. Элатеры располагаются перпендикулярно оси спорогона (Е).
2 Спорогон = коробочка + зона интеркалярной меристемы + гаустория. — Примеч. ред.
В отличие от всех других мхов коробочка спорофита созревает не одновременно, а длительное время удлиняется благодаря деятельности меристематической зоны у основания коробочки. В стенке спорогона имеются двуклеточные устьица (рис. 11.127, G), а кроме того, она содержит в своих клетках хлоропласта.
Представители семейства Аnthocerotaceae развивают на талломе спорогоны длиной 1 — 7 см. В Германии встречаются два рода: Anthoceros (например, А. puncatus) — со слизевыми вместилищами, которые на верхней стороне рассеченного таллома просвечивают точками, и темными густошиповатыми спорами; Phaeoceros (например, Ph. laevis на бедном известью жнивье) — без слизевых вместилищ в лопастном талломе, с желтоватыми, покрытыми сосочками спорами. Таллом Notothylасеае по сравнению с представителями предыдущего семейства крайне мал (несколько миллиметров в диаметре).
Палеонтологические свидетельства эволюции Anthocerotopsida, к сожалению, полностью отсутствуют. В отличие от печеночников и подобно листостебельным мхам, антоцеротовые имеют настоящие устьица. Этот признак сближает листостебельные мхи и антоцеротовые с высшими наземными растениями. Согласно данным анализа ДНК, антоцеротовые, очевидно, ближе к рассматриваемым дальше зеленым наземным растениям, чем прочие группы мхов.
Встречаемость и образ жизни мхов
Мхи завоевали сушу и населили ее множеством видов. Их приспособления к наземной жизни весьма разнообразны: например, значительная устойчивость к высыханию(пойкилогидрические растения, см. 6.10.3.6), сниженная или регулируемая транспирация (в частности, благодаря кутикуле; стерильным стенкам гаметангиев и спорогониев; устьицам; росту плотными подушечками и дерновинками), их приспособления к поглощению, накоплению и проведению воды, выработка различных жизненных форм, соответствующих обитанию на суше (например, древовидные — см. рис. 11.126, Р, листоватые — Q, повислые — S, войлоковидные, корковидные — R, подушечковидные — Н, дерновинные). которые обеспечивают заселение благоприятных ниш и приспособление к экстремальным условиям обитания.
Прочность тканей мхов обеспечивается набуханием клеточных стенок, а не тургором, как у высших наземных растений. Поэтому высушенные мхи при попадании в воду принимают свой исходный облик. Поглощение и отдача воды происходят, за немногими исключениями, всей поверхностью. Капиллярная система между стеблем, ризоидами и, если имеются, листьями обеспечивает значительное накопление воды, которое увеличивается у некоторых листостебельных печеночников благодаря развитию «водяных мешков» (см. рис. 11.114,1). черепицеобразному (набегающему или низбегающему — см. Jungermanniopsida) расположению листьев, наличию брюшных листьев и лопастей (см. рис. 11.114, F, Н, D), у листостебельных мхов — благодаря росту плотными высокими дерновинками. Marchantía, Sphagnum и Leucobryum накапливают воду также в водоносных клетках (см. рис. 11.110; 11.120, G; 11.121). Колонка в коробочке листостебельных мхов служит резервуаром питательных веществ и воды для образующихся спор. У определенных мхов (например, Fuñaría, Encalypta) накопление воды облегчает вздутый колпачок. Именно способностью мхов удерживать значительные количества воды в основном обусловлено буферное действие лесов в балансе воды в ландшафте. Обводненность верховых болот обусловлена осадками и очень большой способностью торфяных мхов (различных видов Sphagnum) накапливать воду.
Вышеупомянутая капиллярная система служит одновременно и внешней системой проведения воды, преобладающей у мхов. Мхи с центральным тяжем (к ним относится большинство акрокарпных листостебельных мхов и немногие печеночники) проводят поглощенную ризоидами воду по гидроидам. Особенно хорошо эта внутренняя проводящая система выражена у Polytrichanae, где даже имеются настоящие, связанные с центральным тяжом тяжи листовых следов, которые обеспечивают снабжение листьев водой (сомкнутая проводящая система).
Распространение сперматозоидов и спор. Если оплодотворение яйцеклетки сперматозоидами связано с наличием капельножидкой воды, то споры распространяются ветром. Их освобождение обеспечивают изменения влажности и специальные механизмы рассеивания (например, элатеры; механизмы вскрывания коробочек; см. 8.4). Зонтиковидная, ярко окрашенная апофиза у спорогонов видов Splachnum (см. рис. 11.126, К) способствует распространению склеенных в комочки спор насекомыми1.
1 Существенную роль в привлечении насекомых играет также запах, выделяемый через устьица на апофизе. — Примеч. ред.
Некоторые виды имеют хорошо развитые органы ассимиляции. Таллом Marchantía уже напоминает по анатомическому строению лист листостебельных растений, в том числе и приспособлениями к газообмену, хотя менее эффективными и имеющими другое строение. На верхней стороне листьев Polytrichum образуются свободно поднимающиеся вверх пластины, поглощающие свет для фотосинтеза. Там, где уже встречаются настоящие устьица (у антоцеротовых на талломах гаметофитов и на спорофитах, у листостебельных мхов — на спорофитах), они часто вторично нефункционирующие. Как и у некоторых ныне живущих мхов, устьица исходно служили для облегчения газообмена и транспорта воды (путем транспирации). Чаще всего они расположены в плоскости эпидермы, но у некоторых видов погружены глубоко.
Ареалы распространения мхов в значительной мере совпадают по характеру с ареалами цветковых растений. Распространение некоторых видов, ныне встречающихся по всему земному шару (Marchantía polymotpha, Bryum argenteum, Fuñaría hygmmetrica), возможно, обусловлено человеком. Мхи составляют в растительных сообществах с доминированием цветковых растений собственные сообщества подчиненного ранга (синузии, см. 14.3.4), нередко конкурируя с лишайниками. Собственные формации они образуют только в Арктике (тундре) и иногда также на верховых болотах, где продуктивность сплошного мохового покрова достигает своих максимальных значений: 200 — 900 г сухого вещества на 1 м2 в год; это соответствует урожаю сена на лугу среднего качества.
Наиболее распространены мхи как гигрофиты в областях с высокой влажностью: в лесах и на болотах. В целом печеночники более влаголюбивы, чем листостебельные мхи. Мхи чаще всего менее страдают от экстремальных условий, таких, как большая сухость, высокая температура и сильное излучение, и обходятся светом меньшей интенсивности, чем цветковые растения. Поэтому они проникают глубоко внутрь расселин и пещер, а также могут успешно расти на почве в лесах и в других затененных местах, особенно в форме войлока, дерновинок и корочек. Наибольшего разнообразия форм, в том числе повислых, метровой длины (см. рис. 11.126, S), и эпифитов, мхи достигают в тропиках, особенно в туманных и горных лесах. У них развиты самые разные варианты приспособлений для капиллярного удерживания воды. Удивительно много видов мхов заселяет поверхность листьев других растений. Такие эпифильные мхи в настоящее время рассматривают не как исключительно эпифиты, а иногда и как полупаразиты, которые своими ризоидами проникают под кутикулу несущего их листа и извлекают из него воду и соли.
Листостебельные мхи умеренной зоны (кроме эпифитов, а также мхов на почве и каменистом субстрате) часто проявляют ритмичность роста, четко связанную с временем года (см. рис. 11.126, В, Q). Реже они бывают однолетними, но чаще зеленые в течение всего года (вечнозеленые) и сохраняют, как облиственные печеночники, свои листья и зимой.
Ксерофитные мхи обладают высокой устойчивостью к высыханию, а также к высоким температурам и могут долгое время (Тоrtula muralis до 14 лет) находиться в воздушно-сухом состоянии, не теряя жизнеспособности. Споры, напротив, гораздо менее устойчивы. Находящиеся на открытом солнце и поэтому часто испытывающие сухость, мхи нередко растут низкими плотными дернинками и плотными подушками (см. рис. 11.126, Н). Зачастую они имеют серебристо-серый цвет, обусловленный длинными мертвыми окончаниями листьев. Такие «стекловидные волоски» (G), возможно, служат для защиты от избытка света и для снижения транспирации. Широкие однослойные края листьев у Polytrichumpiliferum способны заворачиваться над многослойной средней частью, несущей ассимиляционные пластинки, и таким путем защищают ее от высыхания (как скручивающиеся листья,см. 4.3.3.2). Мхи могут выдерживать экстремальные температуры: их можно встретить, с одной стороны, на скалах нивального пояса высокогорий, а также в Арктике и Антарктике, с другой стороны — на открытых местах, где температура почвы может доходить до 70 °С. В эксперименте некоторые высушенные до воздушно-сухого состояния листостебельные мхи выдерживали нагревание в течение 30 мин до 110°С, оставаясь живыми.
Несколько видов вторично приспособились к жизни в воде (гидрофиты), причем были утрачены как внешние, так и внутренние системы проведения воды.
Fontinalis antipyretica и другие водные мхи очень чувствительны к продолжительному пребыванию вне воды. Живущие в богатых известью ручьях и водопадах мхи (например, Eucladium verticillatum, Btyum pseudotriquetrum, Cratoneuron commutatum) вносят наряду с различными видами цианобактерий (Oocardium) и Chara существенный вклад в образование известковых туфов. Извлекая из воды диоксид углерода, они превращают растворенный гидрокарбонат в малорастворимый карбонат кальция, выпадающий в осадок.
Немногие мхи (например, виды Pottia) растут как галофиты по берегам морей и в засоленных местностях в глубине материков.
В полостях талломов Blasia (см. рис. 11.114, С) и Anthoceros (см. рис. 11.127, В) как симбионт живет синезеленая водоросль Nostoc. В ризоидах или клетках таллома либо стебля многих печеночников содержатся гифы гриба, однако в каждом отдельном случае бывает трудно решить, имеет ли здесь место паразитизм или же симбиоз по типу микоризы (см. 9.3.1). Бесхлорофилльный печеночник Ciyptothallus mirabilis, обитающий под ковриками листостебельных мхов, питается как паразит на гифах гриба, тогда как ризоиды Marchanda и других мхов могут, наоборот, поражаться паразитическими грибами.
Обзор мхов. На связь бриофитов и рассматриваемых дальше сосудистых растений с предшествующими Chlorobionta указывает много общих признаков, в том числе одни и те же фотосинтетические пигменты, образование крахмала в пластидах, наличие целлюлозы в клеточных стенках, а также сходное строение подвижных репродуктивных клеток. Мхи, вероятно, произошли от предков, которые соответствовали стрептофитам из Chlorobionta. Из них мхи ближе к харовым (Charophyceae) по двухжгутиковым асимметричным сперматозоидам и ряду различных ультраструктурных и биохимических признаков и, возможно, к Klebsormidiophyceae, чем к другим зеленым водорослям. Протонемы мхов, которые у отдельных форм могут непосредственно развивать гаметангии1 и у мно гих нитевидные, указывают на произошедший переход от нитчатого к тканевому таллому.
1 Гаметангии развиваются на гаметофорах, хотя бы и сильноредуцированных. — Примеч. ред.
Эволюционное развигие мхов, вероятно, шло с рубежа силура и девона параллельно развитию первых наземных папоротникообразных. Неясны родственные связи антоцеротовых, которые, скорее всего, развивались параллельно печеночникам и листостебельным мхам. Действительно ли они наиболее близки (из мхов) к высшим наземным растениям, остается спорным. Исходные формы мхов, как, по-видимому, и сосудистые растения, унаследовали проводящие структуры для воды и ассимилятов от общих предков. Об этом говорят большая степень сходства в строении и функции гидроидов и лептоидов мхов с трахеидами (сосудами) и ситовидными клетками папоротникообразных и наличие архегониев и устьиц в обеих группах. В ходе эволюции бриофитов гаметофит (зеленое растение мха) и, возможно, спорофит претерпели различные упрощения, но одновременно достигли биологического прогресса (прогрессивная редукция). Так, среди листостебельных мхов прямостоячие, высокорослые акрокарпные формы с хорошо развитой, напоминающей протостелу проводящей системой (Polytrichanae) рассматривают как исходные для лежачих, разветвленных плеврокарпных видов без проводящей системы и без жилки в листе. Наличие нефункционируюших устьиц можно трактовать также как результат регрессии. Прогрессивное развитие, напротив, претерпели клеточная сеть листьев (от паренхимной к прозенхимной) и структура устья коробочки (перистом).
Отдельные ископаемые мхи встречаются с верхнего девона; они дают мало информации для познания их происхождения. Sporogoniles из девона, правда, можно рассматривать как сомнительное связующее звено между мхами и примыкающими к ним папоротникообразными (Рsilophytopsida). Талломные и листостебельные печеночники, как и первые листостебельные мхи (Muscites), встречаются в карбоне Англии. Это свидетельствует о большом эволюционном возрасте бриофитов. Листостебельные мхи, которые можно отнести к Sphagnidae и Вrydae. были описаны из перми на территории России (Печора, Кузнецк). Листостебельные мхи нижнего карбона и перми имели жилки в листьях (в том числе и Protosphagnales), тогда как первые формы без жилок известны только из триаса и все более обычны в поздней юре. Большинство находок ископаемых мхов с возрастающей долей плеврокарпных известно из третичного периода. Они могут быть отнесены к современным родам.