БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
Четвертый подотдел; семенные растения (Spermatophytina)
Семенные растения (примерно 250000 видов) — самая большая современная группа растений. Они господствуют в наземной растительности почти во всех частях света и составляют основу всей наземной жизни. Для человека семенные растения имеют огромное значение, так как большинство культивируемых растений относятся именно к этой группе, в особенности к покрытосеменным.
Чередование поколений
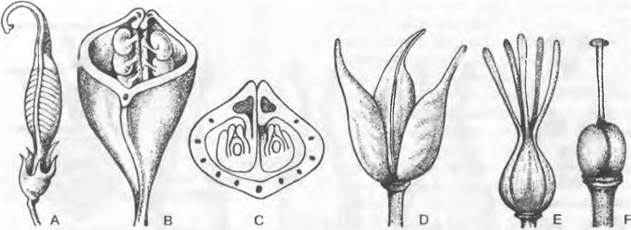
Пяти современным группам семенных растений (Gymnospermae: Cycadopsida, Соniferopsida, Ginkgopsida, Gnetopsida; Angiospermae: Magnoliopsida), как мхам и папоротникообразным, свойственно гетероморфное чередование поколений и диплогаплонтное чередование ядерных фаз с диплоидным спорофитом и гаплоидным гаметофитом (рис. 11.165). Как и у современных папоротникообразных, спорофит семенных растений состоит из корня и побега со стеблем и листьями. Гаметофит в сравнении с большинством современных папоротникообразных сильно редуцирован. Семенные растения вместе с мхами и папоротникообразными составляют группу эмбриофитов, а вместе с папоротникообразными относятся к кормофитам (Kormophyta — растения, имеющие побеги1), или трахеофитам (Тгаcheophyta — сосудистые растения).
1 Группа включает все мхи и большинство печеночников. — Примеч. ред.
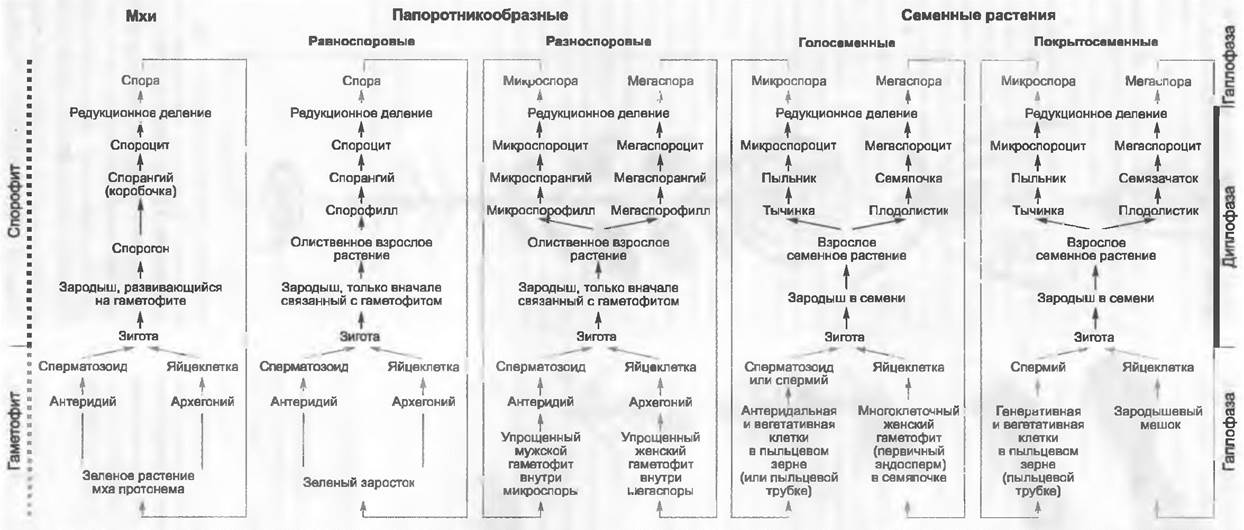
Лишь в 1851 г. W. Hofmeister обнаружил «тайное» чередование поколений у семенных растений и тем самым установил их тесную связь с мхами и папоротникообразными. Уже тогда в ботанике устоялись собственные названия для органов размножения семенных растений. И хотя их гомология с соответствующими органами папоротникообразных надежно установлена, до настоящего времени применяют обе группы понятий (рис. 11.164).
Рис. 11.164. Сравнение смен поколений и ядерных фаз у Embryophyta (Соrmobionta). Изображены схемы смен у мхов, равно- и разноспоровых папоротникообразных, а также у семенных растений. Гомологичные фазы развития, репродуктивные клетки и органы расположены на одном горизонтальном уровне (ср. также рис. 11.108; 11.128; 11.137; 11.165)
Как и отдельные папоротникообразные, семенные растения разноспоровые. В мегаспорангии (= нуцеллусе) чаще всего образуется только одна материнская клеткамегаспоры (= материнская клетка зародышевого мешка), из которой в результате мейоза получаются четыре мегаспоры. Обычно только одна мегаспора развивается в женский гаметофит (= зародышевый мешок). Женский гаметофит образует яйцеклетку2. В отличие от большинства папоротникообразных у семенных растений мегаспора не покидает мегаспорангий и соответственно материнское растение является спорофитом, и поэтому женский гаметофит развивается на материнском растении и за его счет. Кроме того, мегаспорангий семенных растений окружен стерильной оболочкой, интегументом, отсутствующим у папоротникообразных. Интегумент, нуцеллус и зародышевый мешок с яйцеклеткой в совокупности называют семязачатком1. В микроспорангии образуются многочисленные материнские клетки микроспор (= материнские клетки пыльцевых зерен), из которых после мейоза возникает множество микроспор. Мужской гаметофит (пыльцевое зерно) развивается внутри оболочки пыльцевого зерна и образует имеющие жгутики сперматозоиды или безжгутиковые спермин. В процессе опыления мужской гаметофит переносится к семязачатку, где он прорастает, образуя пыльцевую трубку, которая, удлиняясь, доставляет к яйцеклетке или подвижные сперматозоиды, или безжгутиковые, неподвижные спермин. В результате оплодотворения яйцеклетки сперматозоидом или спермием получается зигота, развивающаяся в зародыш. После оплодотворения семязачаток развивается в семя. Оно состоит из семенной кожуры, развивающейся из интегумента, многоклеточного зародыша и (почти всегда) питающей ткани (чаще всего эндосперма). Семя заменило одноклеточную спору папоротникообразных как средство распространения.
1 Семязачаток — это комплекс из нуцеллуса и интегумента (интегументов). — Примеч. ред.
2 У многих семенных растений женский гаметофит образует архегонии. — Примеч. ред.
Мега- или микроспорангии расположены поодиночке или группами на простых или более, или менее сложно разветвленных органах, которые можно назвать микроспорофиллами или мегаспорофиллами (тычинками и плодолистиками)1.
1 Тычинки и плодолистики свойственны только покрытосеменным растениям. — Примеч. ред.
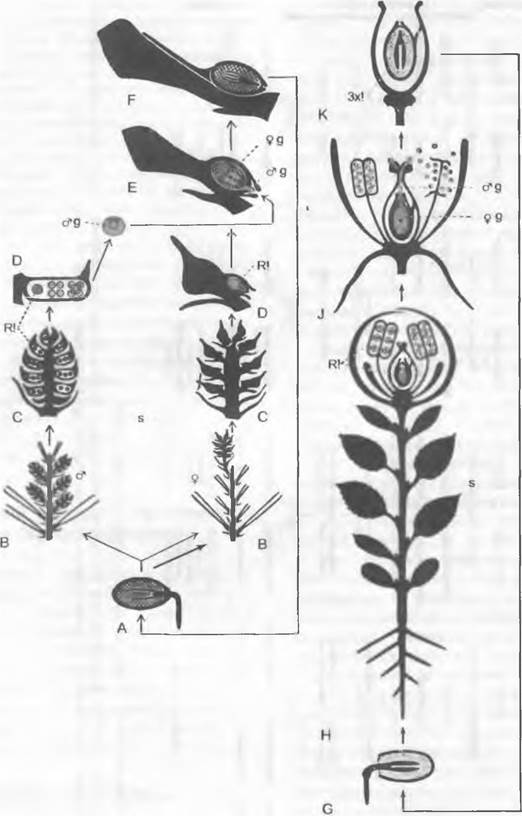
Рис. 11.165. Чередование поколений и ядерных фаз у голосеменных (Gymnospermae, Рinus/ Сoniferopsida, А — F) и покрытосеменных (G — К). Голосеменные: А — прорастающее семя с тестой, первичным эндоспермом (гаплоидным) и зародышем; В — побег со стеблем, листьями, а также ♂ и ♀ соцветиями* 1; С — ♂ стробил и ♀ констробил (молодая шишка); D — микроспорофилл с материнской клеткой микроспор, микроспорами и пыльцевыми зернами (воздушные мешки не показаны) как стадиями развития ♂ гаметофитов; кроющая чешуя шишки, над ней — семенная чешуя и открыто лежащий семязачаток, в котором развивается только 1 из 4 мегаспор; Е — семенная чешуя и семязачаток ко времени оплодотворения проросшим пыльцевым зерном, ♂ и ♀ гаметофит (g) с 2 крупными яйцеклетками; F — семенная чешуя зрелой шишки с (крылатым) семенем и зародышем в (первичном) эндосперме. Покрытосеменные: G — прорастающее семя; Н — целое растение с корнем, стеблем, листьями и бутоном обоеполого цветка; J — распустившийся цветок с околоцветником (чашелистики и лепестки), тычинками (с пыльцевыми зернами) и плодолистиками (завязь, содержащая семязачаток, столбик, рыльце) после опыления (пыльцевая трубка!) и перед самым оплодотворением яйцеклетки в зародышевом мешке; К — семя с тестой, вторичным (триплоидным) эндоспермом и зародышем, выпадающее из односемянного плода. Двойная штриховка: гаметофит (g) и первичный эндосперм; крап: триплоидный эндосперм; черный цвет: диплофаза (2n); светлый тон: гапло фаза (n); R! — редукционное деление (мейоз), s — спорофит
* Эти структуры правильно называть констробилами, а не соцветиями. — Примеч. ред.
Хотя термины «микроспорофилл» и «мегаспорофилл» подразумевают их гомологию с листьями, совершенно не ясно, принимали ли участие и в какой мере листья в образовании микро- и мегаспорофиллов у разных групп семенных растений. У некоторых современных групп растений они совсем не похожи на листья. Также не ясна гомология микро- и мегаспорофиллов разных групп семенных растений. Термин «плодолистик» следовало бы использовать только для обозначения особых структур покрытосеменных2.
2 Это сделано в мировой ботанике еще в первой половине XX в. — Примеч. ред.
Спорофиллы у Spermatophytina почти всегда расположены на укороченных побегах с ограниченным ростом. Такие структуры, имеющиеся уже у некоторых папоротникообразных, можно называть цветками3, но чаще всего это понятие используют только для первично обоеполых собраний спорофиллов покрытосеменных и предположительно близкородственных им ископаемых семенных растений.
3 Такое чрезмерно расширенное применение термина «цветок» вышло из употребления на рубеже XIX — XX вв. — Примеч. ред.
Вегетативные органы
Проростки семенных растений биполярны и имеют конусы нарастания первичного стебля и первичного корня4. Число зародышевых листьев (семядолей) обычно 2, но может увеличиваться или уменьшаться до одного. Апикальные меристемы стебля и корня с несколькими инициальными клетками дифференцированы на несколько зон (см. рис. 3.4; 3.6). Боковые корни образуются эндогенно. У некоторых групп, главным образом из покрытосеменных (например, у однодольных), главный корень рано прекращает рост. Его функции частично принимают на себя снабженные корнями подземные побеги (например, корневище), но в первую очередь — придаточные корни на надземных побегах. Проводящие пучки корня чаще всего образуют актиностелу1. Побеги нарастают моно- или симподиально, они либо однотипные, либо дифференцированы на удлиненные и укороченные. Ветвление в типе боковое, точнее — пазушное (= аксиллярное). В стебле чаще всего имеется эвстела или сифоностела из расположенных кольцом открытых коллатеральных проводящих пучков, реже — атактостела с равномерно распределенными по поперечному сечению стебля закрытыми проводящими пучками. Большинство семенных растений способны к вторичному утолщению, при котором камбий образует внутрь новые слои древесины, а наружу — луб. Однако способность к вторичному утолщению часто бывает утрачена или заменена аномальным вторичным утолщением. Соответственно среди семенных растений имеются как древесные, так и травянистые представители2. Листья расположены спирально, двурядно, накрест супротивно или мутовчато.
4 Такую структуру имеют сформировавшиеся зародыши. У проростков есть уже главный корень и почка главного побега. — Примеч. ред.
1 На стадии первичного строения в корне семенных растений развит единственный радиальный пучок. — Примеч. ред.
2 Большинство трав обладают типичным вторичным утолщением, хотя и незначительным. — Примеч. ред.
Важные систематические признаки дает анатомия побега. В ксилеме встречаются или только трахеиды, или наряду с трахеидами еще и сосуды. У сосудов можно наблюдать разный наклон разделяющих стенок и разную степень их разрушения при редукции перекладин лестничных перфорационных пластинок, а также различные укорочение и расширение члеников сосудов. Однако разнообразие этих признаков отражает не только родство, но и экологические особенности. Во флоэме у голосеменных находятся ситовидные клетки, а у покрытосеменных — происходящие из одной материнской клетки членики ситовидных трубок и клетки-спутники. У ситовидных трубок также можно наблюдать разнообразие степени наклона ситовидных пластинок, размеров ситовидных каналов в них, длины и диаметра самих члеников ситовидных трубок. Проводящие пучки могут быть не только коллатеральными, но и биколлатеральными благодаря образованию внутри от ксилемы еще одного тяжа флоэмы. Другие анатомические признаки стебля, используемые в систематике, — образование специализированных механических элементов (древесинных и лубяных волокон), взаимное расположение древесинной паренхимы и сосудов, пространственное разделение механических и проводящих тканей, а также степень однородности и радиальной вытянутости клеток лучей.
Весьма разнообразны также формы роста и жизненные формы семенных растений. У древесных форм иногда различают лептокаульную форму роста со сравнительно обильным ветвлением и тонкими ветвями (например, Сoniferopsida) и пахикаульную форму со слабым ветвлением и относительно толстым стволом (например, Суcadopsida). Из листопадных или вечнозеленых деревьев, или древовидных форм многократно независимо возникали, особенно у покрытосеменных, кустарники и полукустарники, лианы, многолетние травы, геофиты и эпифиты с самым разным расположением покоящихся органов и, наконец, однолетние травы. Однако древесные формы могут возникать и вторично из травянистых. Это явление особенно убедительно показано для разных островных флор. Большая вариабельность типов роста и жизненных форм покрытосеменных позволила им освоить значительно больше разных биотопов, чем голосеменным. В этом, пожалуй, важная причина видового богатства покрытосеменных.
Наряду с расположением листьев на стебле (спирально, двурядно, накрест супротивно, мутовчато) есть много других признаков листьев, используемых в систематике. Особенности расчлененности листьев: могут быть или отсутствовать прилистники и черешок, листовые пластинки бывают цельные или в разной степени расчлененные. Форма листовой пластинки или листочка сложного листа (общие очертания, форма верхушки и основания и т. п.) также дают много признаков, равно как и форма края листа или листочка, который может быть, к примеру, цельнокрайным или зубчатым (см. рис. 4.2). Наконец, таксономически константным признаком может быть и сложение листьев в почке. Форма листа варьирует и вследствие разной выраженности листового основания и пластинки (низовые и верховые листья, почечные чешуи и т. п.). Важна и анатомия листа. Так, листья бывают би- или унифациальными, значительно варьирует и характер жилкования листьев. Листья с вильчатым (дихотомическим) жилкованием есть, например, у Ginkgo, а у листьев с перистым жилкованием систематическое значение имеют ход и дальнейшее ветвление вторичных жилок, их возможное слияние друг с другом, вхождение в край листа и т. д. (см. рис. 4.60). У однодольных покрытосеменных часто встречается параллельное жилкование. Систематически значимы признаки анатомии зубцов листа (например, захождение жилок в зубцы, расположение в них механических тканей). Проводящие пучки стебля, в узлах продолжающиеся в проводящие пучки листьев, называю! листовыми следами1. Над местом отхождения листовых следов в проводящей системе стебля остаются так называемые листовые прорывы (лакуны). Число листовых следов и листовых прорывов изменчиво и таксономически значимо (рис. 11.166). Так, например, узлы бывают односледовые и однолакунные, трехследовые и однолакунные или трехлакунные, а также многоследовые и многолакунные.
1 Листовым следом нередко называют всю совокупность пучков, отходящих в один лист, а не каждый такой пучок по отдельности. — Примеч. ред.
Рис. 11.166. Листовые следы и листовые прорывы
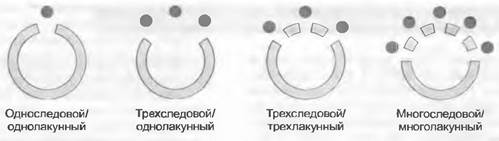
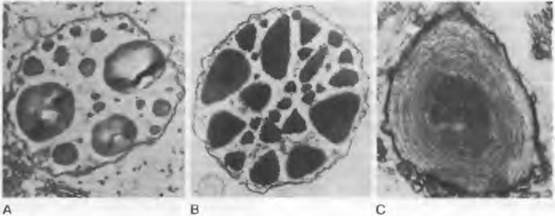
Рис. 11.167. Пластиды ситовидных трубок у покрытосеменных: А — S-тип с крахмальными включениями: Nuphar (Nymphaeaceae) (20 000х); В — Р-тип с кристаплоидными белковыми включениями: Gloriosa (Liliales: Colchicaceae); С — Р-тип с филаментозными белковыми включениями: Allenrolfea (Amaranthaceae) (В, С 30000х)
Другие примеры систематически значимых комплексов признаков вегетативных органов — это строение устьичных аппаратов и, если имеется, опушение. Устьичные аппараты можно, к примеру, классифицировать по строению око- лоустьичных, или побочных, клеток. Например, у аномоцитных устьичных аппаратов побочные клетки не отличаются от остальных клеток эпидермы, а у парацитных аппаратов замыкающие клетки окружены двумя побочными, лежащими параллельно замыкающим клеткам. Устьичные аппараты различаются также по развитию замыкающих и побочных клеток. Они могут происходить как от одной (мезогенные), так и от разных материнских клеток (перигенные). Волоски весьма разнообразны по числу слагающих клеток, их расположению (ветвистые или неветвистые волоски), относительному размеру, структуре стенок (жесткие, мягкие), окраске, наличию железок и т. п., что можно использовать в систематических целях (см. рис. 3.14). Систематическое значение также могут иметь идиобласты (например, кристаллоносные, масляные клетки), отличающиеся по строению окружающей однородной ткани.
Бокс 11.10. Семенные растения (Spermatophyta)
Возникшие скорее всего в самом начале карбона (более 360 млн лет назад) семенные растения (Spermatophyta) характеризуются прежде всего особенностями чередования поколений. Их исходно древовидные спорофиты разноспоровые и образуют микро- и мегаспоры, в которых эндоспорически развиваются гаметофиты1. Мегаспора с женским гаметофитом не покидает спорофит, а остается в невскрывающемся мегаспорангии, окруженном стерильным покровом (интегументом). Эти структуры (интегумент + мегаспорангий + ♀ гаметофит) вместе составляют семязачаток. Из семязачатка после оплодотворения яйцеклетки женского гаметофита развивается семя, давшее название всей этой группе растений. Семя исходно — единица распространения семенных растений, оно состоит из семенной кожуры (тесты), образовавшейся из интегумента, питающей ткани (эндосперма) разного происхождения и развившегося из зиготы зародыша (см. рис. 11.165). По сравнению с папоротникообразными (Pteridophyta) эволюционное новшество семенных растений, наряду с возникновением новой структуры — интегумента, состоит в том, что мегаспора больше не покидает спорофит и женский гаметофит сохраняет с ним физический контакт. Кроме того, женский гаметофит (♀) сильно редуцирован и иногда не образует архегониев. Также сильно редуцированный и не имеющий антеридиев мужской (♂) гаметофит остается внутри микроспоры и переносится на семязачаток в процессе опыления. Затем происходит оплодотворение яйцеклетки либо подвижным 2 жгутиковыми сперматозоидом, либо неподвижным безжгутиковым спермием.
1 Спорофиты образуют не непосредственно споры, а спорангии — Примеч. ред.
2 Семязачаток составляют мегаспорангий с интегументом/интегументами. — Примеч. ред.
Благодаря эволюции этих признаков оплодотворение у семенных растений стало совсем независимым от капельножидкой воды, а женский гаметофит и развивающийся на нем после оплодотворения зародыш снабжаются всем необходимым от материнского спорофита. Главным образом, именно в этих двух особенностях семенных растений следует видеть их преимущество среди современных наземных растений.
Гомология в чередовании поколений у мхов, папоротникообразных и семенных растений была установлена в 1851 г. W. Hofmeister. К этому времени для описания репродуктивных структур семенных растений уже разработали особую терминологию, которую используют и поныне (см. рис. 11.164). Так, мегаспорангий называют нуцеллусом, а женский гаметофит — зародышевым мешком. Для микроспоры с заключенным в ней мужским гаметофитом используют понятие пыльцевое зерно.
Ныне существующие семенные растения включают пять отдельных групп: саговники (Cycadopsida), хвойные (Coniferopsida), гинкговые (Ginkgopsida) и гнетовые (Gnetopsida) — эти четыре группы объединены в группу голосеменных (Gymnospermae), а также Magnoliopsida (покрытосеменные = Angiospermae). Их ближайшие родственники среди ныне живущих Pteridophyta — хвощи, псилотовые и папоротники.
Если семя в эволюции наземных растений, вероятно, возникло единственный раз, то семяподобные структуры возникали и у Lyco- podiopsida. Так, у карбонового рода Lepidocarроnмегаспорангий с мегаспорой и женским гаметофитом был окружен мегаспорофиллом и оставался на спорофите.
В систематике используют и ультраструктурные особенности, например, строение пластид ситовидных трубок и эпикутикулярного воска.
Применение трансмиссионной электронной микроскопии пластид ситовидных трубок позволило выделить разные их типы, иногда характерные для таксонов высокого ранга. Они различаются по наличию крахмальных и/или белковых включений, а также по форме и размеру последних (рис. 11.167). У голосеменных и покрытосеменных преобладает так называемый 5-тип пластид, содержащих только включения крахмала. Особый Р-тип с клиновидными белковыми кристаллами имеется преимущественно у однодольных покрытосеменных, а Р-тип с белковыми филаментами ограничен Сaryophyllales в узком смысле. Пластиды ситовидных трубок Р-типа могут содержать наряду с белком также и крахмал. Видимый в сканирующий электронный микроскоп эпикутикулярный воск может иметь вид пластинок, волосков или трубочек (см. рис. 3.11) и иногда характеризует определенные систематические группы.
Вторичные метаболиты
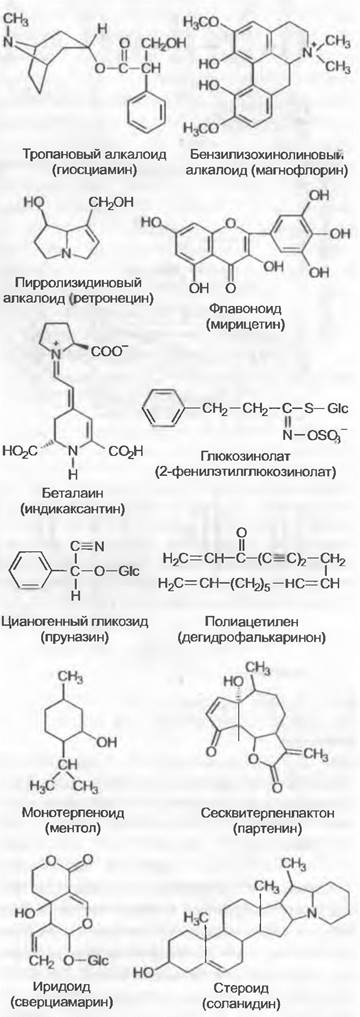
В систематике семенных растений, и особенно покрытосеменных, учитывают большое разнообразие так называемых вторичных метаболитов. Вторичными эти весьма различные соединения названы потому, что они не обязательны для осуществления основного обмена веществ растения. Важные классы веществ для систематики покрытосеменных (рис. 11.168) — это структурно очень гетерогенные алкалоиды (например, тропановые, бензилизохинолиновые, пирролизидиновые и т. п.), флавоноиды, беталаины, глюкозинолаты (гликозиды горчичного масла), цианогенные гликозиды, полиацетилены, терпеноиды (например, монотерпеноиды, сесквитерпенлактоны, стероиды, иридоиды и т. п.) и т. д. Если беталаины и антоцианы играют важную роль как пигменты прежде всего цветков, а другие вещества обусловливают запах цветков, то многие из названных классов веществ важны, вероятно, в основном как средства защиты от травоядных животных и патогенных организмов. Причиной видового многообразия покрытосеменных наряду с многообразием форм роста оказалось, вероятно, и богатство вторичными метаболитами.
Рис. 11.168. Важные для систематики покрытосеменных классы вторичных метаболитов
Упрощенно можно представить, что отчасти именно очень специфические взаимоотношения между растениями и травоядных и животными или патогенными организмами привели к ускорению процесса видообразования, например, в связи с тем, что возникновение нового растительного вещества, защищающего от травоядных, могло привести к появлению нового вида в новой экологической нише (без травоядных).
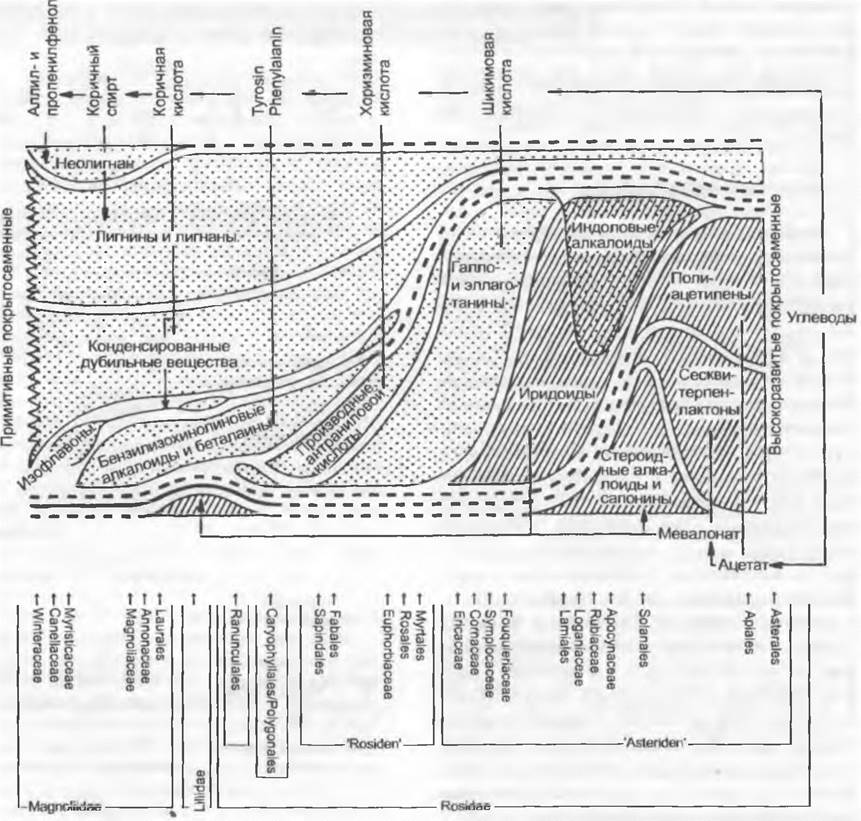
Важнейшие пути синтеза вторичных метаболитов — шикиматный и путь ацетатмевалоновой кислоты, причем шикиматный путь у более продвинутых таксонов всегда заменяется на путь ацетатмевалоновой кислоты (рис. 11 169).
Рис. 11.169. Упрощенная схема преобразования спектра накапливаемых биологически активных «вторичных метаболитов» у примитивных (слева) и продвинутых (справа) покрытосеменных. Производные шикиматного пути, среди них лигнин (показаны крапом), все больше заменяются дериватами пути ацетатмевалоновой кислоты (заштриховано). Участие ацетата в конденсации дубильных веществ и изофлавонов не рассмотрено
Цветки
Цветки — собрания спорофиллов, т. е. укороченные побеги с ограниченным ростом, несущие микро- и/или мегаспорофиллы. Хотя гомология спорангиофоров в разных группах семенных растений, как и гомология их листьев, никоим образом не доказаны, и в большинстве групп семенных растений микроспорангиофоры и мегаспорангиофоры не похожи на листья, их следует называть соответственно микро- и мегаспорофиллами. Среди современных семенных растений только Cicas имеет женские структуры, о которых нельзя говорить, как об укороченных побегах с ограниченным ростом. Здесь после образования многочисленных мегаспорофиллов на побеге вновь развиваются нормальные вегетативные листья. Цветки могут быть либо однополыми — только с микро- или только с мегаспорофиллами, либо обоеполыми — с микро- и мегаспорофиллами. Однополые цветки могут располагаться как на разных индивидуумах (двудомность), так и на одном — однодомность. Бывает, что обоеполые и однополые цветки встречаются на одном индивидууме в разных соотношениях (полигамия; см. 10.1.3.1).
Цветки семенных растений служат для полового размножения: это и образование мужских и женских гаметофитов на спорофитах, и участие в переносе пыльцевых зерен к семязачаткам или плодолистикам (опыление) и в самом процессе оплодотворения, а также забота о развитии зиготы в зародыш, семязачатка — в семя и, у покрытосеменных, завязи — в плод.
Околоцветник
Только Gnetopsida и Angiospermae имеют снаружи от спорофиллов стерильные, но функционально явно связанные с цветком листовые органы1. Функция околоцветника, с одной стороны, состоит в защите остальных элементов цветка на стадии бутона. С другой стороны, на стадии цветка он может играть важную роль в привлечении опылителей и выполнять другие функции, связанные с опылением, а у покрытосеменных также участвовать в распространении плодов.
У Gnetopsida покров мужских и женских «цветков» состоит из одной или двух пар чаще всего срастающихся хотя бы у основания брактей, которые иногда могут трубковидно срастаться (см. рис. 11.211). Их гомология с околоцветником покрытосеменных не ясна2.
1 У Gnetopsida цветков нет. — Примеч. ред.
2 Gnetopsida далеки от Angiospermae в таксономическом отношении, и едва ли имеет смысл искать гомологии их покровов с околоцветником цветковых растений. — Примеч. ред.
Покровы цветка покрытосеменных в целом называют околоцветником; он бывает весьма различной формы. Если все элементы околоцветника одинаковы, его называют простым,а отдельные листья — листочками простого околоцветника. Простой околоцветник может состоять из одного (монохламидный), двух или более кругов листочков или (при спиральном их расположении) из двух или более витков спирали (гомойохламидный). В цветках с неодинаковыми листочками околоцветника (двойной околоцветник, цветки гетерохламидные) наружные листочки, чаще всего зеленые, называют чашелистиками, составляющими чашечку (calyx). Внутренние, преимущественно яркоокрашенные листочки околоцветника — это лепестки, слагающие венчик. Общепринято, что листочки околоцветника могут иметь различное происхождение (рис. 11.170). С одной стороны, они могут развиваться из верховых листьев под цветком (обычно это признают за чашелистиками — см. рис. 4.6; переход от верховых листьев к листочкам околоцветника можно видеть, например, у Helleborus — см. рис. 4.5), а с другой стороны — из тычинок (обычно это приписывают лепесткам, и переходы между тычинками и лепестками можно найти, в частности, у Nymphaea).
Рис. 11.170. Переход от верховых листьев (А, В) к листочкам простого околоцветника (С) у Helleborus niger, переход от тычинок (К — F) к лепесткам (Е, D) у Nymphaea
Соответственно своей различной морфологической природе чашелистики и лепестки часто различаются и по снабжению проводящими пучками. В то время как в чашелистики входит обычно несколько, чаще всего три проводящих пучка, а сами они в целом анатомически сходны с листьями, в лепестки, как правило, входит только один проводящий пучок. В махровых цветках самых разных декоративных растений в качестве отклонения от нормы на месте тычинок образуются лепестки.
Элементы околоцветника покрытосеменных могут располагаться по спирали и/ или кругами. Простые околоцветники со спирально расположенными листочками можно с уверенностью считать исходными для покрытосеменных растений. Круговое расположение листочков околоцветника допускает их конгенитальное срастание. Пример сростнолистного простого околоцветника, по-видимому, Polygonatum, сростнолистной чашечки — часть Caryophyllaсеае или Fabaceae, а сростнолепестного венчика — большинство Asteridae. Элементы околоцветника могут «срастаться» и постгенитально, например, путем сцепления эпидерм папиллами или склеивания.
В спайнолепестном венчике можно различить базальную трубку и отгиб со свободными вершинами лепестков. Похожие части есть и в сростнолистном простом околоцветнике. В венчике можно различать раннюю симпеталию: образование трубки до или одновременно с вершинами лепестков и позднюю симпеталию: образование трубки после формирования вершин лепестков. Срастания в области околоцветника обеспечивают лучшую защиту или же лучшую пространственную интеграцию органов цветка в связи с особенностями опыления.
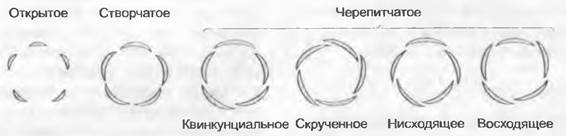
В систематике часто используют особенности сложения листочков околоцветника в бутоне (рис. 11.171). Они могут либо соприкасаться краями (створчатое сложение), различным образом перекрываться краем (черепитчатое) или вообще не контактировать между собой (открытое). В черепитчатом сложении можно выделить далее еще несколько типов: квинкунциальное (например, чашелистики у Rusa), скрученное (например, венчик Malva, Gentiana', в этом случае можно далее различать право- и левозакрученные варианты), а также кохлеарное (например, венчик Fabaceae с так называемым нисходящим сложением лепестков).
Рис. 11.171. Варианты сложения элементов околоцветника в бутоне. Листочки околоцветника в бутоне могут не соприкасаться (открытое сложение), касаться друг друга краями (створчатое) или различным образом перекрываться (черепитчатое)
Наряду с защитной функцией на стадии бутона и привлечением животных-опылителей благодаря особенно заметной окраске на стадии цветения, околоцветник (как чашечка, так и венчик) может участвовать и в самом процессе опыления, например, продуцируя и/или накапливая нектар, создавая посадочную площадку для опылителя, как в цветках губоцветных, или осуществляя вторичное «предложение» пыльцы. Однако в связи с измельчанием (например, при переходе к автогамии или при возникновении псевдантиев) цветков и при переходе к анеморфилии околоцветник может очень сильно упроститься или полностью редуцироваться в развитом цветке (ахламидные цветки). Остающиеся при плодах и иногда разрастающиеся чашелистики могут способствовать и распространению плодов.
Микроспорофиллы
Микроспорофиллы разных групп семенных растений сильно различаются по строению и расположению. Их гомология не ясна, и вопрос об участии листьев в их возникновении, как это подразумевается самим названием (микроспорофилл), остается открытым. Лучше было бы называть их нейтрально — микроспорангиофоры. У Ginkopsida микроспорофилл состоит из ножки с двумя висящими на ее кончике микроспорангиями (см. рис. 11.202). В мужском цветке1 микроспорофиллы расположены спирально на удлиненной оси. Мужские цветки2 современных Gonifero- psida шишковидные и состоят из обычно большого числа спирально или (реже) накрест супротивно расположенных микроспорофиллов. Отдельный микроспорофилл несет на нижней стороне от 2 до 20 часто сросшихся между собой микроспорангиев (см. рис. 11.203). Только у Taxus (см. рис. 11.210) микроспорангии расположены радиально на конце ножки микроспорофилла3. На нижней стороне чаще всего чешуевидных микроспорофиллов Cycadopsida (см. рис. 11.201) находится от 5 до 1 000 микроспорангиев, обычно расположенных отчетливыми группами по 3 — 5. Многочисленные микроспорофиллы в мужском цветке4 расположены спирально. У Gnetopsida микроспорофиллы расположены либо мутовками, либо терминально. Мутовчато расположены микроспорофиллы у Welwitschia, где 6 сросшихся между собой основаниями микроспорофиллов несут на концах по 3 сросшихся микроспорангия (см. рис. 11.211). В мужских цветках5Gnetum имеется только один терминальный микроспорофилл с нитевидным основанием и один или два верхушечными микроспорангиями (см. рис. 11.211), а у Ephedra терминально располагается чаще всего один, на конце обычно раздваивающийся нитевидный микроспорофилл с 2 — 8 группами микроспорангиев, почти всегда сросшихся по 2 (см. рис. 11.211).
1 У Ginkgopsida есть (мужские) микростробилы, а не цветки. — Примеч. ред.
2 Правильно — микростробилы. — Примеч. ред.
3 Это щитовидные микроспорофиллы, характерные также для Cupressaceae s.1 и Тaxodiaceaceь.s.1 — Примеч. ред.
4 У Cycadopsida цветков нет. — Примеч. ред.
5 Это на самом деле микроконстробил. — Примеч. ред.
Микроспорофиллы (тычинки) покрытосеменных (рис. 11.172) чаще всего дифференцированы на тычиночную нить и обычно терминально расположенный пыльник. Пыльник состоит из двух соединенных связником половинок — тек, каждая из которых содержит по два сросшихся микроспорангия.
Рис. 11.172. Тычинка покрытосеменных и детали ее строения. Общий вид тычинки у Hyoscyamus niger: А — спереди (адаксиально), В — сзади (абаксиально) (увеличено). Поперечные срезы пыльника Hemerocallis fulva: С — с еще не вскрывшимся, D — с уже вскрывшимся пыльником и проводящим пучком. Lilium pyrenacum: Е — поперечный срез стенки пыльника с эпидермой, фиброзным слоем, промежуточными слоями и остатками тапетума; отдельная клетка фиброзного слоя снаружи (F) и изнутри (G) (150х); Н, J — схема клетки фиброзного слоя до и во время сжатия; е — эпидерма; f — тычиночная нить, fs — фиброзный слой; к — связник; I — проводящий пучок; р — гнездо пыльника; t — тапетум; th — тека; z — промежуточный слой
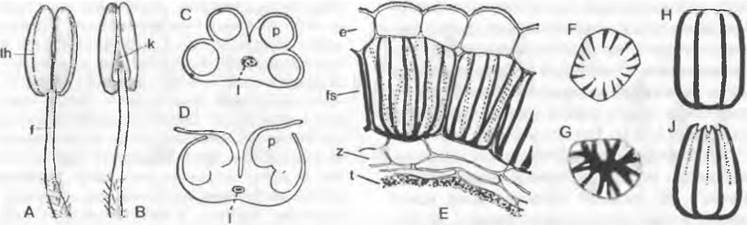
Тычинки покрытосеменных сравнительно редко отклоняются от этой базовой структуры. Микроспорофиллы бывают листовидно уплощенными и не дифференцированными на тычиночную нить и пыльник, но и в этом случае чаще всего отчетливо различимы две группы по два сросшихся микроспорангия. Пыльник может прикрепляться к тычиночной нити как основанием (базификсный), так и дорсальной (дорсификсный) или вентральной (вентрификсный) сторонами. Иногда число микроспорангиев в теке может быть уменьшено до одного. Благодаря поперечному и/или горизонтальному разделению микроспорангиев при сохранении в целом структуры теки могут возникать полиспорангиатные пыльники, как, например, у некоторых Rubiace и Rhizoprасеае. У Rafflesiасеае и Visсасеае дифференциация пыльников на теки утрачена, но пыльники точно так же полиспорангиатны: их многочисленные микроспорангии вскрываются по отдельности или все вместе через одну общую пору.
Число тычинок в цветке покрытосеменных варьирует от одной до примерно 2 000. Совокупность всех тычинок называют андроцеем. Тычинки могут располагаться по спирали или мутовчато, реже — более сложным образом или беспорядочно (рис. 11.173). В то время как при спиральном расположении чаще всего число тычинок большое и нефиксированное(первичная полиандрия), мутовчатое расположение тычинок обычно ведет к уменьшению и фиксации (олигомеризация) их числа. Цветок с двумя кругами тычинок — диплостемонный, с одним кругом — гаплостемонный. В цветках с исходно определенным числом тычинок в одном или немногих кругах могло произойти увеличение числа тычинок— вторичная полиандрия — благодаря разделению зачатков тычинок. При этом сначала ясно различимые отдельные зачатки могут вычленять многочисленные зачатки тычинок центробежно (рис. 11.173) или центростремительно, либо центробежно или центростремительно разделяется единый кольцевой зачаток. Благодаря расщеплению тычинок во время развития могут сформироваться «половинные» тычинки. Наконец, тычинки могут превращаться в стерильные стаминодии, которые служат как нектарники или уподобляются лепесткам для привлечения опылителей. Иногда тычинки в одном цветке устроены по-разному (гетерантерия). Мутовчато расположенные тычинки могут срастаться чаще своими нитями, но иногда и боками пыльников (синандрия) или постгенитально склеиваются между собой. Сериальное срастание тычинок и лепестков или тычинок и плодолистиков также возможно.
Рис. 11.173. Ранние стадии развития цветков покрытосеменных (околоцветник удален) с многочисленными зачатками тычинок: А — первичная поландрия у Magnolia denúdala(Magnoliaceae), зачатки расположены спирально на конусовидном цветоложе; В — вторичная полиандрия у Stewartia pseudocamellia (Theaceae), центробежное заложение многочисленных зачатков тычинок на пяти расположенных между зачатками лепестков и плодолистиков секторах блюдцевидно вогнутого цветоложа
Среди архаичных покрытосеменных наиболее широко распространено спиральное расположение тычинок, число которых варьирует.
Стенка пыльника, окружающая археспорий (ткань, образующую пыльцу1), всегда многослойна. Функционально особенно значимы слой клеток, ответственный за вскрывание пыльника, а также самый внутренний слой — тапетум. У Coniferopsida, Су- cadopsida и Gnetopsida вскрывание микроспорангия обеспечивает самый наружный слой стенки — экзотеций (см. рис. 11.201), а у Ginkgopsida и Angiospermae — субэпидермальный эндотеций (фиброзный слой, см. рис. 11.172), который иногда может отсутствовать, как, например, у Ericaceae. Как в экзо-, так и в эндотеции утолщены антиклинальные и внутренние периклинальные стенки клеток, а их наружные периклинальные стенки остаются тонкими. Как и в колечке спорангиев папоротников (см. рис. 8.37), потеря воды клетками экзо/эн- дотеция приводит к тангенциальному укорочению их наружных стенок. Вследствие этого стенка микроспорангия разрывается чаще всего в виде продольной щели (стомиум) в строго определенном месте. В эндотеции покрытосеменных антиклинальные стенки клеток несут гребневидные утолщения, которые часто становятся толще к внутренней стенке, где и сливаются. У покрытосеменных два микроспорангия одной теки чаще всего вскрываются одной общей продольной щелью, возникающей после разрушения клеток перегородки, разделявшей прежде полости микроспорангиев.
1 Археспорий образует спороциты, в холе мейоза которых формируются споры — Примеч. ред.
В зависимости от ориентации шели относительно центра цветка можно различать пыльники, вскрывающиеся интрорзно (внутрь), экстрорзно (наружу) и латрорзно (вбок).
Есть также пыльники, в которых каждый микроспорангий вскрывается собственной продольной щелью (например, Strelitzia), пыльники, вскрывающиеся порами при растворении ткани стенки в определенных местах — поры могут возникать, однако, и как очень короткие продольные щели (например, у Ericaceae) — или фиброзный слой развивается только в ограниченной области, которая отворачивается наружу в виде клапана (например, Lauraceae). Клапан может образовываться и благодаря раздвоению продольной щели на обоих концах. Иногда утолщения клеточных стенок фиброзного слоя имеют противоположную ориентацию, и микроспорангии при высыхании сжимаются (например, Welwitschia и Агасеае), выталкивая пыльцу наружу через отверстие стенки.
Экзотеций или эндотеций отделены от тапетума по меньшей мере одним промежуточным слоем, у покрытосеменных временным (рис. 11.172). Тапетум состоит чаще всего из богатых протоплазмой клеток с обычно эндополиплоидными ядрами и участвует в питании пыльцевых зерен и образовании веществ, откладываемых в стенки пыльцевых зерен1 либо на их поверхность (например, пыльцевой клей; вещества, важные для реакций самонесовместимости). Секреторный тапетум долго остается интактным, в то время как периплазмодиальный тапетум после растворения стенок его клеток и слияния их протопластов может амебоидно проникать между развивающимися пыльцевыми зернами.
1 Правильно — микроспоры. — Примеч. ред.
Хотя главная функция микроспорофиллов — образование пыльцы, они могут иметь и дополнительные функции, связанные прежде всего с опылением. Например, они могут сделать цветки более заметными или даже быть единственным их визуальным аттрактором. Микроспорофиллы могут продуцировать запахи, привлекающие опылителей, выделять нектар или определенным образом ограничивать движения опылителей в цветке, повышая таким образом эффективность процесса опыления. Наконец, они являются важными компонентами различных механизмов вторичного предложения пыльцы.
Из археспория возникает множество материнских клеток пыльцы, из которых после мейоза получается по четыре одноядерных пыльцевых зерна2. Если все клеточные стенки образуются одновременно, то говорят о симультанном способе образования микроспор. При сукцессивном способе образования микроспор первая клеточная стенка образуется уже после первого деления мейоза3.
2 Правильно — микроспор. — Примеч. ред.
3 Эти типы различают не по последовательности образования стенок, а по последовательности цитокинезов. — Примеч. ред.
Возникающие четыре микроспоры чаще всего располагаются в тетраэдрической тетраде (при симультанном типе образования). Но существуют и тетрады, в которых микроспоры располагаются в ряд (линейно), в одной плоскости (изобилатерально), в двух взаимно-перпендикулярных плоскостях (крестообразно) или Т-образно (при сукцессивном типе образования).
Пыльца
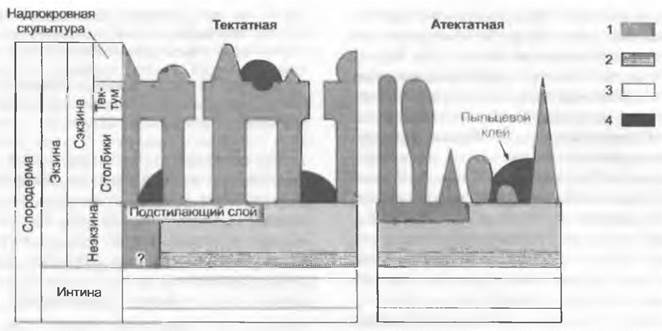
При переносе пыльцевых зерен по воздуху из микроспорангиев к женским репродуктивным органам они часто довольно долго подвергаются воздействию экстремальных условий, поэтому для размножения чрезвычайно важна защита их содержимого. Она осуществляется благодаря особому строению оболочки пыльцевого зерна, спородермы, состоящей из двух комплексных слоев — наружной экзины и внутренней интины (рис. 11.174).
Рис. 11.174. Схема тонкого строения стенки пыльцевых зерен покрытосеменных разных типов, схематизированный рисунок: 1 — эктэкзина, 2 — эндэкзина, 3 — интина, 4 — пыльцевой клей
Интина, сплошным слоем окружающая протоплазму, чаще всего нежная и химически мало устойчивая. Обычно интина состоит из двух-трех слоев. Самый наружный из них часто содержит много пектинов, что позволяет ей легко отделяться от экзины. Основной компонент внутреннего слоя/слоев — фибриллы целлюлозы. При прорастании пыльцевых зерен только интина разрастается в пыльцевую трубку1.
1 В пыльцевую трубку разрастается сифоногенная (вегетативная) клетка мужского гаметофита, покрытая интиной. — Примеч. ред.
Экзина образована в основном химически устойчивыми спорополленинами. Это терпены, которые, как полагают, возникают в результате оксидативной полимеризации из каротиноидов и их эфиров. Основной конструктивный элемент экзины — гранулы размером около 6 нм.
У голосеменных экзина дифференцирована на внутреннюю эндэкзину, имеющую пластинчатую (ламеллярную) структуру, и наружную эктэкзину, которая состоит из внутреннего подстилающего слоя, компактного наружного слоя и гранулярного или альвеолярного среднего слоя. Эндэкзина покрытосеменных не ламеллярная, а гранулярная2. Эндэкзину вместе с самым внутренним слоем эктэкзины — плотным и гомогенным подстилающим слоем — у покрытосеменных (рис. 11.174) также называют нэкзиной. Называемая сэкзинойнаружная часть эктэкзины покрытосеменных чаще всего сильно структурирована и скульптурирована. У атектатных пыльцевых зерен сэкзина представлена только столбиками в виде палочек, булав, конусов, бородавок или сетчатой структурой на нэкзине. Столбики могут сливаться дистальными концами в дополнительный наружный слой, покров (тектатные пыльцевые зерна). Тектум может быть пронизан порами самой различной формы, сам бывает многослойным и скульптурированным снаружи. В полостях тектума могут откладываться протеины самонесовместимости, пыльцевой клей и т. п. Очень редко, к примеру, у опыляемой под водой морской травы Zostera, экзины нет. У Pinaceae могут возникать воздушные мешки за счет локального расхождения наружного и внутреннего слоев экзины (см. рис. 11.203).
2 У очень многих покрытосеменных эндэкзина ламеллярная. — Примеч. ред.
Электронно-микроскопические исследования развития пыльцевых зерен показывают, что еще внутри каллёзной оболочки материнской клетки микроспор, на плазмалемме микроспоры откладывается тонкий слой фибриллярного материала— примэкзина1. Затем в нем появляются более компактные элементы, которые, удлиняясь и утолщаясь, становятся столбиками, расширяются на обоих концах и образуют таким образом тектум и подстилающий слой эктэкзины. Внутри от подстилающего слоя затем возни кает неламеллятная эндэкзина и, наконец, инти на. Таксоноспецифическое строение экзины кон тролируется самим пыльцевым зерном, но сильно зависит от тапетума спорофита. Это, среди прочего, можно видеть по реакции спорофит ной самонесовместимости (см. 10.1.3.1).
1 Примэкзина часто бывает везикулярной. — Примеч. ред.
Пыльцевые зерна сильно различаются по форме, расположению и числу апертур (рис. 11.175). Бывают пыльцевые зерна без апертур (безапертурные). Обращенный к центру тетраэдрической тетрады полюс пыльцевого зерна называют проксимальным, направленный наружу — дистальным. Экваториальная плоскость перпендикулярна к оси, связывающей оба полюса. У пыльцевых зерен семенных растений бывают только дистальные, только экваториальные или разбросанные по всей поверхности апертуры. Встречающиеся у Pteridophyta проксимальные апертуры здесь не известны2. Продольно вытянутые апертуры называют бороздами (соответственно дистальной — sulcus или экваториальной — colpus), округлые — порами (дистальной — ulcus или экваториальной — porus).
2 У Рteridophyta есть (микро)споры, а не пыльцевые зерна. — Примеч. ред.
Рис. 11.175. Некоторые обычные типы пыльцевых зерен семенных растений Центральной Европы. Слева в каждом случае вид с дистального полюса, справа — вид с экватора Монады (одиночные зерна): дистально-однобороздное (многие Magnoliopsida, Liliopsida), однопоровое (Роасеае), трехбороздное (часть Ranunculaceae, Quercus, Acer, Brassicaceae, Salix, часть Lamiaceae), трехпоровое (Betula, Corylus, Urticaceae, Onagraceae), трехборозднооровое (Fagus, часть Rosaceae, Apiaceae, Tilia, Asteraceae), стефанокольпатное (Rubiaceae, часть Lamiaceae), стефано-юратное (Alnus, Ulmus), пантопоратное (Juglans, большая часть Caryophyllaceae, Amaranthaceae, Plantaginaceae), безапертурное (Populus, Callitriche) из таксонов в целом с экваториально-апертурными пыльцевыми зернами. Тетрады: у таксонов, где встречаются и монады (часть Orchidaceae, часть Typha, Ericaceae)
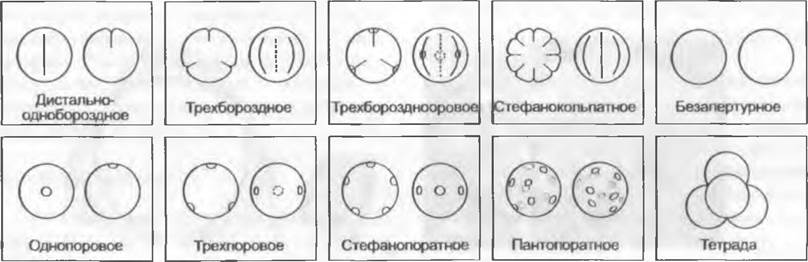
У голосеменных, пыльцевые зерна чаще всего дистально-однобороздные. Такие пыльцевые зерна находят также среди покрытосеменных у большинства Magnoliidae и у однодольных, где, однако, имеются так же однопоровые и безапертурные пыльцевые зерна. Большая группа настоящих двудольных характеризуется первично трехборозднымипыльцевыми зернами, борозды которых перпендикулярны экваториальной плоскости. Таким образом, исходны для покрытосеменных дистально-однобороздные пыльцевые зерн... Борозды могут быть заменены порами, например, у трехпоровых пыльцевых зерен. Если в экваториальной плоскости встречается более трех борозд или пор, то говорят о стефанокольнатных или стефанопоратных пыльцевых зернах. Апертуры, рассеянные по всей поверхности пыльцевого зерна (глобально-апертурные, например, пантопоратные), имеются, к примеру, у Cactaceae или Caryophyllaceae. При этом число апертур может возрастать до 100 (например, у Amaranthaceae). Пыльцевые зерна с апертурами, состоящими из борозд и пор, называют борозднооровыми. Вследствие изменчивости края апертур или образования утолщенного оперкулума могут возникать очень сложно устроенные апертуры.
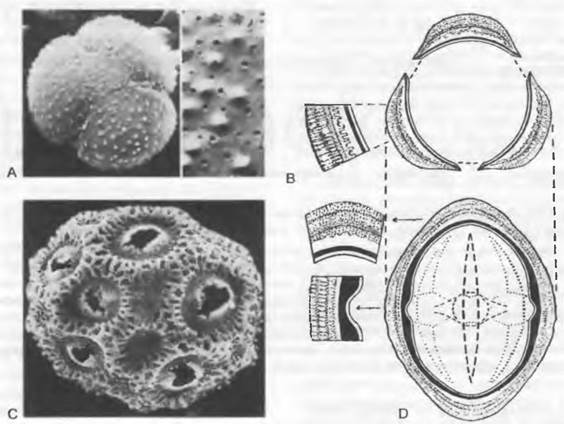
Невероятное многообразие типов пыльцевых зерен оказалось возможным классифицировать с помощью NPC-системы: по числу (Numerus), положению (Positio) и типу (Charakter) апертур. Кроме того, систематическое значение имеют еще многие различия в симметрии, форме и размере пыльцевых зерен, а также в тонкой структуре их экзины. Структуру пыльцевых зерен можно показать на так называемых палинограммах или на электронно-микроскопических фотографиях (рис. 11.176; см. бокс 4.1, рис. F).
Рис. 11.176. Пыльцевые зерна разных кактусов при сканирующей электронной микроскопии: А, В — Gymnocalycium mthanovichii (трехбороздное, внешний вид: 500х; детали строения остробородавчатого и пронизанного порами тектума: 5000х); С — Opuntia sp. (пантопоратное: 1000х); D — палинограмма пыльцевого зерна Centaurea scabiosa(трехборозднооровое): вид с экватора, оптическое сечение и детали структуры оболочки (световая микроскопия, 1 500х и 3000х)
Во время роста пыльцевых зерен тапетум образует особую клейкую субстанцию, содержащую липиды и каротиноиды, — пыльцевой клей. У опыляемых животными видов он откладывается прежде всего на поверхности пыльцевых зерен, что приводит к склеиванию их в комочки и способствует прикреплению к опылителю.
Функцию склеивания пыльцевых зерен выполняют и висциновые нити, которые чаще всего образуются внутри микроспорангия и содержат спорополленин, целлюлозу или протеины. Наконец, бывает и так, что пыльцевые зерна склеиваются секретами других элементов цветка. С другой стороны, приспособления для склеивания пыльцевых зерен могут отсутствовать, например, у ветроопыляемых семенных растений.
Пыльцевые зерна одной тетрады вовсе не всегда распространяются только в виде монад, т. е. по отдельности. Наряду со слипанием зерен благодаря пыльцевому клею или висциновым нитям микроспоры одной материнской клетки могут длительное время оставаться соединенными в тетраду и в таком виде распространяться (например, у Ericaceae, Drosera и др.). Путем прогрессирующей редукции трех клеток одной тетрады могут возникать и псевдомонады (например, у Сурегасеае). Если остаются объединенными пыльцевые зерна, образовавшиеся из нескольких материнских клеток, то получаются полиады, которые могут состоять из 8, 16 или 32 пыльцевых зерен (например, у Mimosoideae). Наконец, и все пыльцевые зерна одного микроспорангия могул объединяться в поллиний, а зерна двух или нескольких микроспорангиев — в поллинарий; обычно они покрыты общей спорополлениновой оболочкой (например, у некоторых Apocynaceae, Orchidaceae; см. рис. 11.274 и 11.227).
Мужской гаметофит
Мужской гаметофит семенных растений состоит из очень немногих клеток и никогда не имеет морфологически различимых антеридиев. Его развитие проходит в основном внутри оболочки пыльцевого зерна и лишь частично уже после его прорастания. У голосеменных, к примеру, у Pinaceae и Ginkgo, путем двух неравных делений образуются две проталлиальные клетки и одна антеридиальная клетка, которая делится на сифоногенную клетку и генеративную клетку. Из генеративной клетки образуются клетка ножка и сперматогенная клетка, а из последней — два сперматозоида или два спермия (рис. 11.177). Перед делением сперматогенной клетки мужской гаметофит, таким образом, пятиклеточный (2 проталлиальные клетки, клетка пыльцевой трубки, клетка-ножка и сперматогенная клетка; см. рис. 11.204). Есть немало отклонений от этого типа. Число проталлиальных клеток может возрастать до 8 (Podocarpaceae) или до 40 (Araucariaceae), или же она только одна (Cycadopsida; рис. 11.178), либо их совсем нет (Taxys, Gnetum, Welwitschia, Angiospennae). Сифоногенная клетка развивается у всех семенных растений, а клетка-ножка у покрытосеменных отсутствует. У Microcycas (Cycadopsida) увеличено число половых клеток, так как клетка-ножка образует свыше 20 сперматозоидов.
Рис. 11.177. Развитие ♂ гаметофитов от микроспоры до образования спермиев/сперматозоидов у Pinaceae и Ginkgo
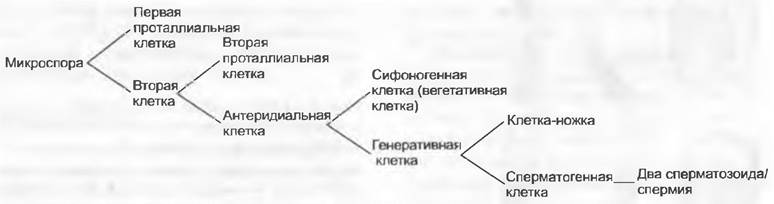
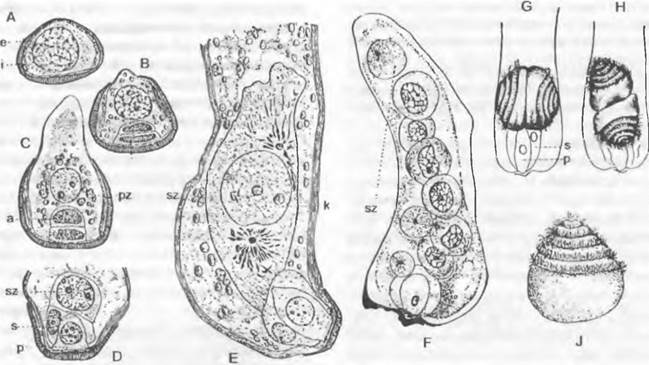
Рис. 11.178. Развитие ♂ гаметофитов у Cycadopsida: A — E — прорастание пыльцевого зерна y Dioon edule (A — C — 840x; D — 667x; E — 420x); F — проросшее пыльцевое зерно Microcycas calocoma c 9 сперматогенными клетками (около 200х); G — J — пыльцевая трубка и сперматозоид Zamia floridana (G, H — 50x, J — 75x); a — антеридиальная клетка; e — экзина; i — интина; k — ядро; p — протаплиальная клетка; pz — сифоногенная клетка, s — клетка-ножка; sz — сперматогенная клетка
Мужской гаметофит покрытосеменных, поскольку у него нет ни проталлиальных клеток, ни клетки-ножки, состоит только из сифоногенной клетки, которую здесь называют вегетативной, и второй, называемой генеративной, клетки, из которой образуются два спермия (рис. 11.179). Деление генеративной клетки может происходить перед или после прорастания пыльцевого зерна, время этого деления коррелирует с типом системы самонесовместимости (см. 10.1.3.1).
В любом случае пыльцевые зерна прорастают пыльцевой трубкой. Ее функция, с одной стороны, — доставить мужские половые клетки к яйцеклеткам, но она служит также для заякоривания мужского гаметофита и как гаустория для поглощения питательных веществ, необходимых для развития и роста самой пыльцевой трубки (рис. 11.180). В качестве половых клеток у Cycadopsida и Ginkgopsida образуются подвижные многожгутиковые сперматозоиды (см. рис. 11.178; 11.180; 11.202), a y Coniferopsida, Gnetopsida и Angiospermae— неподвижные спермин (см. рис. 11.204; 11.179).
Рис. 11.179. Развитие ♂ гаметофита у покрытосеменного растения LiUum martagón. Вегетативная клетка с ядром (к) и генеративная клетка (g) в пыльцевом зерне (А, В) и пыльцевой трубке (С). В переднем конце пыльцевой трубки (D) генеративная клетка разделилась на два спермия (s)
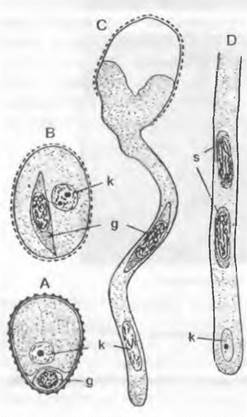
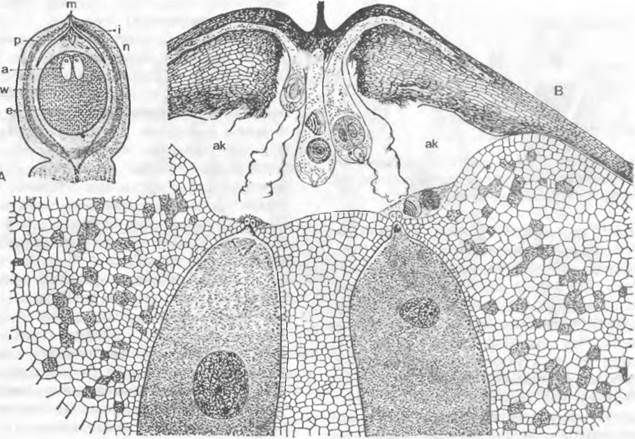
Рис. 11.180. Семязачаток и оплодотворение у Cycadopsida: А — продольный разрез семязачатка Ceratozamia с микрoпиле (m), интегументом (i). нуцеллусом (n) и пыльцевой камерон (р) с прорастающими пыльцевыми зернами; проросшая мегаспора: ♀ гаметофит (= зародышевый мешок, е) со стенкой (w) и двумя архегониями (каждый из двух клеток шейки и одной яйцеклетки) (2,5х); В — верхняя часть нуцеллуса ко времени оплодотворения у Dioon edule, пыльцевые трубки заякорились в ткани нуцеллуса и проникли в архегониальную камеру (ак), часть сперматозоидов высвобождена, левый из двух архегониев (а) уже оплодотворен (около 100х)
Мегаспорофиллы
Семязачатки семенных растений расположены в цветках1 очень разного вида. Хотя до сих пор точно не известны ни гомологии семяносных структур у разных групп семенных растений, ни участие листьев в их возникновении, за ними закрепилось название мегаспорофиллы вместо нейтрального мегаспорангиофоры. Понятие плодолистик традиционно применяют для обозначения мегаспорангиофора покрытосеменных растений.
1 Цветки есть только у покрытосеменных. Семенные органы других растений нельзя называть цветками! — Примеч. ред.
Несомненно, на листьях располагаются семязачатки у саговниковых (Cycadopsida) (см. рис. 11.200). У них чаще всего два семязачатка расположены на нижнем крае листовой пластинки черешкового, чешуевидного или щитовидного мегаспорофилла. До 8 семязачатков развивается на рахисе явно перистого (или по меньшей мере зубчатого) на верхушке мегаспорофилла у представителей рода Cycas. У современных хвойных (Coniferopsida) бывает от одного до 20 семязачатков на верхней стороне уплощенной семенной чешуи (см. рис. 11.205). Реже семязачатки кажутся расположенными на верхушках укороченных побегов (например, у Taxus; см. рис. 11.210). Исходя из положения семенной чешуи в пазухе кроющей чешуи, а также из сравнения с ископаемыми родичами (см. рис. 11.206; 11.207) современных хвойных семенные чешуи следует считать модифицированными укороченными побегами. Не ясно, находились ли исходно семязачатки на листьях этих укороченных побегов или на оси. У Ginkgo два семязачатка расположены на концах вильчато разветвленной ножки (см. рис. 11.202), а у родов гнетовых (Gnetopsida) каждый цветок имеет только один терминальный семязачаток (см. рис. 11.211).
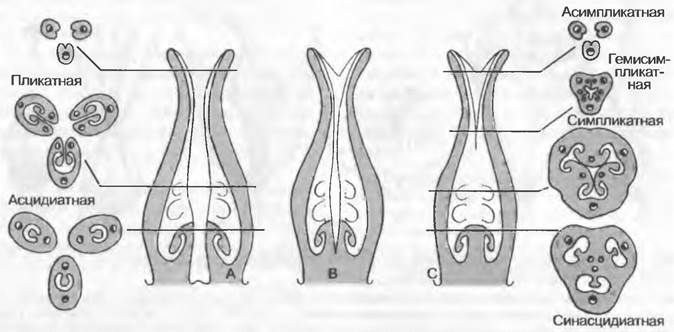
В то время как у вышерассмотренных групп, объединяемых как голосеменные (Gymnospermae), семязачатки расположены открыто, у покрытосеменных (Angiospermae) они заключены в плодолистики. Совокупность всех плодолистиков цветка покрытосеменных с содержащимися в них семязачатками называют гинецеем1. Структуру отдельного плодолистика можно понять по его развитию. При этом на ранних стадиях чаше всего возникает седловидный зачаток (рис. 11.181), более низкий край которого, называемый поперечной зоной, обращен к центру цветка. В течение некоторого времени края зачатка растут вверх совместно, благодаря чему возникает трубковидная (асцидиатная) зона (рис. 11.182). Если зачаток прекращает свой рост на внутренней стороне и далее растет только боками и спинной стороной развивающегося плодолистика, то образуется открытая в середину цветка брюшной щелью зона, называемая пликатной, или кондупликатной. Над этой областью расположен обычно стеблевидный столбик2, на котором располагается преимущественно покрытое сосочками рыльце для улавливания пыльцы. Столбик, как правило, не сплошной, а содержит чаще всего очень узкое продолжение полости асцидиатной и пликатной зон — так называемый трансмиссионный канал, по которому к семязачаткам различным образом попадают пыльцевые трубки. В развившемся таким образом плодолистике имеются снизувверх: ножка плодолистика. с самого сначала, т. е. конгенитально, замкнутая асцидиатная зона и первоначально открытая вентральной щелью и лишь постгенитально замыкающаяся (с помощью секрета и/или благодаря сцеплению эпидермальных зубчиков) пликатная зона. Область полого плодолистика, содержащую семязачатки, называют завязью.
1 Наличие внутри плодолистика(ов) семязачатков не является критерием гинецея. Гинецей остается гинецеем, даже если он полностью или частично состоит из стерильных плодолистиков. — Примеч. ред.
2 На самом деле — стилодий. — Примеч. ред.
Рис. 11.181. Схема этапов (А — С) развития типичного плодолистика покрытосеменных: вид спереди и продольный срез (светлый)
Рис. 11.182. Схема строения гинецеев. Продольные (А, В, С) и поперечные (А, С) срезы: А — апокарпного, В — гемисинкарпного, С — ценокарпного гинецея с асцидиатной, пликатной и соответственно синасцидиатной, симпликатной, геми- симпликатной и асимпликатной зонами
Относительные размеры асцидиатной и пликатной зон бывают весьма различными. Если в крайнем случае выше ножки образуется только асцидиатная зона, то плодолистик полностью асцидиатный. Если трубчатая зона отсутствует и образуется только пликатная зона, то плодолистик полностью пликатный (кондупликатный).
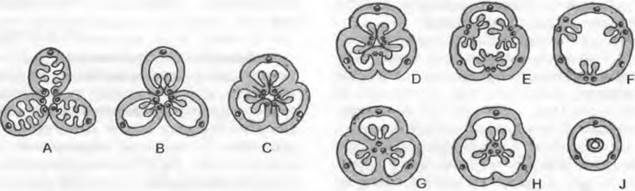
Плаценты — участки внутренней поверхности плодолистика, на которых находятся семязачатки. При этом плоские или выпуклые плаценты чаще всего находятся вблизи края вентральной щели — субмаргинально. Здесь они могут располагаться или О- образно вокруг всей вентральной щели (очень редко) или быть U-образными, находиться только у боковых краев (латерально) или у нижнего края вентральной щели (медиально). Кроме того, семязачатки могут также располагайся по всей внутренней поверхности завязи (ламинально; рис. 11.183). Число семязачатков в плодолистике варьирует от одного до нескольких миллионов.
Рис. 11.183. Разные типы гинецеев, поперечные срезы через фертильную главную зону выросших завязей: А — апокрапный гинецей, ламинальная плацента (П); В — апокарпный гинецей, субмаргинальная П; С — гемисинкарпный гинеций, центрально-угловая П; D, G — синкарпный гинецей, центрально-угловая П (D — плодолистики пликатные, G — плодолистики асцидиатные); Е, F — паракарпный гинецей, париетальная П; Н, J — паракарпный гинецей, центральная П; многочисленные семязачатки или один базальный семязачаток
В цветке бывает от одного до примерно 2 000 плодолистиков. Они могут располагаться спирально или мутовками. Как и в случае микроспорофиллов, при спиральном расположении плодолистики многочисленные, а число чаще всего не постоянно; мутовчатое расположение связано с уменьшением (олигомеризацией) и постоянством числа плодолистиков.
Если плодолистики не срастаются между собой, то гинецей называют апокарпным. Когда плодолистики срастаются — что, как правило, происходит при мутовчатом их расположении, — гинецей называют ценокарпным. Ценокарпный гинецей называют также пестиком1. Степень срастания плодолистиков может быть различной; к примеру, только основаниями завязей, всеми за вязями или также и стилодиями (рис. 11.184). Обычно число плодолистиков ценокарпного гинецея можно установить по числу рылец, но даже и рыльца могут срастись в единую структуру. Значительно варьирует и внутренняя структура ценокарпных гинецеев (см. рис. 11.183). Если у плодолистиков полностью развиты боковые и дорсальный участки и гинецей сооветственно разделяется на камеры боками отдельных плодолистиков (септами), то это синкарпный (септированный ценокарпный) гинецей. В зависимости от того, срастаются ли плодолистики краями в центре или нет, можно различать синкарпный и гемисинкарпный (не полностью септированный) гинецей. В таком гинецее выделяют снизувверх синасцидиатную и симпликатную или соответственно гемисимпликатную зоны. Плацентацию по краям отдельных плодолистиков называют центрально-угловой. Если развитие боковых сторон плодолистиков подавлено, то возникает паракарпный гинецей без септ. При этом плаценты располагаются париетально, т. е. там, где соседние плодолистики смыкаются друг с другом краями, или на дне завязи находится свободная центральная плацента, на которой семязачатки сидят на более или менее массивной колонке, не контактирующей со стенками завязи2. В паракарпных гинецеях могут быть и единичные семязачатки, располагающиеся на дне завязи или свисающие с ее крыши.
1 Пестиком называют как весь ценокарпный гинецей, так и каждый плодолистик апокарпного гинецея. - Примеч. ред.
2 Такой тип гинецея иногда называют лизикарпным. — Примеч. ред.
Рис. 11.184. Строение плодолистиков (А — С) и их прогрессирующее срастание (D — F): А — внешний вид сформировавшегося одиночного и свободного плодолистика с вентральной стороны с замкнутым брюшным швом (у основания чашечка; около Зх); В, С — он же в поперечном разрезе, с одним дорсальным и двумя вентральными пучками, двураздельной плацентой и семязачатками на ней (около 10х); D — апокарпный, Е, F — ценокарпный гинецей соответственно со свободными и сросшимися стилодиями (А, В — Colutea arborescens;С, D — Delphinium elatum, E — Linum usitatissimum; F — Nicotians rustica)
В ценокарпных гинецеях некоторые гнезда завязи могут быть стерильными или гинецеи кажутся одноплодолистиковыми (псевдомономерные). Могут, однако, образовываться и добавочные, так называемые «ложные» септы (см. рис. 11.265; 11.277; 11.282).
Функционально важной структурой ценокарпных гинецеев является компитум, под которым понимают общую для всех плодолистиков проводниковую ткань в области столбика и соответственно в симпликатной зоне. Это позволяет, к примеру, в гинецеях с раздельными рыльцами, пыльцевым трубам пыльцевых зерен, попавших только на одно рыльце, прорастать во все плодолистики.
Плодолистики, как и листочки околоцветника, и микроспорофиллы могут участвовать в процессе опыления, к примеру, образуя нектар или как органы вторичного предложения пыльцы. Наконец, структуру гинецея следует понимать и в аспекте образования плодов.
Самые примитивные покрытосеменные чаще всего имеют апокарпный гинецей с варьирующим числом спирально расположенных плодолистиков. Замыкание вентральной щели таких плодолистиков происходит с помощью специального секрета.
Семязачатки
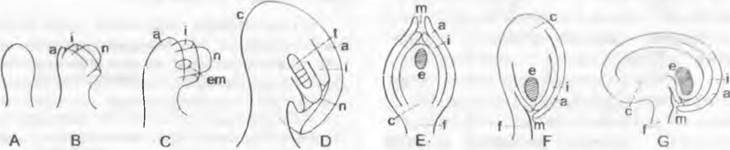
Семязачатки — это заключенные в стерильную оболочку мегаспорангии семенных растений (рис. 11.185). Они состоят из семяножки, чаще всего одной или двух (редко трех) оболочек, или интегументов (семязачатки уни- или битегмальные), и заключенного в интегументы мегаспорангия, или нуцеллуса. Область перехода семяножки в нуцеллус называют халазой, а на противоположном полюсе семязачатка в интегументах остается отверстие — микропиле.
Рис. 11.185. Развитие и типы семязачатков у покрытосеменных: А — D — развитие; Е — атропный; F — анатропный; G — кампилотропный тип; а — наружный, i — внутренний интегументы; с — халаза; е — зародышевый мешок (заштрихован); еm — материнская клетка мегаспор; f — семяножка; m — микропиле; n — нуцеллус; t — тетрада мегаспор
Во время развития семязачатка сначала образуется нуцеллус, а затем его снизу обрастают интегументы. У голосеменных в типе имеется только один интегумент. Если, как у многих покрытосеменных, образуется два интегумента. то внутренний возникает раньше наружного. Унитегмальные семязачатки у покрытосеменных образуются благодаря тому, что один из двух интегументов рано прекращает рост или интеркалярно разрастается общее основание обоих интегументов так, что на верхушке уже не заметна двойная природа или с самого начала имеется только один интегумент. Интегументы могут быть либо очень тонкими и состоять только из двух слоев эпидермы, либо между эпидермами могут быть слои клеток. Самый внутренний слой интегумента, особенно у унитегмальных семязачатков, дифференцирован в подобный тапетуму эндотелий. У некоторых паразитических покрытосеменных (например, у Loranthасеае) интегументов нет.
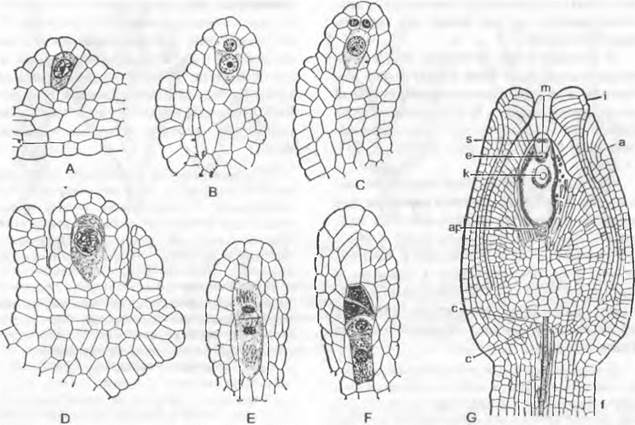
В зависимости от положения материнской клетки мегаспор в нуцеллусе можно выделить следующие формы семязачатков. Если образованная у апикального полюса нуцеллуса материнская клетка мегаспор расположена под эпидермой, семязачаток называют тенуинуцеллятным. В нем мегаспора и развивающийся из нее зародышевый мешок сверху и с боков окружены только эпидермой нуцеллуса. И напротив, если материнская клетка мегаспор отделена от эпидермы нуцеллуса хотя бы одной клеткой (париетальная, или кроющая, клетка), то семязачаток крассинуцеллятный (рис. 11.186). Объем ткани, окружающей апикально и с боков мегаспору и зародышевый мешок в крассинуцеллятных семязачатках, весьма различен. У семенных растений, оплодотворяемых подвижными сперматозоидами (Cycadopsida, Ginkgopsida), на апикальном конце нуцеллуса образуется углубление, называемое пыльцевой камерой (см. рис. 11.180).
Рис. 11.186. Развитие ♀ гаметофитов покрытосеменных (А — F Hydrilla verticillata: Hydrocharitaceae; G — Polygonum divaricatum). В развивающемся нуцеллусе семязачатка дифференцируется одна гиподермальная клетка (А), затем она отделяет кроющую клетку, делящуюся далее (В, С), разрастается в материнскую клетку мегаспор (D) и в ходе мейоза (Е, F) образует 4 мегаспоры, из которых только самая нижняя развивается далее в зародышевый мешок; G — зрелый семязачаток с микропиле (m), наружным (а) и внутренним (i) инте- гументами, халазой (с) и семяножкой (f); зародышевый мешок содержит синергиды (s) и выступающую из-под них яйцеклетку (е), вторичное ядро (к) зародышевого мешка и 3 антиподы (ар) (200х)
Семязачатки можно различать и по ориентации их продольной оси (см. рис. 11.185). Если семяножка и микропиле расположены на одной линии, семязачаток атропный/ орготропный. В антропных семязачатках вследствие изгибания интегументов и нуцеллуса в области халазы на 180° микропиле оказывается вблизи семяножки, но сам нуцеллус остается прямым. В разной степени изогнутый нуцеллус имеется в кампилотропных семязачатках.
У голосеменных встречаются только атроп- ные семязачатки. Изгибание семязачатков многих покрытосеменных связано с достижением большей доступности микропиле для пыльцевой трубки. У покрытосеменных микропиле может быть связано с плацентой обтуратором, развившимся из семяножки, интегументов, нуцеллуса или самой плаценты (см. рис. 11.248).
В области апикального полюса нуцеллуса чаше всего возникает одна материнская клетка мегаспор. В результате мейоза из нее получается чаще всего линейная, реже Т-образная, тетрада мегаспор. Мегаспоры голосеменных имеют клеточную стенку, в которой можно обнаружить спорополленин. Этого не бывает у мегаспор покрытосеменных.
У покрытосеменных исходно, вероятно, были анатропные, крассинуцеллятные семязачатки с двумя интегументами.
Женский гаметофит
В развитии женского гаметофита (зародышевого мешка) принимает участие чаще всего только одна мегаспора (моноспорический зародышевый мешок), реже две
(биспорический зародышевый мешок;некоторые покрытосеменные) или все четыре мегаспоры (тетраспорический зародышевый мешок;Gnetum, Welwitschia, некоторые покрытосеменные). Моноспорический зародышевый мешок развивается чаще всего (в случае линейной тетрады) из самой внутренней мегаспоры, реже — из самой наружной. Женский гаметофит разных групп семенных растений редуцирован в разной степени. В любом случае его развитие начинается с делений ядер, а образование клеточных стенок происходит позже. У голосеменных могут возникать женские гаметофиты из нескольких тысяч клеток (см. рис. 11.180). У всех групп семенных растений, кроме Gnetum, Welwitschia(Gnetopsida) и покрытосеменных, на апикальном полюсе зародышевого мешка образуется разное число архегониев (см. рис. 11.180 и 11.204). У Ginkgo бывает два или три архегония, у хвойных (Coniferopsida) — до 60 и до 100 — у Microcycas. Архегонии состоят из одной, иногда очень крупной, яйцеклетки (диаметром до 6 мм у Cycadopsida; см. рис. 11.180), различного числа клеток шейки (шейковые канальцевые клетки отсутствуют) и иногда из одной брюшной канальцевой клетки или, по крайней мере, из ядра брюшной канальцевой клетки. У семенных растений, оплодотворяемых подвижными сперматозоидами (Ginkgo и Cycadopsida), верхний конец зародышевого мешка отделен от окружающего нуцеллуса расположенной под пыльцевой камерой архегониальной камерой (см. рис. 11.180).
Gnetum имеет тетраспорический зародышевый мешок, целлюларизация которого ограничена только нижним (халазальным) концом. До оплодотворения невозможно различить, какая клетка фукционируст в качестве яйцеклетки. Зародышевый мешок Welwitschia также тетраспорический. Неупорядоченное образование клеточных оболочек приводит здесь к появлению клеток с разным числом ядер, которые затем могут сливаться друг с другом. И у вельвичии распознать яйцеклетку до оплодотворения не удается.
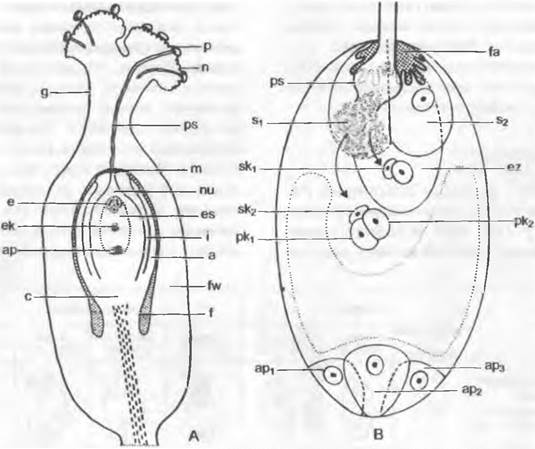
В самом обычном случае у покрытосеменных из одноядерной мегаспоры за три последовательных деления возникают два, четыре и наконец восемь свободных ядер (рис. 11.187; 11.188). По три из них окружаются собственной протоплазмой на верхнем и нижнем концах зародышевого мешка и образуют таким образом клетки, одетые сначала лишь мембраной, а позднее и тонкой клеточной стенкой1. Три верхние клетки называют яйцевым аппаратом. Средняя из них становится более крупной яйцеклеткой, а две других — синергидами.Возможно, что синергиды гомологичны клеткам шейки архегония. Три нижние клетки называют антиподами. Оба оставшихся в цитоплазме зародышевого мешка ядра называют полярными ядрами. До или после проникновения пыльцевой трубки в зародышевый мешок они сливаются во вторичное, диплоидное, ядро зародышевого мешка.
1 Клеточная оболочка яйцеклетки неполная, она прервана на халазальном конце. — Примеч. ред.
Рис. 11.187. Некоторые типы образования зародышевого мешка у покрытосеменных: мейоз диплоидной материнской клетки мегаспор (мегаспорогенез) и развитие гаплоидных мегаспор в зрелый зародышевый мешок (мегагаметогенез)
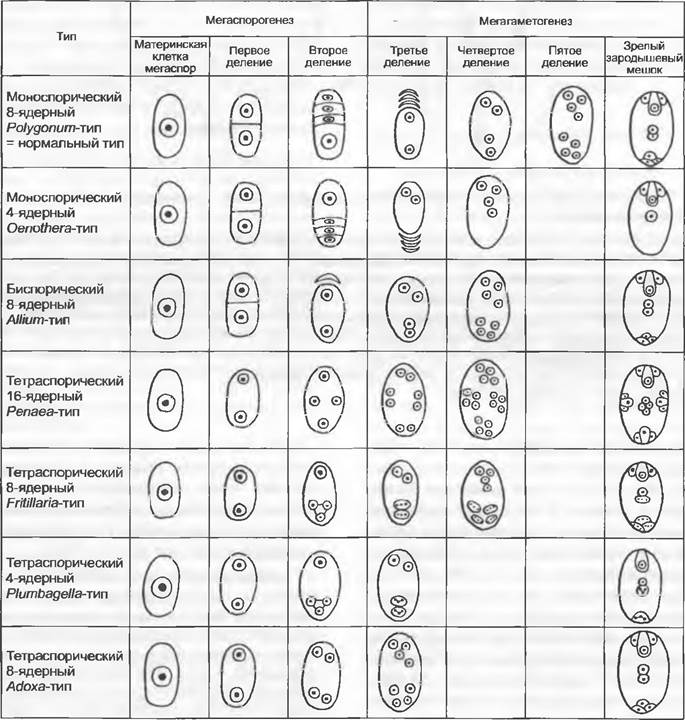
Рис. 11.188. Опыление и оплодотворение у покрытосеменных: А — завязь Fallopia (Polygonum) convolvulus с атропным семязачатком (схематизированный продольный срез, 48х); стенка завязи (fw), столбик (g), рыльце (n) с прорастающими и образовавшим пыльцевую трубку (ps) пыльцевыми зернами (р), семязачаток с семяножкой (f), халазой (с), наружным (а) и внутренним (i) интегументами, микропиле (m) и нуцеллусом (nu), а также зародышевый мешок (es) с яйцевым аппаратом (е), вторичным ядром зародышевого мешка (ек) и антиподами (ар); В — схема зародышевого мешка во время оплодотворения. При проникновении пыльцевой трубки (ps) в область нитчатого аппарата (fa) разрушается одна из двух синергид (s1, s2); ядра спермиев сливаются — одно (sk1) с ядром яйцеклетки (ez), другое (sk2) с двумя сливающимися полярными ядрами (рк1 = рк2); в основании 3 антипода (ар1, ар2, ар3)
Есть много отклонений от этого типа развития (рис. 11.187). Так, в развитии зародышевого мешка могут участвовать не только одна, но и две или все четыре мегаспоры, в результате чего зародышевый мешок бывает би- или тетраспорическим. Другие отклонения затрагивают число ядерных делений, расположение названных групп клеток или особенности слияния ядер. В то время как, например, сформированный зародышевый мешок Реnаеа-типа состоит из 16 клеток, в зародышевом мешке Oenothera-типа их всего 4.
Питание зародышевого мешка обеспечивают прежде всего антиподы, а также гаустории, которые могут образоваться из мегаспор, синергид или антипод.
Нектарники
Нектар — «плата» за посещение цветка опылителями. В то время как у опыляемых животными голосеменных эту функцию может выполнять опылительная капля, выделяемая через микропиле, у покрытосеменных имеются выделяющие нектар железки — нектарники. Они могут быть множественными и не связанными друг с другом (см. рис. 11.264) или чаще всего собраны в кольцевидный диск — разрастание оси цветка между андроцеем и гинецеем (см. рис. 11.267). Чашелистики могут образовывать нектар в определенных местах на внутренней или внешней стороне, а лепестки — на внутренней стороне. Тычинки, к примеру, могут нести нектарники на нитях или преобразуются в стаминодии и служат только для производства нектара (см. рис. 11.236). В гинецее функцию нектара может выполнять секрет рылец, а в ценокарпном гинецее бывают септальные нектарники — полости между краями плодолистиков, сообщающиеся протоками с внешней средой (см. рис. 11.228). Возможно также образование нектара и на поверхности завязи.
Нектарники за пределами цветков называют экстрафлоральными, они могут служить, в частности, для «расплаты» с охраняющими растение муравьями.
Расположение элементов цветка
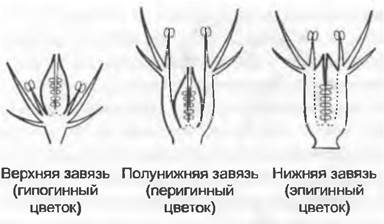
Возможные варианты расположения элементов цветка — спиральное или мутовчатое (круговое), в более редких случаях беспорядочное — были рассмотрены выше. Устройство цветков у покрытосеменных разнообразно. В так называемых полных цветках почти всегда следуют друг за другом околоцветник, андроцей и гинецей. В зависимости от числа и положения элементов цветка его ось (цветоложe) бывает более или менее вытянутой, или сильно укороченной. Она может даже превратиться в диск или даже в бокал или трубку (гипаптий). Благодаря образованию гипантия основания элементов околоцветника и тычинок отдалены от основания гинецея. Существует также возможность удлинения междоузлий внутри цветка. Если вытянуто междоузлие между андроцеем и гинецеем (см. рис. 11.265), то имеется гинофор. При вытягивании междоузлия между околоцветником и андроцеем/гинецеем получается андрогинофор. В зависимости от локализации ростовых процессов в гинецее его относительное положение в цветке бывает разным (рис. 11.189). Если в первую очередь растут свободные спинные стороны (и бока) плодолистиков, то получается верхняя завязь. Околоцветник и тычинки при этом расположены у основания гинецея, поэтому такие цветки называют также гипогинными. Рост цветоложа ниже периферического участка гинецея приводит к образованию нижней завязи. Из-за прикрепления околоцветника и тычинок выше завязи такие цветки называют также эпигинными. Промежуточный тип — полунижняя завязь. Если завязь свободно располагается в цветочном бокале, то она перигинная. В циклических цветках члены одного круга чаще всего располагаются в промежутках между членами предыдущей мутовки, так что члены соседних кругов чередуются. Как отклонение от правила чередования кругов органы следующих друг за другом мутовок располагаются на тех же радиусах («противолежат» друг другу). В пятикруговых цветках некоторых покрытосеменных (околоцветник с чашечкой и венчиком, два круга тычинок, гинецей) при соблюдении правила чередования кругов тычинки внешнего круга расположены на радиусах, чередующихся с лепестками, и противолежат чашелистиками (эписепальные), а тычинки внутреннего круга — на радиусах, чередующихся с наружными тычинками (эпипетальные).
Рис. 11.189. Положение завязи в цветках покрытосеменных
Чередование кругов может нарушаться, если какой-либо круг выпадает. Если это случается, к примеру, с наружным из двух кругов тычинок, то тычинки единственного оставшегося круга расположены против лепестков (эпипетально). Из-за нарушения чередования кругов в гинецее в цветке с двухкруговым андроцеем тычинки внутреннего круга могут быть смещены наружу разрастающимися эпипетальными плодолистиками. В зрелом цветке при этом кажущийся внешний круг тычинок расположен против лепестков (эпипетально), а кажущийся внутренний — против чашелистиков (эписепально). Это явление называют обдиплостемонией.
В цветках покрытосеменных наиболее обычно мутовчатое (циклическое) расположение элементов. Число мутовок (кругов) в цветке бывает разным. Особенно часто встречаются пятикруговые цветки с пятью мутовками (два круга околоцветника, к примеру, чашечка и венчик, два круга тычинок и чаще всего не подчиняющийся правилу чередования кругов круг плодолистиков) и — при выпадении одного круга тычинок — четырехкруговые цветки. Бывают, однако, и двух- или однокруговые цветки всего с одним кругом элементов.
Строение и расположение элементов зрелого цветка определяют его симметрию (рис. 11.190). Цветки со спиральным расположением элементов первично асимметричны. Среди циклических цветков различают радиально-симметричные (актиноморфные) с более чем двумя плоскостями симметрии, дисимметричные — с двумя плоскостями симметрии и зигоморфные цветки — только с одной плоскостью симметрии. Циклические цветки могут быть и вторично асимметричными.
Рис. 11.190. Типы симметрии и диаграммы цветков: А — Sedum sexangulare, актиноморфный; В — Dicentra spectabilis, дисимметричный; С — Lamium album, зигоморфный
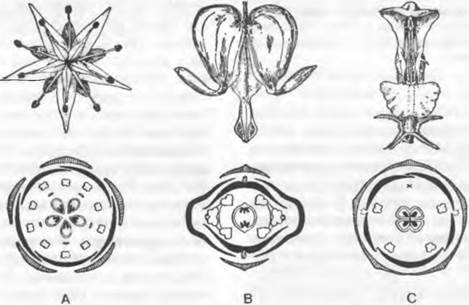
При более тщательном рассмотрении симметрии выделяют медианную плоскость, которую можно провести через стебель материнского побега, цветоложе и кроющий лист цветка. Перпендикулярна к ней трансверсальная плоскость, а остальные плоскости косые. Соответственно можно различать, к примеру, медианно-, трансверсально- или косозигоморфные цветки.
Строение цветков можно лучше всего представить на диаграмме цветка (рис. 11.190). Эмпирические диаграммы представляют непосредственно наблюдаемое строение цветка, а теоретические содержат также интерпретации и указывают, к примеру, что какие-то из ожидаемых органов не развиты. Формулы цветков содержат информацию: о типе симметрии цветка
![]()

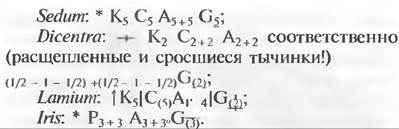
Соцветия
Семенные растения имеют чаще всего несколько или много цветков, которые нередко собраны в соцветия (см. 4.2.5.3). Однозначного определения соцветия нет. Здесь мы понимаем его как репродуктивную систему ветвей, которая после завершения цветения и плодоношения далее не растет и обычно сбрасывается.
Листья в соцветиях либо отсутствуют, либо представлены более или менее невзрачными верховыми листьями (брактеями и предлистьями1). Отдельные мелкие цветки становятся более заметными при тесном расположении (например, Iberis), при наличии увеличенных, часто стерильных краевых цветков (например, Hydrangea, Viburnum opulus) или ярко окрашенных верховых листьев (например, Astrantia, Comus suecica). Так, благодаря разделению функций между отдельными цветками и при участии добавочных осевых и листовых образований в конце концов возникает функциональное биологическое единство, аналогичное отдельному цветку и называемое псевдантием. В качестве примеров можно назвать циатии Euphorbia, корзинки Dipsacaceae и Asteraceae или окруженный окрашенным покрывалом початок Arum. Впрочем, бывает и наоборот: такое функциональное единство представлено только частью цветка — мерантием (например, Iris).
1 Предлистья — листья низовой формации. — Примеч. ред.
Опыление
Опыление (англ. pollination) — процесс переноса пыльцевых зерен на микропиле семязачатков голосеменных или на рыльца плодолистиков покрытосеменных растений.
Различают самоопыление (автогамия) — опыление в пределах одного индивида, и перекрестное опыление (аллогамия) — опыление между двумя индивидами. Аллогамия — единственный тип опыления, когда мужские и женские репродуктивные структуры находятся на разных индивидах (двудомность), как у Cycadopsida, Ginkgo, многих Comferopsida и Gnetopsida и малой части покрытосеменных. Если однополые цветки находятся на одном и том же растении (однодомность) или хотя бы некоторые цветки обоеполые, то в принципе возможна и автогамия. Важнейшие механизмы, которые во многих ситуациях препятствуют вредной в эволюционно-биологической перспективе автогамии и самооплодотворению, — это самонесовместимость, гетероморфия, дихогамия и геркогамия (см. 10.1.3.1 и 10.1.3.2). Крайнюю степень автогамии представляет клейстогамия — самоопыление и самооплодотворение в нераскрывающемся бутоне. Например, у некоторых видов Viola и кислицы (Oxala acetosella) один и тот же индивид образует как клейстогамные, так и распускающиеся (хасмогамные) цветки. У Lamium amplexicaule кпейстогамные цветки развиваются главным образом в начале и в конце вегетационного периода.
Важнейшими переносчиками пыльцы являются ветер, вода и разные животные.
Ветроопыление (анемофилия) требует образования большого количества пыльцы, быстрого и равномерного высвобождения ее в воздух, максимально долгого пребывания в воздухе1, а также открытых и крупных рылец, чтобы легко улавливать пыльцу. Ветроопыляемые цветки чаще всего невзрачные, без запаха и нектара. Обычно они однополые; и мужских, или тычиночных, цветков значительно больше, чем женских; пыльцевые зерна более или менее гладкие, а пыльца порошковидная вследствие отсутствия или раннего высыхания пыльцевого клея.
1 Пыльца ветроопыляемых растений теряет способность оплодотворять после часового пребывания в воздухе, поэтому пыльцевые зерна анемофилов относительно тяжелые, что позволяет сократить время пассивного полета. — Примеч. ред.
В связи с этими потребностями у ветроопыляемых видов возник комплекс (синдром) типичных признаков. Пыльцевые зерна у них не слипаются и способны долго держаться в воздухе либо из-за своей малой величины, либо благодаря особым воздухоносным мешкам (некоторые хвойные). Массовое образование пыльцы обусловлено увеличением пыльников и/или числа мужских цветков или тычинок. У Cotylus, к примеру, один семязачаток приходится на 2,5 млн пыльцевых зерен. Высыпание пыльцы из пыльников облегчается подвижностью тычиночных нитей (например, Роасеае; см. рис. 11.232), цветоножек (например, конопля: Cannabis; см. рис. 11 257) или оси всего соцветия (например, мужские сережки лещины: Corylus, ольхи: Alnus, дуба: Quercus; см. рис. 11.259и 11.260). При этом пыльца часто сначала оседает на элементах цветка или соцветия, с которых потом ее сдувает ветер. В мужских цветках Urtica (см. рис. 8.34) и Pilea происходит взрывное высвобождение пыльцы при резком распрямлении тычиночных нитей. Длительность нахождения пыльцы в воздухе и определяемое этим расстояние ее распространения в значительной мере зависят от высоты расположения мужских цветков над землей, а также от структуры растительности. Пыльца ветроопыляемых растений вызывает у многих людей аллергию (например, сенную лихорадку). Столбики и рыльца женских цветков ветроопыляемых покрытосеменных чаще всего сильно увеличены, что повышает вероятность опыления. Число семязачатков в завязях обычно сильно редуцировано, а цветки расположены открыто. Околоцветник, только мешающий опылению, редуцирован или совсем отсутствует. Наконец, опыление облегчается и благодаря раннему цветению, в умеренном климате часто даже предшествующему распусканию листвы (например, дуб, ольха, вяз, тополь, ясень, лещина).
Географическое распространение ветроопыляемых покрытосеменных зависит от условий местообитания. Они часто растут массовыми зарослями в открытых ветру саваннах, степях и арктическо-альпийских биотопах или в древесном ярусе субтропических и бореальных лесов. Во влажных тропических лесах ветроопыляемые растения редки.
В некоторых группах растений, где встречаются как ветроопыляемые, так и зоофильные виды (например. Thalictrum, Sanguisorba; рис. 11.191, Acer, Fraxinus, Artemisia), иногда легко понять возникновение анемофилии. Это относится, в частности, к Tilia и Calluna, а у Castanea пыльца сначала покрыта пыльцевым клеем, и ее собирают насекомые, но когда она высыхает, то разносится ветром.
Рис. 11.191. Переход от энтомофилии (А) к вторичной анемофилии (В, С) у представителей рода Sanguisorba (Rosaceae): А — обоеполый цветок S. officinalis с 4 тычинками, сосочковым рыльцем и нектарником; В, С — однополые цветки S. minor без нектара, ♀ с перистыми рыльцами, ♂ с многочисленными тычинками (около 6х)
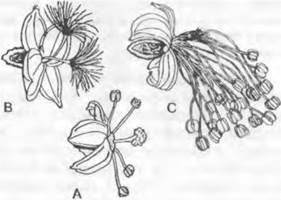
Опыление водой (гидрофилия), т. е. перенос пыльцы или мужских цветков водой, встречается лишь у немногих покрытосеменных. Здесь можно различать транспорт над, по или под поверхностью воды.
В прямостоячих цветках дождевая вода служит самоопылению, реже — способствует перекрестному опылению. Гидрофилия встречается отнюдь не у всех водных растений. Чаще всего их цветки поднимаются над водной поверхностью и опыляются ветром или животными (например, Potamogeton-, см. рис. 11.223). У Vallisneria (см. рис. 11.223) и Elodeaоторвавшиеся мужские цветки, а у Callitriche плавучая пыльца достигают рылец, находящихся в это время на поверхности воды. Под водой опыляются, к примеру, CeratophyUum, Najas и Zostera (см. рис. 11.223). У последнего рода пыльцевые зе- ча нитевидные, длиной до 0,5 мм, без экзины. У многих опыляемых водой видов наблюдается параллелизм в строении цветков с ветроопыляемыми формами.
Необыкновенное многообразие цветков покрытосеменных можно понять только в контексте приспособления к опылению животными (зоофилия). Зоофилия — важная причина видового разнообразия покрытосеменных, так как все возрастающая специализация цветков к опылению немногими или единственным опылителем ведет к репродуктивной изоляции — важному фактору видообразования.
Зоофилия может осуществляться только в том случае, если опылитель замечает цветки и посещает их регулярно и достаточно долго и, если цветки устроены так, что опылитель соприкасается с пыльцой и рыльцем и при этом переносит пыльцу. Чтобы привлечь внимание потенциального опылителя, цветки обладают средствами дальней аттрактации (англ. advertisment), а регулярность посещения обычно обеспечивают приманки (англ. reward).
Средства дальней аттрактации имеют оптическую и химическую природу: окраска и запах. Окраску цветков в первую очередь обусловливают пигменты, содержащиеся в вакуолях (антоцианы: синий, фиолетовый, красный; антоксантины: желтоватый, белый, ультрафиолетовый; беталаины: красно-фиолетовый, желтый; халконы и ауроны: желтый, ультрафиолетовый) или в пластидах (каротиноиды: оранжевый каротин и желтый ксантофилл). С краски могут быть модифицированы благодаря тому или иному расположению в органе клеток с пигментами, наложению слоев с различной окраской, частоте и величине межклетников, а также поверхностной структуре эпидермы. Так, гладкие эпидермальные клетки делают окраску блестящей, а клетки с сосочками — матовой. Белая окраска цветков может возникать и за счет полного отражения света, прежде всего межклетниками.
Окраска цветка может изменяться в процессе его старения. Например, после опыления окраска пятен в цветках Aesculus меняется с желтой на красную. Для потенциальных насекомых-опылителей красные цветки менее заметны и привлекательны, так как воспринимаются ими как уже «бесполезные».
Понимание оптического воздействия цветков требует знания физиологии зрения опылителей, которая изучена пока у немногих животных, таких, как медоносная пчела, шмели, бражники, жужжало среди мух и некоторые колибри. Оказалось, что пчелы и шмели не видят чисто красный цвет, но зато воспринимают невидимый для человека ультрафиолет с длиной волны 310 — 400 нм, а среди остальных окрасок цветков реагируют только на желтые (520 — 650 нм), синефиолетовые, включая пурпурные оттенки (400 — 480 нм), и белый цвет, который воспринимают как сине-зеленый. Зрительное восприятие птиц подобно человеческому: для них наиболее заметен красный цвет. Опыты по дрессировке насекомых-опылителей показали, что оптическая привлекательность может зависеть также и от совместного действия таких факторов, как степень сочности, яркости цветка, контраст яркости и цвета, а также форма частей цветка. Подобным образом удалось доказать значение рисунка и цветных пятен на лепестках цветков, показывающих дорогу к нектару, как, например, оранжево-желтый зев в лимонно-желтых цветках Linaria vulgaris. Распространено мнение, что цветовые указатели имитируют пыльники или пыльцу. Часто указатели заметны только для насекомых, воспринимающих ультрафиолетовый свет (например, кажущиеся нам равномерно желтыми листочки околоцветника Caltha palustris). Оптическим аттрактантом могут быть и движения всего цветка или его частей.
Запах цветкам придают самые разные вещества. Чаще всего запахи, приятные для человека, обусловлены терпенами и бензеноидами, а также простыми спиртами, кетонами, эфирами органических кислот, фе- нилпропанами и др. Некоторые из этих веществ, например, у Ophrys, могут имитировать феромоны самок насекомых, побуждая самцов совершать попытки копуляции. Неприятные запахи многих цветков или соцветий, опыляемых мясными или навозными мухами, обусловлены аминами, аммиаком или индолами.
Запах в принципе может исходить от любых частей цветка и локализован значительно менее упорядоченно, чем окраска. Соответственно и движение опылителей на запах обычно менее регулярное и отчетливое, чем при зрительной аттрактации. Многие цветки обладают сходными со зрительными также и запаховыми указателями, которые иногда расположены в тех же местах. Примеры: корона Narcissus (см. рис. 11.228) и чешуи на основании отгиба лепестка у некоторых видов Silene (см. рис. 11.239).
У пчел и других перепончатокрылых (Нуmenoptera) повторное успешное посещение цветков одного вида вызывает ограниченное во времени стремление посещать цветки этого вида и далее. Это основано на стимулирующем действии видоспецифического запаха частей цветка, принесенного в улей нектара и пыльцы, а также высокоразвитой памяти этих животных и их способности обмениваться информацией.
Опылители посещают цветки ради разных первичных аттрактантов. Огромное значение для опылителей имеет пищевая ценность пыльцы и нектара. В пыльцевых цветкахобразуется в изобилии пыльца, богатая белками, жиром, углеводами и витаминами. Такие цветки, доступные и примитивным насекомым с грызущим ротовым аппаратом, имеют многие Magnoliidae (например, Winteraceae, Victoria) и Ranunculales (Anemone), а также таксоны с вторично полимерным андроцеем (например, Papaver, Rosa). Рад пыльцевых цветков имеет немного тычинок со снабженными порами пыльниками (например, Solanum dulca marа), из которых пыльца высвобождается при вибрации посещающих цветки насекомых (англ. buzz polination).
Нектар нектарных цветков — это в основном водный раствор сахаров (сахарозы, фруктозы, глюкозы), но обычно содержит также аминокислоты. Предложение нектара менее обременительно для растения, чем предложение богатой фосфором и азотом пыльцы. Нектар могут образовывать разные части цветка, он в разной степени доступен животным-опылителям. Он может находиться открыто, например, на дне цветка у многих Rosaceae, но может быть скрыт глубоко в трубке венчика или в шпорцах (например, Viola, Linaria, Cotydalis), откуда его могут достать лишь животные с длинным ротовым аппаратом.
Масличные цветки некоторых покрытосеменных (например, Lysimachia-, Calceolaria; многие Malpighiaceae) образуют в особых железках масло, служащее опылителям пищей и иногда как строительный материал. Смолу для строительства гнезд образуют цветки Dalechampia (Euphorbiaccac) и Clusia (Clusiaceae) — смоляные цветки. В душистых цветках(например, Stanhopea: Orchidaceae; Gloxinia; Gesneriaceae) самцы пчел эуглоссин собирают пахучие вещества, имитирующие феромоны опылителей и, как полагают, используют их при спаривании.
Цветки могут по-разному использовать стремление животных к размножению. Так, соцветия Ficus и цветки Zamia (Cycadopsida)1, Yucca или Siparuna служат насекомым местом откладки яиц.
1 У всех саговниковых есть стробилы, а не цветки. — Примеч. ред.
У Ficus carica в течение всего года появляются кувшиновидные соцветия, в которых в разных сочетаниях находятся цветки трех типов (рис. 11.192). Наряду с мужскими имеются и женские цветки с длинным столбиком и коротким столбиком. Цветки с длинными столбиками образуют семена, тогда как цветки с короткими столбиками служат местом для откладывания яиц и развития личинок галловой осы Blastophaga psenes. Последовательность распускания цветков в течение года, с одной стороны, и в каждом соцветии — с другой, обеспечивает, во-первых, опыление и оплодотворение растения и, во-вторых, размножение насекомого.
Рис. 11.192. Зоофилия у разных покрытосеменных: А — шмель как опылитель; В — цветок Salvia pratensis (фиолетово-синий, несколько увеличено); С, D — рычажный механизм у Salvia pratensis; каждая из двух тычинок имеет вытянутый в длинный рычаг связник (с), подвижно соединенный с короткой тычиночной нитью (f); только на переднем, удлиненном, конце рычага имеется фертильная тека; вторая, стерильная (s), тека образует более короткое плечо рычага, которое объединено в пластинку с соответствующей частью второй тычинки; Е, F — птица-медосос Arachnothera longirostris как опылитель Sanchezia nobilis (Acanthaceae, цветки желтые, брактеи пурпурные, около Зх); G — разрезанное продольно соцветие Аrum maculatum на женской стадии развития со светло-зеленым покрывалом и невзрачными мужскими, женскими и стерильными цветками (h), загораживающими вход (2/Зх); Н — соцветие инжира (Ficus carica) в продольном разрезе (немного увеличено) с J — мужскими и К — длинностолбиковыми женскими фертильными цветками, а также L — с короткостолбиковыми женскими галловыми цветками (увеличено)
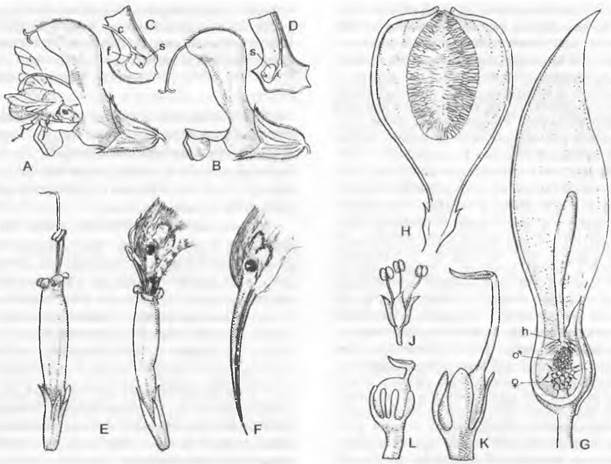
В роде Ophrys (Orchidaceae) губа цветка имитирует формой, опушением и даже запахом самку определенных видов пчел и роющих ос, что побуждает самцов совершать попытки спаривания, которые могут привести к опылению. Недавно удалось показать, что запах цветков одного из видов имитирует феромоны соответствующих насекомых. Так как Ophyrysничем не вознаграждает опылителя, он — цветок-обманка. Конструкция цветка должна гарантировать, что опылитель в соответствии со строением своего тела будет контактировать с пыльцой и рыльцем. При этом пыльца может быть помещена на опылителя с помощью
специальных рычагов, зажимов, путем приклеивания или «выстреливания» пыльцой.
Протандрические цветки Salvia pratensis известны благодаря своему эффективному рычажному механизму, описанному С. К. Sprengel еще в 1793 г (рис. 11.192). У них всего две тычинки, каждая из которых несет по вытянутому в длинный рычаг связнику, примыкающему к верхней губе цветка и подвижно соединенному с короткой тычиночной нитью. Только на переднем2, более длинном плече рычага находится фертильная тека. Другая, стерильная, тека образует заднее1, более короткое плечо, объединенное в пластинку с соответствующей частью второй тычинки. Эта пластинка закрывает доступ к нектару в основании трубки венчика. Когда шмель надавливает на пластинку, более длинный конец рычага изгибается и прижимает теку с пыльцой к спине Животного. В более старых цветках в том же положении оказывается уже рыльце, на которое насекомое и переносит пыльцу.
1 На самом деле это заднее плечо. — Примеч. ред.
2 Переднее. — Примеч. ред.
Пример особенно сложного механизма опыления представляет функционирующее как ловушка соцветие видов рода Arum (рис. 11.192). Однополые цветки собраны на нижнем конце толстого обоеполого протогинного початка, окруженного светлым покрывалом, которое внизу расширено, выше сужается, а наверху широко раскрыто. В самом низу початка расположены женские цветки, наверху— мужские, а между ними и над мужскими находятся стерильные цветки, преобразованные в толстые щетинки, загораживающие вход в полость покрывала. Вне полости ось початка булавовидно утолщена и у особенно хорошо изученного A. nigrum в первое после распускания утро издает резкий навозный запах. Выделение пахучего вещества стимулируется повышением температуры в початке, а его выход наружу осуществляется через отверстия системы межклетников («дырявая эпидерма»). Этот запах привлекает разных любителей навоза (мух и жуков), часть которых уже несет на себе пыльцу из других соцветий. Садясь на гладкую, покрытую маслом эпидерму внутренней поверхности покрывала или оси початка и пытаясь удержаться на ней, насекомые соскальзывают и падают в полость покрывала. Выбраться тем же путем невозможно, так как «горлышко» полости перегорожено торчащими внутрь щетинками стерильных цветков, а ее стенки, как и верхняя часть, гладкие и скользкие. Копошась внизу, насекомые опыляют принесенной пыльцой женские цветки. Ночью из вышерасположенных мужских цветков высыпается пыльца и прилипает к насекомым. Одновременно прекращается выделение запаха. Наконец, на следующий день после увядания загораживающих выход цветков нагруженное пыльцой насекомое может выбраться из ловушки и отправиться на поиски новых соцветий. «Ловушками» являются также цветки разных видов Aristolochia. У представителей рода Ceropegia (Apocinaceae) в цветках-ловушках есть еще и зажимы.
Опылитель может получить пыльцу не только из пыльников. При вторичном предложении пыльцы другие части цветка принимают пыльцу из пыльников и нагружают ее на опылителя.Пример — протерандрические цветки Asteraceae, у которых пыльца высыпается в трубку, образованную склеившимися пыльниками, а уже из нее выталкивается удлиняющимся столбиком. На опылителя пыльца попадает с верхушки столбика.
Для удержания пыльцы на поверхности тела опылителей служат прежде всего пыльцевой клей и висциновые нити, но наверняка отчасти и снабженная шипиками или другими подобными структурами поверхность экзины (см. бокс 4.1, рис. F).
Можно выделить несколько типов цветков по их особенностям, тесно связанным со взаимодействием с опылителями, которые могут реализоваться на уровне части цветка, отдельного цветка или соцветия. В плоских дисковидных и чашевидных цветках (например, Anemone, отдельный цветок; Matricaria: соцветие; Tilia, Еuporblа: соцветие) доступ к центру цветка и имеющемуся там вознаграждению практически беспрепятственный. Он ограничен в бокальчатых и колокольчатых цветках (например, Hyoscyamus, Crocus) и еще более — в трубчатых цветках и цветках с длинноноготковыми лепестками (например, Silene, Nicotiana). В шпорцевых цветкахнектар скапливается в шпорце (например, Linaria, Viola), а в трубчато-колесовидных цветках имеется несколько проходов к вместилищам нектара (например, Gentiana acaulis). Зигоморфны мотыльковые (например, Corydalis, Pisum, Polygala), одно- и двугубые цветки (например, Aconitum, Viola: отдельные цветки, Mimetesr. соцветие). В щетковидных цветках обычно многочисленные тычинки далеко выступают из цветка (Syzygium, Acacia и Salix: соцветия) и, наконец, цветки-ловушки на время ловят насекомых (например, Asclepias: ловушки-зажимы1, Arum: скользкие ловушки).
1 В цветках Аsclepias поллинии прикрепляются к телу опылителя специальными зажимами. Мелкие насекомые, не способные опылять виды этого рода, слишком слабы, чтобы извлечь прикрепившийся к ним поллиний. Они остаются в цветке и погибают. Опылители же без проблем покидают цветок, унося поллинии. Ничего общего с цветками-ловушками это не имеет, никакого временного удержания опылителя в цветке здесь нет. — Примеч. ред.
Эту классификацию можно сделать еще более дробной, а некоторые цветки проявляют черты разных типов. Поэтому не всегда легко отнести цветок к какому-то определенному типу.
Строение тела и ротового аппарата, особенности поведения и пищевые пристрастия многих животных-опылителей создают специфические требования к устройству посещаемых ими цветков, обусловив их селективное преобразование. Таким образом, можно различать характерные опылительные синдромы цветков.
Правильность этого воззрения подтверждают экспериментальное исследование высокой способности к различению у большинства посетителей цветков и тот факт, что функционально очень сходные синдромы возникали на совершенно разной структурной основе: одиночного цветка, мерантия или соцветия. По особенностям цветка иногда предсказывали неизвестного еше опылителя, которого обнаруживали впоследствии. При анализе селективных влияний опылителей цветка на его строение следует принимать во внимание, что большинство цветков опыляется значительным числом разных видов животных и, таким образом, полифильны. Возрастающая специализация привела, однако, и к возникновению олиго- и монофильных цветков, опыляемых немногими или всего одним опылителем. Учитывая обратное селективное влияние цветков на опылителей, можно говорить о коэволюции между растениями и животными.
Среди (опыляемых насекомыми) энтомофильных растений опыляемые жуками (кантарофилия) чаще всего имеют легкодоступные крупные дисковидные или чашевидные цветки зеленой или светлой окраски без нектара, обычно сильно пахну щие и продуцирующие много пыльцы. Это соответствует поведению жуков — сравнительно неловких насекомых с грызущим ротовым аппаратом, нередко повреждающих цветки. Жуки опыляют цветки многих Magnoliidae, а также дисковидные цветки эволюционно продвинутых таксонов (например, Comus, Viburnum-, соцветия — псевдантии). Гетерогенны цветки, опыляемые мухами (миофилия). К ним относятся, с одной стороны, мелкие, более или менее не пахучие дисковидные цветки с открыто расположенным нектаром (например, Apiaceae, Ruta), а с другой стороны, цветки, опыляемые навозными мухами (сапромиофилия), подражающие зелено-пурпурной пятнистой окраской и запахом навоза источникам пищи своих опылителей. Последние чаще всего являются цветками-обманками и/или цветками-ловушкам и и попросту «эксплуатируют» своих опылителей (например, Aristolochia; цветок; Arum: соцветие-псевдантий). Особенно разнообразны и обычны опыляемые пчелами цветки (мелиттофилия). Для них характерны зигоморфное строение мотыльковых и двугубых цветков с посадочной площадкой для опылителя, обычно желтая, фиолетовая или синяя окраска, легкий запах и чаще всего глубоко скрытый нектар (например, Salvia). Особые приспособления для опыления имеются у крупных тропических древесных пчел рода Xylocopaи у неотропических эуглоссиновых пчел. Цветки, опыляемые дневными бабочками (психофилия), выделяются прямым положением, узко-трубчатым околоцветником, обычно карминово-красной окраской и глубоко спрятанным нектаром (например, Dianthus carthusianorum, Nicotiana tabacum). В противоположность открытым днем психофильным цветкам опыляемые ночными бабочкам и молями цветки (сфингофилия и фаленофилия) раскрываются вечером. Это расположенные горизонтально или свисающие узко-трубчатые цветки с беловатой окраской, сильным ароматом и глубоко спрятанным нектаром (например, Oenothera, Silene, Lonicera periclymenum). Замечательна орхидея Angraecumsesquipedale с Мадагаскара со шпорцем длиной до 43 см. Предположили, что ее опыляет какой-то бражник, и в дальнейшем его действительно обнаружили. Отсюда происходит название этой бабочки — Xanthopan morgani praedicta.
Названные синдромы цветков— лишь отдельные примеры. Энтомофильные цветки столь же гетерогенны, как и названные группы их опылителей (например, мухи и пчелы). Среди насекомых опылителями являются также Orthoptera, Hemiptera, Thysanoptera и представители других таксонов.
Опыляемые птицами цветки (орнитофилия) отчетливо отличаются от цветков, опыляемых насекомыми. Места для посадки в самом цветке чаще всего нет, и значи
тельно более тяжелые птицы должны, посещая цветки, зависать в воздухе (колибри) или доставать до них с прочной опоры. Обычно это крупные цветки бокальчатого, трубчатого или щетковидного типа, преимущественно огненно-красные или с контрастным рисунком красных, а также синих, желтых и зеленых тонов («попугайные расцветки»). Запах у них отсутствует, и соответственно у птиц-опылителей плохо развито обоняние, зато обильно выделяется жидкий, как правило глубоко спрятанный, нектар, который птицы достают с помощью языка, трубковидного или снабженного щеточкой папилл. Пыльца переносится иногда на клюве, но чаще на других частях головы и реже на ногах (см. рис. 11.192). Орнитофильные цветки есть почти во всех тропических семействах с зоофильными цветками (например, Erythrina, Fuchsia, Hibiscus tiliaceus, Tropaeolum majus, Salvia splendens, Aloe,в том числе и виды, культивируемые в Центральной Европе). Важнейшие птицы-опылители — колибри в Неотропической области, нектарницы и медососы в Палеогропической. Тем не менее в обеих этих областях встречаются птицы- опылители и из многих других семейств.
Цветки, опыляемые рукокрылыми (хироптерофилия), ограничены тропиками. Их опылители — в первую очередь длинноязычные крыланы в Старом Свете или летучие собаки и длинноязычные вампиры — в Новом. Для хироптерофильных видов характерны открыто расположенные, прочные, чаще всего бокальчатые с широким зевом или щетковидные цветки, раскрывающиеся ночью, часто светло- или темноокрашенные, с сильным фруктовым или винным запахом и очень большим количеством нектара (и пыльцы) (например, Camegiea, Adansonia, Cobaea, виды родов Musa и Agave).
Некоторые другие мелкие млекопитающие, к примеру грызуны, но прежде всего сумчатые, также могут опылять растения.
У современных голосеменных наряду с анемофилией известно опыление жуками (у Сусаdopsida), а также молями (например. Gnetum). Опылителями первых покрытосеменных были, вероятно, преимущественно жуки и мухи, но, возможно, также осы и некоторые моли. Такие опылители, как чешуекрылые (дневные и ночные
бабочки), птицы, рукокрылые и высокоразвитые пчелы, появились лишь в позднем мелу или даже в третичном периоде и эволюционировали затем совместно с опыляемыми ими покрытосеменными растениями.
Оплодотворение
После того как пыльцевые зерна в результате опыления попали в пыльцевую камеру на верхушке нуцеллуса (Cycado- psida, Ginkgo), в микропиле (Coniferopsida, Gnetopsida) или на рыльце пестика (Аngiospermae) начинается прорастание пыльцы, приводящее к оплодотворению. Опыление и оплодотворение могут быть разделены во времени, особенно у некоторых групп голосеменных, несколькими месяцами. Пыльцевое зерно прорастает пыльцевой трубкой из апертуры в оболочке.
У семенных растений с подвижными сперматозоидами (Cycadopsida, Ginkgo) функция пыльцевой трубки состоит в основном в том, чтобы закрепить мужской гаметофит на нуцеллусе и в качестве гаустории всасывать из него питательные вещества (см. рис. 11.180). Впрочем, пыльцевая трубка растет также и по направлению к архегониям при разрушении нуцеллуса между пыльцевой и архегониальной камерами. В заполненной жидкостью архегониальной камере над архегониями сперматозоиды выходят из пыльцевой трубки и плывут к архегониям. После их проникновения в архегонии оплодотворение завершается слиянием гамет (плазмогамия) и ядер (кариогамия). Оплодотворение подвижными сперматозоидами у саговников и гинкго называют зооидиогамией.
При оплодотворении спермиями, т.е. неподвижными мужскими гаметами (сифоногамия: Coniferopsida, Gnetopsida, Angiospermae), пыльцевая трубка должна доставить спермин непосредственно к яйцеклеткам (см. рис. 11.204; 11.188). При этом она должна прорасти через нуцеллус, по пути локально растворяя его клетки. У Welwitschia (Gnetopsida) обнаружено совершенно необычное явление, когда не только пыльцевая трубка растет к женскому гаметофиту, но и от женского гаметофита навстречу ей растут трубковидные структуры.
У покрытосеменных пыльцевая трубка с поверхности рыльца должна прорасти сквозь ткани плодолистика, например, столбика, чтобы достичь семязачатка. Рост идет по проводниковому каналу. При этом пыльцевая трубка может расти либо в секрете на поверхности этого канала, в его эпидерме, либо в более глубоко расположенных слоях клеток. В ценокарпных гинецеях существует возможность того, что объединенная в симпликатной области всех плодолистиков часть проводниковых каналов, компитум, распределит трубки пыльцевых зерен, проросших только на одном рыльце, по всем плодолистикам. Путь пыльцевой трубки от плаценты к микропиле облегчается благодаря анат- ропному или кампилотропному строению семязачатков либо разнообразным выростам тканей между плацентой и микропиле (обтуратору).
В пыльцевой трубке покрытосеменных протоплазма с двумя спермиями и ядром пыльцевой трубки всегда находится в концевом участке, перетекая из более старых частей и часто отделяясь от них каллозной пробкой. Скорость роста пыльцевой трубки составляет 1 — 3 мм/ч, но может быть и значительно меньше, так что оплодотворение и у покрытосеменных может произойти спустя недели или месяцы после опыления (например, у орхидных). Однако задержка может быть связана также с запаздыванием развития самого семязачатка.
У хвойных и Welwitschia (Gnetopsida) в оплодотворении участвует только один из двух спермиев. Двойное оплодотворение происходит у Ephedra и Gnetum, а также у покрытосеменных. У Ephedra благодаря слиянию одного спермия с яйцеклеткой и второго с брюшной канальцевой клеткой, а у Gnetum — двух спермиев с двумя клетками женского гаметофита образуются две зиготы, из которых далее развивается чаще всего только одна. Двойное оплодотворение покрытосеменных протекает иначе (см. рис. 11.188). Когда пыльцевая трубка добирается до яйцевого аппарата женского гаметофита, она изливает свое содержимое в одну из синергид, которая при этом разрушается. Отмирает и ядро пыльцевой трубки, а оба спермия или их ядра перемещаются далее, вероятно, амебоидно1. В результате слияния одного спермия с яйцеклеткой возникает зигота. Второй спермий сливается с центральной клеткой зародышевого мешка, где его ядро объединяется с вторичным ядром зародышевого мешка или с обоими полярными ядрами, образуя триплоидное ядро эндосперма, из которого развивается типичная питательная ткань семени покрытосеменных. Эндосперм отсутствует, к примеру, у орхидей, а у Оnаgraceae эндосперм диплоидный из-за наличия только одного полярного ядра.
1 Пыльцевая трубка прорастает через разрушающуюся синергиду в щель между яйцеклеткой и центральной клеткой зародышевого мешка, где лопается на конце, изливая спермин (не ядра!) непосредственно на яйцеклетку и центральную клетку зародышевого мешка. Ни спермин, ни ядра не перемещаются. — Примеч. ред.
Семя
После оплодотворения из семязачатка развивается семя, состоящее обычно из семенной кожуры, питательной ткани и зародыша. Семенная кожура (теста) образуется из одного или двух интегументов и может быть устроена весьма по-разному2. Обычно из наружной эпидермы наружного или единственного интегумента возникает экзотеста, из внутренней эпидермы — эвдотеста, из имеющихся в разном числе (но они могут и отсутствовать) промежуточных слоев — мезотеста. На зрелых семенах снаружи, как правило, можно заметить место обрыва семяножки — семенной рубчик, а в семенах, образующихся из ана- и кампилотропных семязачатков, — семенной шов, содержащий проводящий пучок (рис. 11.193).
2 Теста формируется из единственного — наружного — интегумента. Из внутреннего интегумента развивается тегмен. — Примеч. ред.
Рис. 11.193. Семя и его прорастание (Ricinus communis): А — вид с брюшной стороны; В — медианный, С — трансверсальный продольные срезы семени (А — С 2х); D — проросток (натур, величина); с — карункула (элайосома); е — эндосперм; h — рубчик; hk— гипокотиль; к — семядоли; г — корешок; га — семенной шов; t — семенная кожура
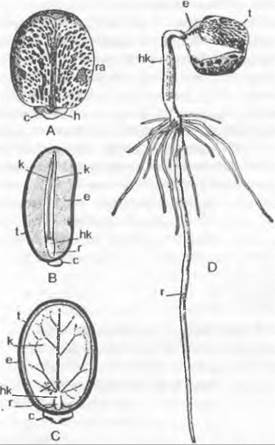
У саговников и Ginkgo наружная зона единственного интегумента превращается в мясистую и чаще всего ярко окрашенную саркотесту, а внутренняя часть — в
твердую склеротесту1. Интегумент хвойных дает жесткую семенную кожуру2. У Gnetopsida семенная кожура очень тонкая и в зрелом семени так сжата, что ее клеточная структура едва различима. У покрытосеменных с двумя интегументами в образовании семенной кожуры могут принимать участие оба или только один интегумент. В обоих случаях возможна дифференциация семенной кожуры на мясистую саркотесту и жесткую склеротесту (например, Magnolia сеае, Paeoniaceae, Púnica granatum). При участии двух интегументов граница между сар- ко- и склеротестой отнюдь не должна совпадать с границей между интегументами. У многих покрытосеменных с семязачатками, имеющими два интегумента, семенная кожура развивается только из одного, а другой обычно внутренний интегумент редуцируется. При наличии только одного интегумента семенная кожура развивается из него.
1 В склеротесту дифференцируется средняя часть интегумента. Внутренняя часть становится эндотестой или паринхотестой бумажистой консистенции. — Примеч. ред.
2 В семенной кожуре хвойных также можно различить слои. — Примеч. ред.
Для семенной кожуры покрытосеменных важно еще одно различие. Если она возникает из наружного интегумента, ее обозначают как теста, а если из внутреннего — то как тегмен.
Кроме дифференциации семенной кожуры на сарко- и склеротесгу можно обнаружить и другие вариации, особенно у покрытосеменных. Так, например, мясистыми могут быть тольконекоторые участки в целом жесткой семенной кожуры. Если это происходит в области перехода семяножки в интегумент, то возникает ариллус, который может почти полностью окружать семя (например, Еuonymus), быть лопастным (например. Мyristica; см. рис. 11.218) или дифференцироваться в заполненный воздухом поплавок (например, Nymphaea; рис. 11.194). Если семенная кожура мясистая в области микропиле, то говорят о карункуле (см. рис. 11.193), а если она мясиста в области семенного шва, — о строфиоли. Такие дополнительные образования — богатые жиром, белком или сахаром элайосомы (рис. 11.194) способствуют распространению семян муравьями (например, Соrydalis, Chelidonium). Часто семенная кожура ослизняется (например, у разных Brassicaceae, Linum, musía, Plantago), образуя миксотесту. На сухой тесте могут вырастать волоски (например, Epilobium,хлопчатник — Gossipium) или крыловидные придатки (например, Zanonia) (рис. 11.194) В нераскрывающихся плодах, где защитную функцию часто берет на себя стенка плода (например, Apiaceae, Asteraceae, Роасеае), семенная кожура обычно очень тонкая.
Рис. 11.194. Семя и его развитие: А, В — семязачатки разного возраста (70х); С — зрелое семя (9х) Epilobium angustifolium. Семена: D — Zanonia javanica (Cucurbitaceae, семя крылатое, 1/2х), Е — Papaver rhoeas, F — Pseu dofumaria alba, G — Chelidonium majus с элайосомами (e), а также H — Nymphaea alba с меш ковидным ариллусом (увеличено); f — семяножка; h — рубчик; m — микропиле; s — семязачаток
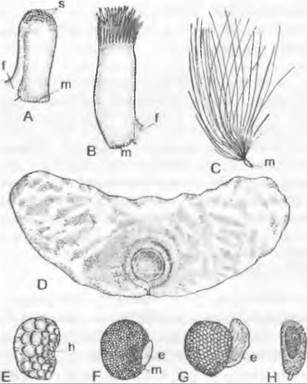
Семена сильно различаются размером. Так, семена сейшельской пальмы (Lodoicea: Агесасеае) весят несколько килограммов, в то время как семена Orchidaceae — всего несколько тысячных долей миллиграмма. Объяснить особенности размера, формы семян и характер их поверхности можно только в связи с их приспособлениями к распространению, прорастанию и устройством проростков.
Питательная ткань семени представлена чаще всего эндоспермом. У голосеменных гаплоидная ткань массивного женского гаметофита дает первичный эндосперм. Ткань нуцеллуса различима в зрелых семенах только в виде сильно спрессованной пленочки. У покрытосеменных возникает вторичный эндосперм, чаще всего триплоидный благодаря оплодотворению вторичного ядра зародышевого мешка одним из двух спермиев. Нуцеллус либо вообще неразличим, либо едва заметен, но в некоторых группах растений может служить и питательной тканью.
Вторичный эндосперм покрытосеменных обычно образуется по нуклеарному типу. В результате свободных делений ядер может образоваться более 2000 ядер, расположенных сначала в постенном слое цитоплазмы увеличивающегося зародышевого мешка (рис. 11.195). Лишь позднее начинается образование клеточных стенок (см. рис. 2.33). В целлюлярном типе образования эндосперма клеточные стенки образуются уже при первых делениях ядер, в гелобиальном типе развитие эндосперма в верхней части зародышевого мешка идет по нуклеарному, а в нижней части — по целлюлярному типу (см. рис. 11.223).
Рис. 11.195. Развитие зародыша и вторичного эндосперма у покрытосеменных: А — зигота; В — F — развитие подвеска (суспензора, s) и молодого зародыша с гипофизой (h), а также зачатков гипокотиля (hk) и семядолей (k); G — Н — зародыши с гипофизой (h), зачатками семядолей (к) и апикальной меристемой побега (m) (А — G — Rosopsida: Capsella bursa-pastoris Н — Uliopsida: Alisma plantago-aquatica) (ok. 200x); J — К — молодой зародыш (e) с суспензором в нуклеарном и целлюлярном эндосперме (es) (Lepidium sp. и Ageratum mexicanum) (увеличено); L — из эндосперма через микропиле выросла трубковидная ветвящаяся гаустория, которая местами прилегает к стенке завязи (w), местами — к семяножке (f); в зародышевом мешке виден также зародыш с суспензором (продольный срез молодого семени Globularia cordifolia, увеличено)
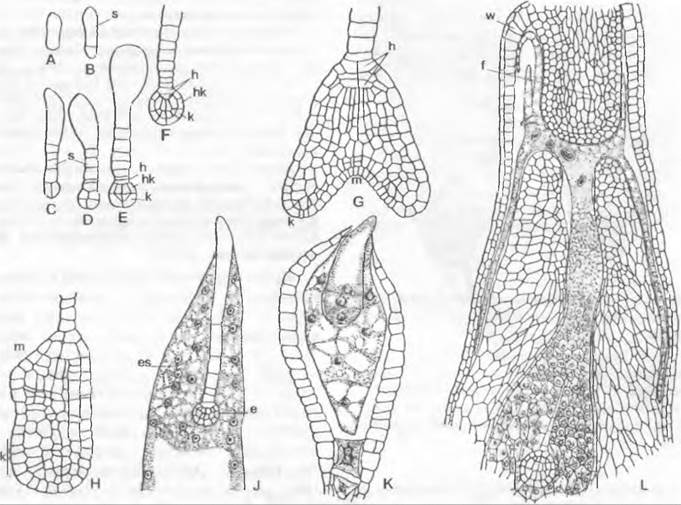
Образующийся эндосперм (и зародыш) поглощает питательные вещества. Прежде всего растущий зародышевый мешок вытесняет нуцеллус, большая часть которого или весь он расходуется на питание зародышевого мешка. Иногда из эндосперма или из суспензора зародыша в окружающие ткани внедряются трубковидные гаустории.
В некоторых семенах, например, у мускатного ореха и пальмы Агеса, от нуцеллуса, а у некоторых Аnnоnасеае от интегумента, в эндосперм врастают складки ткани, разделяя его бороздами. Таким образом возникает руминированный эндосперм (см. рис. 11.218).
Рис. 11.196. Развитие зародыша у Cycadopsida (Zamia floridana): A — свободное деление ядер (12х); В — образование клеточных стенок и ткани в основании зародыша (18х); С — начинающаяся дифференцировка предзародыша в суспензор (с продольно вытянутыми клетками) и базальный зародыш (22х)
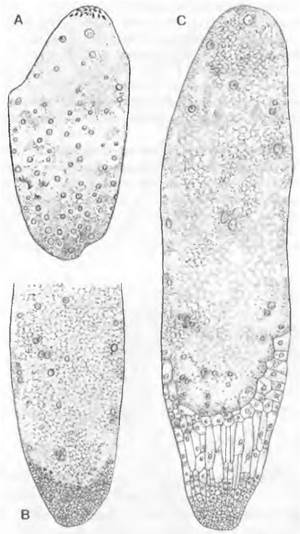
Кроме вторичного эндосперма у покрытосеменных питательной тканью может также служить нуцеллус. Подобного рода перисперм имеется в дополнение к эндосперму, к примеру, у Nymphaeaceae, Pipera- сеае (см. рис. 11.217) и Zingiberales, и как единственная питательная ткань — у Саryophyllales. Наконец, возможно накопление питательных веществ в самом зародыше, в частности в мясистых семядолях (например, Fabaceae, Quercus, Jugions, Aesculus), бывают семена вообще без эндосперма (например, Orchidaceae)1.
1 Семена орхидных вообще не содержат питательной ткани. — Примеч. ред.
Запасные вещества семени — это крахмал, белки или масла, откладываемые внутри клеток, либо резервная целлюлоза в клеточных стенках2. Соответственно этому эндосперм или другие запасающие ткани мучнистые, как у злаков, маслянистые, как у Cocos, или твердые, рогоподобные или каменистые, как у многих Liliales и некоторых пальм (например, Phytelephas: «растительная слоновая кость»).
2 На самом деле — гемицеллюлозы. — Примеч. ред.
Развитие зародыша из зиготы может происходить по-разному (см рис. 3.1; 11.195). Сформированный зародыш чаще всего состоит из прилегающего к микропиле подвеска (суспензора), благодаря удлинению которого остальная часть развивающегося зародыша (собственно зародыш) погружается в питательную ткань. Суспензор и собственно зародыш связаны между собой несколькими клетками, так называемой гипофизой. В зрелом семени суспензор и гипофиза чаще всего уже не различимы. Собственно, зародыш состоит из зародышевого корешка, расположенного под семядольным узлом участка оси — гипокотиля, разного числа семядолей и меристемы побега с самыми молодыми зачатками листьев (почечки).
Во всех группах семенных растений с несколькими архегониями или яйцеклетками в одном семязачатке могут образоваться несколько и зигот, и зародышей (полизиготическая полиэмбриония). Правда, чаще всего в таких случаях полностью развивается только один из них, поэтому зрелое семя содержит только один зародыш.
У Cycadopsida (рис. 11.196) и Ginkgo вначале происходят свободные деления ядер, так что может получиться более 1000 ядер. Лишь затем начинается образование клеточных стенок. Зародыши в этих двух группах имеют по две семядоли. У Coniferopsida развитие зародыша может начинаться со свободных ядерных делений, а продолжаться пугем нормальных делений клеток, но развитие его может и сразу идти по целлюлярному типу. Зародыш чаще всего имеет несколько семядолей (рис. 11.197). Развитие зародыша у Gnetopsida целлюлярное, зародыши имеют по две семядоли. У Gnetum и Welwitschia зародыш имеет боковое выпячивание, внедряющееся пальцевидно в питательную ткань и служащее, вероятно, для всасывания питательных веществ.
Рис. 11.197. Семя и его прорастание у Соniferopsida (Pinus pinea): А — семя (продольный разрез); В — D — прорастание семени; зародыш и проросток с семядолями (к), гипокотилем, главным и придаточными корнями (w); е — первичный эндосперм; s — семенная кожура; х — выпяченный и разорванный зародышевый мешок
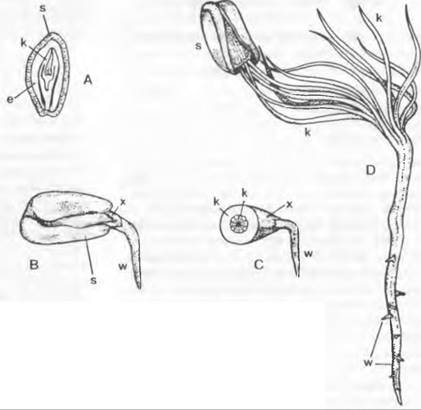
Особенность многих хвойных состоит в том, что из одной зиготы может возникать несколько зародышей (монозиготическая полиэмбриония). У Pinus (рис. 11.198) в зиготе путем двух свободных ядерных делений получается четыре ядра, которые расположены в одной плоскости на обращенном к микропиле конце. В результате дальнейших делений ядра располагаются в два яруса по четыре ядра в каждом. В результате образования клеточных стенок и дальнейшего клеточного деления получается четыре яруса по четыре клетки в каждом. Из самого нижнего яруса путем отхождения его четырех клеток друг от друга могут образоваться четыре зародыша, а вышерасположенный ярус благодаря удлинению и делению своих клеток становится суспензором. В других семействах и родах хвойных монозиготическая полиэмбриония либо не встречается, либо значительно большее число зародышей может возникать несколько иным путем.
Рис. 11.198. Монозиготическая полиэмбриония у сосны (Pinus): A — D — Pinus nigra (100x); E — P. banksiana (80x); e — зародыш; p — базальная пластинка; г — розетка; s — суспензор с s' вторичными клетками подвеска
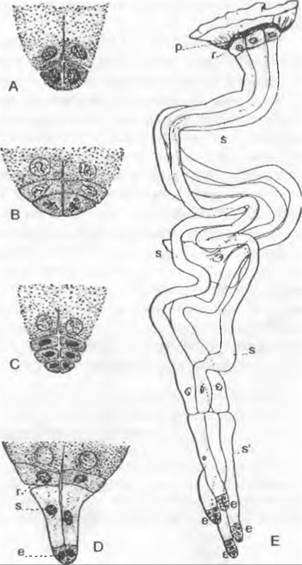
Монозиготическая полиэмбриония обнаружена также среди Gnetopsida — у Ephedra и Gnetum. У Ephedra из зиготы в результате трех клеточных делений образуется восемь клеток, и все могут развиться в зародыши. У Gnetum из зиготы возникают два первичных суспензора. Путем удлинения и ветвления, но без дальнейшего деления клеток получается обычно разветвленная трубчатая система, на многочисленных концах которой образуются многоклеточные вторичные суспензоры. Их концы развиваются в зародыши, вследствие чего из одной зиготы может подучиться очень много зародышей.
При монозиготической полиэмбрионии, как и при полизиготической, завершает развитие чаще всего только один зародыш. Таким образом, зрелое семя содержит только один зародыш
У покрытосеменных зародыш развивается по целлюлярному типу (см. рис. 11.195). После одного или немногих клеточных делений возникает так называемый предзародыш из одного ряда (но часто и Т-образно расположенных) клеток. Из него, с одной стороны, образуется суспензор, состоящий из клеток, расположенных всегда в один ряд, а с другой стороны — сначала шаровидный, состоящий из четырех или восьми клеток собственно зародыш, который затем развивается далее. Зрелый зародыш покрытосеменных имеет одну или две семядоли и может (например, у Роасеае; см. рис. 11 233) морфологически дифференцироваться далее на колеоптиль, колеоризу, щиток и т. п.
В зависимости от деталей хода клеточных делений и участия в развитии собственно зародыша и суспензора нижних и верхних клеток предзародыша различают разные типы эмбрионального развития. Наиболее часты Asteraceae- и Onagraceae-типы.
Сформировавшиеся зародыши покрытосеменных различаются по величине и степени дифференциации. Так, у Orchidaceae зародыши состоят из немногих клеток и совершенно недифференцированные. Наконец, у покрытосеменных возможно развитие зародыша без оплодотворения (агамоспермия; см. 10.1.3.3).
У Coniferopsida и Gnetopsida в распространении семян ветром и животными могут принимать участие структуры, не являющиеся частью семени. Например, имеющееся у семени Pinus крылышко — производное не семенной кожуры, а семенной чешуи (см. рис. 11.203). Сочные структуры у хвойных развиваются либо из почти всей женской шишки (например, «шишкоягоды» Juniperus-, см. рис. 11.209), либо мясистая семенная чешуя, так называемый эпиматий, окружает семя, либо мясистой становится часть оси шишки (Podocarpaceae), либо семя окружает разрастание оси ниже его (например, Taxus; см. рис. 11.210). У Gnetopsida функцию семенной кожуры берет на себя пара чешуевидных листьев, окружающих семязачатки, которые становятся жесткими. Наружная пара чешуевидных листьев может становиться мясистой (Gnetum) или разрастаться в крылья (Wel- witschiä). У Ephedra только с одной парой брактей вокруг семязачатков другие брак- теи стробила могут стать мясистыми и участвовать в распространении семени.
Плоды
У покрытосеменных семязачатки заключены в плодолистики. Поэтому по мере того как семязачатки после оплодотворения развиваются в семена, из плодолистиков, а иногда и из других частей цветка и цветоложа развивается плод, который, с одной стороны, заключает семена внутри себя и защищает их до созревания, а с другой стороны, участвует в распространении семян, высеивая их, либо вместе с ними отделяясь от растения. При развитии плодолистиков в плод они чаще всего изменяют форму, размер и анатомическое строение. Строение зрелого плода зависит от структуры гинецея, анатомии стенки плода, способа вскрывания плода при его созревании. При развитии плода из единственного плодолистика получается мономерный плод1. Апокарпные плоды развиваются из нескольких плодолистиков апокарпного гинецея. Из ценокарпных гинецеев развиваются разные ценокарпные плоды. Стенка зрелого плода (околоплодник) дифференцирована на наружную эпидерму (экзокарпий), внутреннюю эпидерму (эндокарпий) и разное число промежуточных слоев клеток (мезокарпий). Впрочем, разные слои стенки плода часто называют иначе. Если,
как, например, у костянок, имеется многослойная каменистая внутренняя часть околоплодника, то всю ее обычно называют эндокарпием. В зависимости от консистенции околоплодника различают сухие и сочные плоды. Среди сочных плодов есть ягоды с полностью сочным околоплодником и костянки с мясистой наружной частью и деревянистой внутренней частью стенки плода2. Эндокарпий может быть представлен мясистыми волосками, врастающими в виде пульпы в полость завязи (например, у Citrus). О вскрывающихся плодах говорят, если после созревания плод раскрывается и освобождается от семян. По особенностям вскрывания различают несколько типов плодов.
1 Мономерные плоды чаще всего рассматривают как один из вариантов апокарпных плодов. — Примеч. ред.
2 Типичные костянки — апокарпные плоды. Ценокарпные плоды с похожим околоплодником называют пиренариями. — Примеч. ред.
Если свободные плодолистики вскрываются по брюшному шву, они вентрицидные. Если ценокарпные плоды вскрываются по границам соседних плодолистиков, то говорят о септицидных (вскрывающихся по перегородкам) плодах. Дорсицидные (локулицидные; вскрывающиеся по створкам) плоды вскрываются вдоль на спинной стороне плодолистика. У порицидных плодов образуются поры — сравнительно небольшие отверстия, при вскрывании поперечной кольцевой щелью на верхушке ценокарпного плода получается коробочка с крышечкой.
Невскрывающиеся плоды с однородной сухой стенкой — орехи. Но невскрывающиеся плоды при созревании могут и распадаться на остающиеся замкнутыми части (распадающиеся плоды). Если ценокарпные плоды распадаются на отдельные плодолистики, то говорят о дробных плодах, а части плодов называют мерикарпиями. Если же разломы возникают в стенках самих плодолистиков, то говорят о членистых плодах.
Многообразие плодов как средства распространения семян повышается благодаря участию других органов или их частей, а также путем образования соплодий. У Nuphar цветоложе обрастает плод, развивающийся из верхней завязи, а у многих Rosaceae гипантий (Rosa) или цветоложе (Fragaria) становятся мясистыми. В образовании плода могут участвовать простой околоцветник (к примеру, мясистый в соплодии Morus; волосистый у Eriophorum), чашечка (например, сильно увеличенная и ярко окрашенная у Physalis alkekengi;представленная хохолком у Valerianaceae и Asteraceae), предлистья и кроющие листья (брактеи) (например, крыловидные у Carpinus или Humulus, трубковидные у Carex), плодоножки (например, мясистая у Anacardium occidentale), а также стеблевые и листовые органы соцветий (например, купулы Fagaceae, мясистые ось соплодия и брактеи у Ananas).
Ниже будет более подробно рассмотрена классификация плодов по их функционированию (по способам вскрывания) и анатомии (структуре стенки плода) (рис. 11.199). Плод определяют, как цветок ко времени созревания семян, поэтому можно классифицировать плоды и на основе морфологии цветка и принимать, к примеру, первичное подразделение плодов на монокарпические (из одного плодолистика), апокарпные и ценокарпные.
Рис. 11.199. Сухие вскрывающиеся (А — G) и невскрывающиеся (H —М) плоды. Мономерные плоды: А — листовка (Consolida regalis); В — боб (Laburnum anagyroides). Ценокарпныеплоды: С — стручок* (Chelidonium majus); D — септицидная коробочка (Hypericum perforatum); Е — дорзицидная** коробочка (Iris sibirica); F — коробочка с крышечкой (Anagallis arvensis); G — коробочка с порами (Antirrhinum majus). Орех лещины (Corylus avellana): H — общий вид; J — продольный разрез. Дробный плод: К — двукрылатка (Acer pseudoplatanus; с двумя односеменными мерикарпиями). Членистые плоды: L — членистый боб (Ornithopus sativus; мономерный плод с односеменными члениками); М — членистый стручок (Raphanus raphanistrum; ценокарпный ломкий плод) (А — ок. 4х; В, С, Н —К — в натур. вел.; D, Е — Зх; F — 2х; G — 3/4х; L, М — 2/Зх); а — рубец плодоножки; с — семядоля; I — проводящий пучок к семязачаткам; m — дорсальная средняя линия плодолистика; г— корешок; s — изогнутый семязачаток; t — семенная кожура; w — стенка плода
* Типичный стручок имеет ложную перегородку и встречается у крестоцветных (Вrassicасеае). Плод чистотела следует считать стручковидной коробочкой. — Примеч. ред.
** Локулинидная. — Примеч. ред.
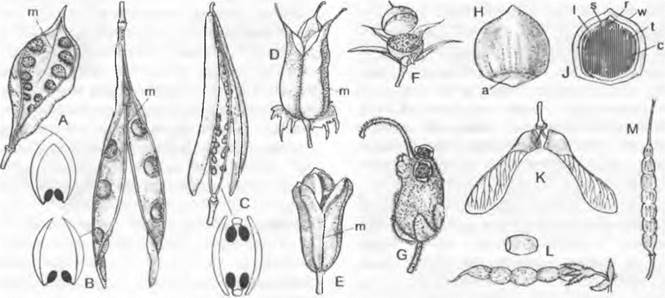
Вскрывающиеся плоды
У листовок свободные плодолистики апокарпного гинецея при созревании чаще всего сухие и вскрываются по брюшному шву. У Consolida, например, в цветке только один плодолистик и соответственно плод — однолистовка. У многих Magnoliidae, Paeonia, Delphinium, Trollius или Spiraea (см. рис. 11.255) в цветке образуются многочисленные листовки, так что в целом плод можно назвать сборной листовкой1.
1 Многолистовкой. — Примеч. ред.
Бобы — это также чаще всего сухие плоды, которые развиваются из одного плодолистика, но вскрываются с брюшной и спинной сторон. Один боб в цветке характерен, например, для Leguminosae (см. рис. 11.249), несколько — Magnolia (см. рис. 11.218).
Коробочки чаще всего сухие, они развиваются из ценокарпных гинецеев. По числу полостей
(гнезд) (развиваются из синкарпных или соответственно паракарпных гинецеев) и способу вскрывания (к примеру, по всей длине, только апикально или базально, по перегородкам или по створкам, порами или крышечкой) можно выделить разные типы коробочек (см. рис. 11.199). К коробочкам, строго говоря, относится и стручок Brassicaceae (см. рис. 11.265). Он состоит из двух паракарпно сросшихся между собой плодолистиков и вскрывается створками, отделяющимися от несущей плаценты рамки, на которые «натянута» ложная перегородка. Коробочкой является и активно разбрасывающий семена плод Geranium1 (см. рис. 11.243). Мясистые коробочки часто встречаются у тропических растений, а из центральноевропейских — у Еuоnуmus; коробочка, разбрасывающая семена при прикосновении, имеется у Impatiens.
1 Плод гераней представляет собой стеригму. — Примеч. ред.
Невскрывающиеся плоды.
Орехи с полностью одревесневшим невскрывающимся околоплодником могут образоваться как из отдельных плодолистиков апокарпного гинецея (например. Anemone, Ranunculus),так и из ценокарпного гинецея (например, Betula, Ulmus, Fraxinus)2. У Clematis и Pulsatilla удлиняющийся перисто-волосистый стилодий, а у Geum завернутый крючком стилодий принимают участие в распространении орешков. Функцию распространения орехов у многих Dipsacaceae берет на себя наружная чашечка3 (см. рис. 11.285). Тесно прижаты друг к другу или срастаются семенная кожура и стенка плода в зерновках Роасеае (см. рис. 11.233) и в семянках Asteraceae (см. рис. 11.286), которые также являются орехами. В сборных орехах4 несколько орехов апокарпного гинецея могут быть объединены благодаря мясистому цветоложу (Fragaria) или мясистому гипантию (Rosa) (см. рис. 11.255).
2 Орехами следует называть ценокарпные плоды. Похожие на них апокарпные плоды называют орешками. — Примеч. ред.
3 Обвертка из прицветничков. — Примеч. ред.
4 Многоорешках. — Примеч. ред.
К сухим невскрывающимся относят также и распадающиеся плоды. Образующиеся из ценокарпного гинецея дробные плоды разделяют на много (например, Malva) или только два (например, Acer: см. рис. 11.199) мерикарпия септицидно, т.е. по границам плодолистиков. У большинства Apiaceae (см. рис. 11.284) два мерикарпия остаются прикрепленными к центральной колонке (карпофору). Плоды с поперечно или вдоль разламывающимися плодолистиками называют ломкими плодами. Они могут развиваться из ценокарпных гинецеев (например, поперечно разламывающиеся членистые стручкиBrassicaceae, см. рис. 11.199, продольно разламывающиеся и к тому же перпендикулярно к разлому расщепляющиеся ценобии многих Lamiaсеае и Boraginaceae; см. рис. 11.277, 11.282) либо из единственного или из отдельных плодолистиков апокарпного гинецея (например, поперечно разламывающийся членистый боб некоторых Leguminosae: см. рис. 11.199).
Костянки характеризуются тем, что наружная часть околоплодника мясистая, а внутренняя деревянистая. Только из одного плодолистика образуется, в частности, костянка Prunus (слива, вишня и т. п.; см. рис. 11.255). Ценокарпные костянки1 образуются, например, у Jugions, Olea или Sambucas. У Cocos мезокарпий волокнистый, содержащий воздух (см. рис. 11.230) и позволяющий плодам эффективно распространяться водой. Малина и ежевика имеют сборные костянки2.
1 Пиренарии. — Примеч. ред.
2 Многокостянки. — Примеч. ред.
Яблоко (например, Malus; см. рис. 11.255) развивается из нижней завязи ценокарпного гинецея. Наружная часть его околоплодника мясистая, внутренняя кожистая или бумажистая.
Целиком мясистый околоплодник характерен для ягод. Многие Annonaceae, Actaea или финиковая пальма имеют ягоды из одного плодолистика. Ценокарпные ягоды характерны для Rites, Vitis, Vaccinium, Atropa или Convallaria. Плоды цитрусовых имеют мясистую пульпу. Если наружная стенка ягоды, как, например, у огурца и тыквы (Cucurbitaceae; см. рис. 11.258), довольно жесткая, то плод называют тыквиной.
Соплодия
Единицей распространения может быть целое соплодие. Примеры — шелковица и инжир (кувшинчатую форму у последнего имеют и соцветие, и соплодие), а также другие роды Моrасеае (см. рис. 11.257), или ананас с мясистым околоцветником, осью соцветия и плодами. У Tilia несколько орехов соединены с служащим крылом предлистом (см. рис. 11.266), у Arctium единицей распространения служит корзинка с крючковидными листьями обвертки. Наконец, единицей распространения может стать и вся надземная система побегов растения (так называемое «перекати-поле»), которую ветер обламывает у основания и гонит по степи, а в это время семена высыпаются. Движению способствует шаровидная форма растения.
Распространение плодов и семян
Распространение расселительных единиц (диаспор) в пространстве выполняет разные функции. При колонизации новых, т. е. не заселенных материнскими индивидами местообитаний, распространение на небольшие расстояния важно для сохранения популяции, а на большие расстояния — для основания новой популяции. Благодаря удалению от материнского индивида уменьшается внутривидовая конкуренция между потомками и родителями, а также между сестринскими индивидами. Правда, множество диаспор почти всегда оказываются в непосредственной близости от материнского индивида (см. 10.1.3.4). Распространение диаспор можно рассматривать и как стратегию избегания травоядных животных и патогенов, «поведение» которых зависит от плотности популяции растений, и, наконец, распространение диаспор является компонентом потока генов внутри и между популяциями.
Следует учитывать, что специализация механизмов распространения диаспор существенно меньше, чем при опылении. Так, многие диаспоры распространяются несколькими способами, т. е. полихорны. Разные способы распространения могут быть и в случае различия диаспор одного индивида (гетероспермия, гетерокарпия). Например, в корзинках некоторых видов Leontodón имеются семянки как с паппусом, так и без паппуса, а у различных Asteraceae самые наружные семянки не имеют паппуса и удерживаются в созревшей корзинке окружающими листочками обвертки. Некоторые растения рассеивают многие или все свои плоды по соседству благодаря активному росту плодоножек, помещая их, например, в трещины скал (Cymbalaria muralis) или в почву (например, земляной орех, Arachis hypo- gaea\ Trifolium subterraneum). Плоды и соответственно их части у Stipa и Erodium(см.рис.8.36) сами вбуравливаются в почву. Приспособления к распространению на большие расстояния обычно свойственны растениям аридных областей.
Наблюдения за распространением плодов и семян чаще всего показывают, что расстояния, на которые разносится подавляющее большинство диаспор одного индивида, довольно малы и часто лежат в пределах нескольких метров. Большое эволюционное значение имеет встречающийся в исключительных случаях занос диас
пор на большие расстояния, что доказывает пример изолированных океанических островов, сравнительно быстро заселяемых многими видами. Молекулярные методы анализа родства показывают, что географически удаленные таксоны растений могут быть близкородственными, а их ареалы возникли в результате дальнего заноса (англ.: long distance dispersial).
Важнейшие способы распространения — с помощью животных, ветра и активное разбрасывание диаспор. Распространение животными (зоохория) может принимать разные формы. При эндозоохории животные съедают диаспоры и затем выделяют их с испражнениями.
Признаками эндозоохории являются наличие у диаспор первичных аттрактантов (питательных веществ, таких, как углеводы, белки, жиры и масла; витамины, органические кислоты, минеральные вещества), вторичных аттрактантов (например, окраска и запах) и образований (склеротеста, жесткие части околоплодника), защищающих семена от разрушения ротовым аппаратом или в кишечнике животных-распространителей. Эти признаки могут быть как у семян, так и у плодов. В то время как мясистые семена и плоды животные чаше всего быстро съедают, из сухих диаспор они создают запасы. Первоначально, вероятно, важными распространителями семян и плодов были рыбы и рептилии (ихтио- и заврохория), позднее к ним присоединились птицы (орнитохория) и млекопитающие. Как и при опылении, при эндозоохории часто возникают тесные связи между растением и животным. Соответственно у мясистых диаспор можно обнаружить характерный синдром признаков приспособления к главному распространителю. При распространении птицами диаспоры чаще всего ярко или контрастно окрашены (красные, желтые, блестящие черные), не имеют запаха, среднего размера или мелкие, с мягкой кожурой и не опадающие. Примеры — мясистые семена (Magnolia, Paeoniu), костянки (Prunus avium, Ligustrum, Olea, Sambucus1), ягоды (Ribes, Vilis, Vaccinium), сборные плоды (Rosa, Rubus) и соплодия (Morus).Млекопитающие как эндозоохорные распространители диаспор играют заметную роль в тропиках. Из-за иных, чем у птиц, органов чувств и ротового аппарата распространяемые ими диаспоры чаще всего окрашены менее ярко, но зато имеют сильный запах; часто они крупнее, с более жесткой кожурой и опадают с растения. Примеры — костянки Prunus 1 pérsica,многие плоды типа «яблоко», часто имеющие сравнительно жесткую кожуру ягоды (какао, Citrus, хурма, Cucurbitaceae, бананы2) и соплодия (Ficus, Artocarpus). Похожи плоды, распространяемые рукокрылыми, но они остаются висеть на деревьях.
1 У последних трех родов — пиренарии. — Примеч. ред.
2 Плоды бананов распространяют рукокрылые. — Примеч. ред.
Среди сухих диаспор есть более мелкие, распространяемые преимущественно зерноядными птицами, и более крупные (например. Quercos, Fagus, Corylus, Jugions), которые собирают в основном грызуны для запасов, причем находят не все, что прячут, так что часть семян всегда благополучно прорастает.
Распространение муравьями (мирмекохория) основано на том, что муравьи собирают и растаскивают семена и плоды, имеющие характерные придатки (элайосомы) с привлекающими и питательными веществами.
Элайосомы образуются из разных частей семени (например, Asarum, Chelidomum, Corydalis, Viola, Cyclamen purpurascens, Melampyrum, Allium ursinum, Galanthus nivalis) или плода (например, Anemone nemoralis. Hepática, Lamium, Knautia). Мирмекохория встречается в лесах умеренного и тропического пояса.
Семена некоторых тропических Leguminosae с красно-черной контрастной окраской, по-видимому, имитируют элайосомы.
Сравнение близкородственных мирмекохорных и немирмекохорных видов показывает, что с этим способом распространения связаны и другие признаки. Так, у не имеющей элайосом Primula elatior семена созревают медленно в прямостоячих коробочках в высыхающих чашечках на длинном цветоносе, а у Р. vulgaris семена с элайосомами созревают быстро в коробочках в зеленых, ассимилирующих чашечках на коротких, полегающих на почву цветоносах.
Эпизоохория осуществляется благодаря тому, что диаспоры прикрепляются самым разным образом к поверхности животных.
Семена многих болотных и водных растений благодаря своим малым размерам могут прилипать вместе с илом к телу водных птиц и далеко разноситься ими. Возможность закрепления на теле животного повышается у семян или плодов, которые во влажном состоянии ослизняются и становятся липкими (например, Plantago, Juncus). Часто цепляются за животных диаспоры с железистыми волосками (например, Salvia glutinosa) и в особенности — с прицепками. Прицепки бывают волосками или эмергенцами плодолистиков (например. Medicago. Circaea. Galium aparine) или развиваются из столбиков (например. Geum urbanum1). чашелистиков (в том числе и из «наружной чашечки») или из листочков обвертки (например. Arctium, Xanthium). В то время как более мелкие цепляющиеся плоды распространяются преимущественно в шерсти мелких животных, крупные плоды с мощными прицепками (например, Tribulus. многие Pedaliaceae) приспособлены для закрепления и переноса на ногах крупных копытных.
1 У этого вида — из стилодиев. — Примеч. ред.
Особая форма распространения с помощью животных обнаружена у растений-баллистохоров. Пробегающие животные цепляют их эластичные стебли, которые, распрямляясь, катапультируют семена или плоды (например, разные виды с плодами-коробочками. Lamiaceae. Dipsacus).
Уже в раннее историческое время человек стал важным фактором в распространении диаспор (антропохория). Многие сорняки были занесены непреднамеренно с посевным материалом, шерстью и кормом для скота, а культурные растения были сознательно расселены по всему миру. В результате в некоторых регионах (например, в некоторых районах Новой Зеландии или Калифорнии) антропохоры доминируют над местной флорой. Замечательно, что диаспоры некоторых полевых сорняков в результате отбора так сильно уподобились по величине и другим особенностям диаспорам соответствующих культурных растений, что их практически невозможно отделить механическими методами от семенного материала (например, Cornelina, засоряющая лен).
Распространение с помощью ветра (анемохория) может быть косвенным, когда диаспоры при раскачивании стеблей вытряхиваются из соответствующих вместилищ (например, семена из коробочек — Papaver, семянки из корзинок — Bellis), или непосредственным, когда ветер сдувает и разносит диаспоры.
Во второй группе встречаются крошечные и легкие пылевидные семена (например, Orobanche, орхидеи), диаспоры с пузыревидными летучками (например, вздутые чашечки у Trifolium fragiferum), летучками из волосков (например, волоски на семенах, на столбике — Clematis2; ости — Stipa; паппус — Asteraceae), крылатые диаспоры (семена, крылатые орехи, дробные плоды-крылатки — Acer; соплодия — Tilia; зонтиковидная обвертка «наружная чашечка» — Scabiosa) и «перекати-поле».
2 Стилодии. — Примеч. ред.
Гидрохория состоит чаще всего в простом переносе диаспор водой. У дождевых баллистохоров энергия падающих дождевых капель благодаря лопатообразной форме сидящих на пружинящих плодоножках плодов преобразуется в метательные движения, когда выбрасываются семена из стручочков (например, Iberis, Thlaspí) или мерикарпии из чашечек (например, Prunella, Scutellaria). Впрочем, дождевые капли могут выбрасывать семена и непосредственно из соответствующим образом устроенных плодов.
Влажность может приводить, к примеру, у Sedum acre и многих Aizoaceae к гигрохастическо- му вскрыванию коробочек. При распространении потоками воды плавучесть диаспор достигается тем, что они не смачиваются или снабжены воздухоносным мешком (например, семена Nymphaea, мешочки разных видов осок, Сагех) или особой воздухоносной губчатой тканью (например, Cocos, Irispreudacorus. Potamogetón, Cakile marítima).
При автохории внешние агенты не используются.
В то время как многие диаспоры просто падают на землю (барохория), некоторые активно разбрасываются. Этот механизм основан на тургоре (например, у «взрывающихся» коробочек Impatiens, разбрасывании семян у Oxalis и у выстреливающего семенами более чем на 12 м бешеного огурца, Есballium; см. рис. 8.35) или на гигроскопических движениях (например, скручивание створок бобов у Dictanmus 1; коробочки-катапульты у Geranium; разбрасывание семян в результате сжатия коробочек у разных видов Viola). Автохорны и соответственно ахорны также те виды, которые активно зарывают в почву диаспоры возле материнского растения (Суmbalaría muralis, Arachis hypogaea).
1 Плод Dictanmus синкарпный. — Примеч. ред.
Все названные биологические особенности семян и плодов возможно лучше всего понять, учитывая биотоп видов. Это, например, отчетливо проявляется в том, что в нижнем травяном ярусе центральноевропейских лиственных лесов преобладает мирмекохория, у более высоких травянистых многолетников — эпизоохория, в кустарниковом ярусе — эндозоохория, в древесном ярусе — анемохория. Это соответствует вертикальному распределению агентов распространения (муравьев, млекопитающих, птиц, ветра).
Прорастание семян
Разнесенные тем или иным способом семена (голые или окруженные стенкой плода) попадают на поверхность или в самый поверхностный слой почвы. Здесь при подходящих условиях (см. гл. 7) и после иногда необходимого периода покоя по крайней мере некоторые семена прорастают. Однако часть семян, в особенности у видов ранних сукцессионных стадий, может оставаться в почве не прорастая, но сохраняя способность к прорастанию иногда весьма долгое время (доказано, что до 1 700 лет у Chenopodium album и Spergulaarvensis). Так возникают очень большие банки семян в почве. При прорастании семя поглощает воду и набухает, а его внутренние ткани разрывают семенную кожуру (или вдобавок стенку плода). Одновременно начинает расти зародыш, расходуя запас питательной ткани. При этом выделяются (в особенности семядолями) ферменты. Некоторое время все это происходит внутри семени. Зародыш всегда расположен в семени так, что его корешок обращен в сторону микропиле, поэтому при прорастании из семени через микропиле всегда сначала появляется корень проростка с гипокотилем (см. рис. 11.193; 11.197). При надземном (эпигейном) прорастании, удлиняющийся гипокотиль вытягивает семядоли из семенной кожуры и поднимает их над поверхностью почвы. При подземном (гипогейном) прорастании крупные, запасающие питательные вещества семядоли остаются в семени, и только эпикотиль появляется над ее поверхностью (например, Vicia faba, Pisum, Quercus, Jugions). Подобным образом прорастают семена и многих однодольных. Их единственная семядоля обычно представляет собой сосущий орган и извлекает питательные вещества из эндосперма.
Биология распространения и прорастания семян многих видов весьма специфична. Так, лишенные эндосперма семена Orchidaceae при прорастании должны вступить в контакт с микоризообразуюшим грибом. Также очень мелкие семена корневых паразитов (например, Orobanche) должны для прорастания попасть на поверхность корня хозяина, а семенам или плодам стеблевых паразитов требуются особые приспособления, например, повышенная клейкость, как у Viscum, что позволяет им удержаться на стебле хозяина. То же можно сказать и об эпифитах, которые отличаются либо мелкосемянностью (например, Bromeliaceae), либо клейкостью семян и плодов (например, Rhipsalis).
Необычно прорастание семян у живородящих представителей мангровых, например, у Rhizophoraceae (см. рис. 11.247). В их обычно односемянных плодах семя прорастает уже на материнском растении, и из висящего на дереве плода вырастает корешок и мощно развитый (у Rhizpphora до 1 м в длину) конусовидный гипокотиль. При созревании гипокотиль и корешок отделяются от семядолей и плода и падают вниз1. При этом тяжелый зародыш втыкается в мягкую илистую почву или смывается водой и укореняется после того, как она отступит.
1 Отделяются не только корешок и гипокотиль: зародыш, отбрасывая семядоли, выпадает из плода. — Примеч. ред.
Система Spermatophytina
Из многочисленных групп семенных растений, возникавших с верхнего девона, в настоящее время во флоре Земли еще можно встретить представителей пяти эволюционных линий. Это Cycadopsida (саговниковые), Ginkgopsida (Ginkgo), Coniferopsida (хвойные), Gnetopsida и Magnoliopsida (покрытосеменные, или цветковые, растения). Если от первых четырех групп, часто объединяемых общим понятием «голосеменные» (Gymnospermae), до настоящего времени сохранилось относительно мало видов (и только хвойные доминируют в некоторых типах растительности), то существующие с мела покрытосеменные (Ап- giospermae) представлены огромным множеством видов и являются важнейшим компонентом значительной части наземной растительности. По поводу родственных связей между различными группами семенных растений нет единого мнения. До относительно недавнего времени считали, что Gnetopsida являются ближайшими родичами покрытосеменных и, таким обра
зом, голосеменные представляют собой группу, парафилетическую по отношению к покрытосеменным. Однако самые различные работы по анализу последовательностей ДНК недавно показали, что вопреки этой концепции голосеменные все же представляют собой монофилетическую группу. Этим были возрождены более старые концепции, касающиеся филогении семенных растений. При последующем рассмотрении систематики и филогении семенных растений мы придаем современным пяти группам равные таксономические ранги.
1. Класс: саговники (Суcadopsida) (Palmfarne)
Ныне живущие Суcadopsida чаще всего имеют один кажущийся неразветвленным ствол, на котором по спирали расположены перистые листья, образующие терминальную розетку (рис. 11.200). Ствол бывает либо подземный, клубневидный, либо может достигать высоты 15 м.
Рис. 11.200: Cycadopsida: А — Cycas rumphii в Новой Гвинее, внешний вид. Мегаспорофиллы: В — Cycas revoluta; С — С. circinalis. D — Dioon edule; Е — Macrozamia sp.; F — Ceratozamia mexicana; G — Zamia skinneri
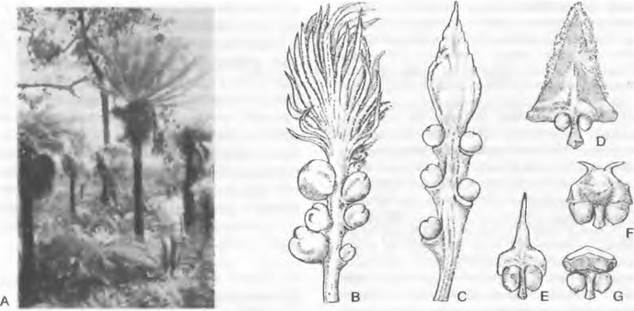
Древесина имеет очень широкие лучи. Она может состоять только из одного древесинного цилиндра, однако у некоторых видов вследствие развития добавочных камбиев снаружи от флоэмы во время вторичного роста в толщину могут возникать многочисленные концентрические древесинные цилиндры. Центр ствола занимает содержащая крахмал сердцевина. Во всех частях растения находятся слизевые ходы. У многих видов к тому же встречаются наряду с подземной корневой системой и растущие над поверхностью земли так называемые «коралловидные» корни, в которых поселяются цианобактерии из родов Nostoc, Calothrix и иногда Anabaena, фиксирующие атмосферный азот. Установлено, что растение может использовать фиксированный ими азот.
Листья чешуевидные или одиножды-, изредка дваждыперистые (Вowenia) фотосинтезирующие. Они имеют явно ксероморфное строение: кутикула толстая, а устьица чаще всего погруженные; листья очень жесткие. Их длина может быть от 5 см до 3 м. Листья отличаются длительным верхушечным ростом и у некоторых видов во время развития полностью или в области отдельных перышек улиткообразно закручены, как у папоротников.
Немецкое название этих растений — Palmfame (дословно «пальмы-папоротники») — связано с обликом, часто напоминающим пальму, и с иногда наблюдаемой закрученностью растущих целых листьев или отдельных перышек1.
1 В русскоязычной литературе, особенно в ненаучной, эти растения часто встречаются под названием «саговые пальмы»; интересно, что научное название класса происходит от родового названия Сусаs, которое, в свою очередь, происходит от греческого слова kykas — пальма. — Примеч. пер.
Эти растения всегда двудомные, но существование половых хромосом достоверно не установлено. Мужские цветки2 (рис. 11.201) имеют ограниченный рост и состоят из многочисленных расположенных по спирали тычинок3 (микроспорофиллов), чаще всего чешуевидных. На их нижней стороне развивается от 5 до примерно 1 000 микроспорангиев. Они часто собраны в группы и вскрываются благодаря экзотецию. Мужской гаметофит, развитие которого происходит отчасти в микроспорангии, а отчасти после высеивания из него, содержит наряду со сперматогенной клеткой еще три клетки. Сперматогенная клетка делится на два сперматозоида с расположенными по спирали многочисленными жгутиками. Имея диаметр до 400 мкм, эти сперматозоиды являются самыми крупными из всех сперматозоидов и в растительном. и в животном мире.
2 Микростробилы. — Примеч. ред.
3 Называть микроспорофиллы саговниковых тычинками ошибочно. — Примеч. ред.
Пыльцевая трубка имеет гаусториальную функцию и врастает в нуцеллус. Оплодотворение происходит по типу зоидогамии. У рода Мicrocyca образуются многочисленные дополнительные сперматозоиды в результате неоднократного деления другой — не сперматогенной — клетки.
Мужские цветки почти всегда занимают конечное положение на ветви. К концу цветения они смещаются в сторону разрастающейся боковой почкой. Так возникает внешне неразветвленный ствол, который на самом деле представляет собой симподий.
Женские цветки1 (рис. 11.201) состоят из мегаспорофиллов, также расположенных по спирали, и могут достигать 70 см. В роде Сycas спорофиллы состоят из ножки (рис. 11.200), по краям которой находится от двух до восьми семязачатков, и конечного участка, который может иметь вид перисто-раздельного листа, гребенчатой или пильчатой пластины. У остальных родов мегаспорофиллы имеют ножку и чешуевидную или щитовидную пластинку, на нижнем крае которой сидят всегда два семязачатка. Женские цветки чаще всего также имеют детерминированный рост; они расположены на конце ветви и после созревания семян, как и мужские цветки, отклоняются боковой ветвью. Только у Сусаs конус нарастания не исчерпывается при образовании стробила и прорастает через женский стробил (пролиферация). Семязачатки (см. рис. 11.180) имеют интегумент, который при созревании семян дифференцируется на наружный слой (саркотесту) розового или оранжевого цвета и внутренний слой (склеротесту)2. В нуцеллусе формируются пыльцевая и архегониальная камеры. Женский гаметофит состоит из большого (до нескольких тысяч) числа клеток и возникает в результате свободного деления ядер, за которым следует образование клеточных стенок. Чаще всего он развивает от двух до шести архегониев, только у Мicrocycas их может образовываться до 100. Величина яйцеклетки — до 6 мм.
1 Мегастробилы. — Примеч. ред.
2 Из интегумента развивается также располагающаяся под склеротестой паренхимная па- ренхотеста, которая идет на питание развивающегося эндосперма. — Примеч. ред.
Рис. 11.201. Cycadopsida: А, В — ♀ и ♂ цветки Encephalartos altensteinii (уменьшено); тычинки: С — Cycas circinalis (примерно 2х); D — Zamia integrifolia (примерно 5х) с группами микроспорангиев (Е, ок. 15х); F — поперечный разрез стенки вскрывшегося микроспорангип Stangeria paradoxa с экзотецием (примерно 80х)
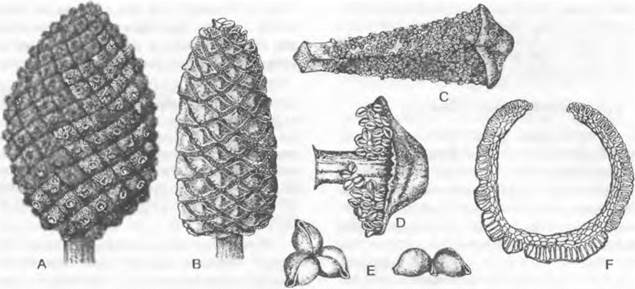
В ископаемом виде Суcadopsida достоверно известны с нижней перми. Ранние представители этой группы, как, например, юрский род Веania, имели женские цветки, в которых мегаспорофиллы были расположены значительно более рыхло, чем у современных саговниковых.
Пыльцу при опылении переносит ветер, однако отчасти и жуки. Для одного вида рода Zamia удалось показать, что жуки после завершения своего развития в мужских цветках летят, нагруженные пыльцой, к женским цветкам и опыляют их. Для представителей рода Encephalartos установлено опыление различными видами жуков. Пыльца, попавшая в каплю, выделяемую из микропиле, при ее высыхании втягивается в пыльцевую камеру, которая замыкается снаружи и соединяется с архегониальной камерой. Между опылением и оплодотворением может проходить до шести месяцев.
Суcadopsida включает около 140 видов из 11 родов и 4 семейств. Семейство Сусаdасеае (1 род/около 40 видов, тропики Старого Света) с единственным родом Сусаsхарактеризуется пролиферирующими женскими цветками. Некоторые виды имеют листовидные мегаспорофиллы. Stangenaceae (1/1, Южная Африка) и Воweniaceae (1/2, северо-восточная Австралия) имеют прилистники с отчетливым жилкованием. Воwenia имеет дважды перистые листья. Zamiaceae (8/около 114, тропики, субтропики, чаще всего Южного полушария) представлены родами Lepidozamia (2 вида) и Macrozamia (около 14) в Австралии, Encephalartos (около 35) в Африке и
Dioon (около 10), Ceratozamia (около 10), Zamia (около 40), Chigua (2) и Microcycas (1) в Америке, в том числе на Кубе (Microcycas). Большинство видов растет в тропических или субтропических лесах, или саваннах.
2. Класс: Ginkgopsida (Ginkgo)
Ginkgo biloba — единственный ныне живущий представитель Ginkgopsida — сильно разветвленное листопадное дерево с удлиненными и укороченными побегами. Его древесина имеет узкие лучи. Вееровидные листья (рис. 11.202) с дихотомическим жилкованием расположены по спирали. Этот вид двудомный и имеет половые хромосомы. Тычинки1 состоят из ножки и двух висящих на ее конце пыльников2 (рис. 11.202). Пыльники вскрываются благодаря эндотению. Мужской гаметофит состоит помимо сперматогенной клетки еще из четырех клеток. Сперматогенная клетка делится на два сперматозоида с расположенными по спирали многочисленными жгутиками.
1 Микроспорофиллы. — Примеч. ред.
2 Микроспорангиев, не гомологичных пыльникам покрытосеменных. — Примеч. ред.
После опыления пыльцевое зерно прорастает пыльцевой трубкой. С одной стороны, она имеет функцию гаустории и врастает в нуцеллус, из которого извлекает питательные вещества для мужского гаметофита. С другой стороны, она растет к архегониям, в которые должны быть высвобождены сперматозоиды. Оплодотворение осуществляют сперматозоиды (зоидогамия).
Многочисленные тычинки на оси образуют мужские цветки3, напоминающие сережки, располагающиеся в пазухах чешуевидных листьев на укороченных побегах.
Семязачатки чаще всего располагаются по два на концах вильчато разветвленной ножки (рис. 11.202). Как и мужские цветки, эта структура находится в пазухе чешуевидного листа на укороченной ветви. Семязачатки имеют один интегумент, который при созревании семени дифференцируется на наружную саркотесту, очень сильно пахнущую масляной кислотой, и внутреннюю жесткую склеротесту. Под склеротестой находится еще один мягкий слой семенной кожуры4. Нуцеллус имеет пыльцевую и архегониальную камеры. Женский гаметофит содержит хлорофилл и окрашен в зеленый цвет. Он состоит из нескольких сотен клеток, которые возникают после свободных ядерных делений, за которыми следует образование клеточных стенок. Он развивает чаще всего два или три архегония. Опыление у Ginkgo осуществляется ветром.
3 Микростробилы. — Примеч. ред.
4 Эндотеста. — Примеч. ред.
Рис. 11.202. Ginkgopsida: А — Baiera muensterana (рэт-лиас): вегетативный лист; семязачатки на оси <р цветка (несколько уменьшено); радиально расположенные, закрытые и открытые группы микроспорангиев на осях ♂ цветков (примерно 2х); В, С — Ginkgo biloba (современный вид): В — укороченный побег с ♂ цветком и молодыми листьями (натур, величина), редуцированные со спинной стороны, двучленные группы микроспорангиев (тычинки; увеличено), семязачатки ( ♀ цветки) и соответственно семена (несколько уменьшено); С — сперматозоид (примерно 200х)
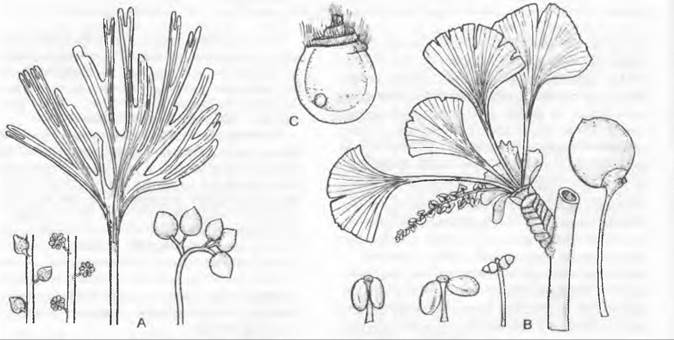
В ископаемом состоянии Ginkgopsida известны с нижней перми (например, роды Trichopitys и Sphenobaiera). Наибольшего разнообразия форм они достигли в юре и мелу. Род Ginkgoс видами, уже очень близкими к современному виду G. biloba. впервые встречается в нижней юре, а ископаемые остатки, не отличимые от G. biloba, известны с начала третичного периода. Этот вид, таким образом, является «живым ископаемым». У вымерших представителей Ginkgopsida (например, Baiera) листья часто бывали рассечены сильнее. Пыльники располагались радиальными группами, а женские цветки1 содержали большое число семязачатков.
1 Мегастробилы. — Примеч. ред.
Родина G. biloba, очевидно, Китай. Поскольку этот вид не менее 1 000 лет культивируется преимущественно в Китае и Японии в парках, окружающих храмы, и иногда дичает, его природный ареал и экология достоверно не известны. Предполагают, что природный ареал его находится в провинции Чжэцзян, где этот вид растет во влажных смешанных лесах.
3. Класс: хвойные (Coniferopsida)
Современные представители Coniferopsida развиваются из зародышей с двумя — многими семядолями (см. рис. 17.197) в разветвленные деревья или, реже, кустарники с моноподиальным стволом, на котором боковые ветви часто располагаются ярусно. Вторичная древесина часто содержит смоляные ходы, а лучи не многорядные1. Нередко имеет место дифференциация на удлиненные и укороченные побеги, а у видов некоторых родов (например, Phyllocladus, Sciadopilys2) укороченные побеги могут принимать вид более или менее уплощенных филлокладиев. Листья располагаются спирально, накрест супротивно или мутовчато, имеют параллельное жилкование или единственную среднюю жилку и обычно лентовидные, игловидные или чешуевидные. Есть как вечнозеленые, так и листопадные (летнезеленые) виды. Цветки3 однополые. Бывают как однодомные, так и двудомные виды.
1 Основная масса лучей — однорядные («узкие лучи»). Многорядные («широкие») лучи, если есть, содержат по одному радиальному смоляному ходу. — Примеч. ред.
2 Строение укороченных побегов Sciadopitys очень своеобразно, но не имеет никакого отношения к филлокладиям. — Примеч. ред.
3 У хвойных нет цветков, а есть микростробилы и шишки. — Примеч. ред.
Шишковидные мужские цветки располагаются поодиночке или рыхлыми группами (рис. 11.203). Тычинки4 располагаются по спирали или изредка накрест супротивно и имеют на нижней стороне от 2 до 20 микроспорангиев, часто срастающихся между собой. Изредка (Taxus) микроспорангии расположены радиально на конце ножки. Микроспорангии вскрываются благодаря экзотецию. Пыльцевые зерна могут иметь воздушные мешки, возникающие вследствие расслоения эктэкзины (рис. 11.203). Мужской гаметофит наряду со спермиогенной клеткой содержит еще 2 (Cephalotaxaceae, Cupressaceae, Sciadopityaceae, Taxaceae), 4 (Pinaceae) или от 10 до примерно 40 (Arauca- riaceae, Podocarpaceae) клеток. Сперматозоиды не образуются; два неподвижных спермия, возникающих из спермиогенной клетки, доставляются к архегониям пыльцевой трубкой (рис. 11.204) (= сифоногамия).
4 Это микроспорофиллы, которые нельзя отождествлять с тычинками. — Примеч. ред.
Рис. 11.203. Pinus: А — F. К — Р. sylvestris, G — I — Р. mugo: А — цветущий и семяносящий побег, в пазухе опадающих чешуевидных листьев располагаются двухиглые укороченные побеги; Е — ♂ цветки (G — продольный срез), тычинки с 2 микроспорангиями (Н — продольный, J — поперечный срезы); К — пыльцевое зерно с 2 воздухоносными мешками; В — ♀соцветие с комплексами из кроющих и семенных чешуй (С — сверху, D — снизу), из которых однолетние еще зеленые, и двухлетняя зрелая, раскрывающаяся шишка (А), несущая по 2 крылатых семени (F) на верхней стороне уже одревесневших к этому времени комплексов чешуй (А — несколько уменьшено, В — F увеличено, G — 10х, Н — 20х, J — 27х, К — 400х)
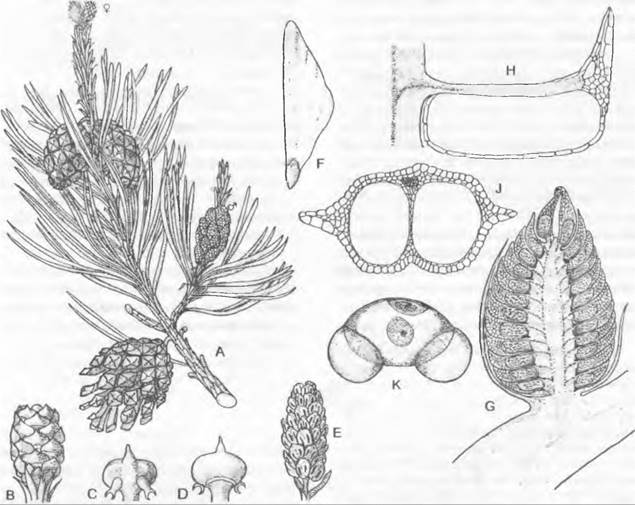
Рис. 11.204. Прорастание пыльцы и оплодотворение у Coniferopsida: А, В — развитие ♂ гаметофита в пыльцевом зерне и пыльцевая трубка: проталлиапьные клетки (р), ядро вегетативной клетки (клетки пыльцевой трубки) (к), антеридиальная клетка (а), дающая клетку- ножку (s) и спермиогенную клетку (sz), образующую в свою очередь 2 спермия (примерно 500х); С — сформировавшийся семязачаток с интегументом (i), пыльцевой трубкой (ps), а также архегониями с шейкой (h), брюшной канальцевой клеткой (Ь) и яйцеклеткой (е) (увеличено); D — пыльцевая трубка с 2 спермиями (sp1, sp2), а также ядрами клетки трубки (рк) и клетки-ножки (sk) возле яйцеклетки (ядро яйцеклетки ек, остаток шейковой клетки h); Е — слияние ядра яйцеклетки с ядром спермия, остальные ядра дегенерируют (367х)
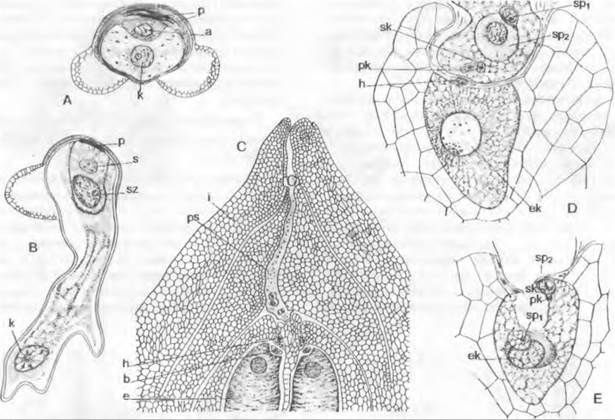
Женские цветки1 чаще всего собраны в характерные для хвойных, шишки (рис. 11.205). Они состоят из оси, на которой располагаются спирально или накрест супротивно кроющие листья отдельных цветков, называемые кроющими чешуями. Женские цветки, или семенные чешуи, представляют собой плоские органы, на верхней стороне которых находится различное число семязачатков (от 1 до примерно 20) Семенные чешуи несмотря на их чаще всего листовидную форму, следует считать модифицированными укороченными побегами, а женская шишка современных Coniferopsida является, таким образом, соцветием2. Семенных чешуй вышеописанного типа нет у Тахасеае и Cephalotaxaceae3.
1 Точнее — это аксиллярные комплексы, а не цветки. — Примеч. ред.
2 Поскольку у хвойных нет цветка, то не может быть и соцветия. — Примеч. ред.
3 Весьма модифицированы также семенные чешуи Podocarpales s. I. и Aracariales. — Примеч. ред.
Рис. 11.205. Pinaceae, Abies: А — побег со зрелыми шишками, справа уже частично рассыпавшейся (несколько уменьшено); В — ♂ цветок с чешуевидными листьями (b) и тычинками (sb) (примерно 2х); С, D — зрелый ♀ цветок с кроющей чешуей (d), семенной чешуей (ss) и 2 семенами (s); Е— семя с верхней (о) и нижней (u) стороны (несколько уменьшено)
Интерпретация семенной чешуи как женского цветка и, следовательно, женской шишки как соцветия вытекает из сравнения с кордаитами и Voltziales, ископаемыми представителями Сoniferopsida, а также, учитывая в типе пазушное ветвление семенных растений, из положения семенной чешуи в пазухе кроющей чешуи.
Кордаиты были деревьями высотой до 30 м, с развесистой кроной и пазушным ветвлением (рис. 11.206), вторичным утолщением и араука- риоидной поровостью трахеид вторичной древесины.
Лентовидные или ланцетные листья с дихотомически-параллельным жилкованием располагались спирально. Тычинки находились в напоминающих шишки цветках1 (рис. 11.206). В нижней части этих цветков располагались по спирали чешуевидные листья, а в верхней части находились также расположенные по спирали тычинки, несшие на вершине несколько микроспорангиев. Несколько таких мужских цветков рыхло располагались в пазухах кроющих листьев на удлиненных осях, выходивших из пазух обычных листьев. Сходно устроенные женские цветки располагались также в пазухе кроющего листа (рис. 11.206). За расположенными спирально чешуевидными листьями у них следовали также спирально расположенные мегаспорофиллы1, которые были палочковидными и на вершине несли по одному семязачатку (вследствие вильчатого ветвления изредка два или больше). Весь женский цветок интерпретируют как гомолог семенной чешуи современных Сoniferopsida. Наличие пыльцевой камеры в нуцеллусе указывает на то, что в отличие от современных Coniferopsida оплодотворение у кордаитов, вероятно, осуществлялось сперматозоидами. Кордаиты появились в карбоне как лесообразующие породы и вымерли в перми.
1 Микроспорофиллы в микростробилах. — Примеч. ред.
Рис. 11.206. Кордаиты (карбон-пермь): А — внешний вид Cordaites sp. (длина ок. 10 м); В — олиственный побег с расположенными в пазухах листьев соцветиями Cordaites laevis(уменьшено); С — два ♂ цветка Cordaianthus concinnus (ок. 2,5х); D — тычинка С. penjonii с группой терминальных микроспорангиев (ок. 10х); Е— ♀ цветок С. pseudofluitans с кроющим листом, стерильными чешуями и расположенными на длинных ножках семязачатками (1,5х)
Древовидные Voltziales, имевшие в древесине трахеиды с араукариоидной поровостью, жили с верхнего карбона до нижней юры. Листья у них были игло- или чешуевидными и иногда на вершине вильчато-рассеченными. Это были однодомные растения. Как и у кордаитов, женские цветки (рис. 11.207) состояли из пазушной оси с расположенными спирально или в одной плоскости стерильными чешуевидными листьями, а также одним или чаще всего несколькими семязачатками на ножках. Однако в отличие от кордаитов эти цветки, как и у современных Соniferopsida, были собраны в компактные шишки, которые, таким образом, надо трактовать как
соцветия. Тычинки Voltziales состояли из ножки и уплощенного конечного участка. Многочисленные микроспорангии располагались главным образом на адаксиальной стороне ножки. Voltziales можно рассматривать как организмы, очень близкие к исходным формам современных Coniferopsida.
Рис. 11.207. Voltziales: А — D — Lebachia piniformis (ротлиген): А — верхушка ветви; главный побег с вильчатыми листьями (1/Зх); В — торчащая вверх ♀ шишка с вильчатыми кроющими чешуями (1/2х); С — D — ♀ цветок сзади и спереди, кроющая чешуя (= кроющий лист, d), стерильные чешуи (s) и уплощенный прямой семязачаток (sa) с двураздельным интегументом* (5х); Е — Glyptolepis longibracteata (нижний триас): ♀ цветок с кроющей чешуей (d), стерильными чешуями и двумя анатропными семязачатками (схематично; 2х)
* Давно показано, что это не семя, а уплощенная семяножка, на адаксиальной стороне которой располагался обращенный семязачаток. - Примеч. ред.
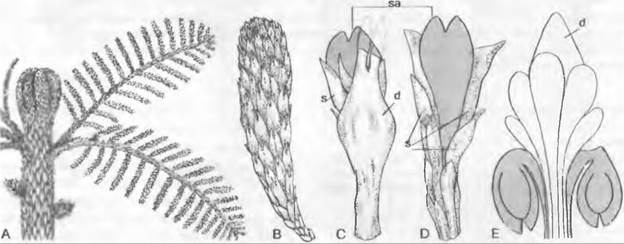
Семязачатки имеют один интегумент, который дифференцируется в твердую склеротесту.
Семена могут быть окружены мясистой оболочкой, которая, однако, никогда не возникает из семенной кожуры. Как и у Ginkgo, у хвойных многоклеточный женский гаметофит возникает путем свободных делений ядер, за которым следует образование клеточных стенок. Он может образовать до 60 архегониев. Насколько известно, все хвойные анемофильны. Пыльца либо попадает в специальную каплю, выделяемую из микропиле семязачатка, при высыхании которой втягивается в микропиле, либо оказывается внутри семязачатка в результате роста интегумента. В ископаемом виде хвойные можно проследить до триаса, а большинство современных семейств известны по ископаемым остаткам уже в триасе и юре.
Современные Соniferopsida насчитывают семь семейств. Рinасеае (12/около 200. чаще всего в умеренной зоне северного полушария) имеют расположенные по спирали игловидные листья и могут быть вечнозелеными или листопадными. Тычинки состоят из ножки, на нижней стороне которой находится 2 микроспорангия1. Пыль цевые зерна бывают с воздухоносными мешками. Семенные чешуи, более или менее несросшиеся с кроющими чешуями, несут два семязачатка, направленные микропиле к основанию чешуи. Растения всегда однодомные. Во флоре Средней Европы, это семейство представлено родами пихта (Abies), лиственница (Larix), ель (Picea) и сосна (Pinus)
1 Микроспорофиллы сосновых пельтатные. Два микроспорангия, находящиеся на нижней половине щитовидной пластинки микроспорофилла, прирастают к его черешку. — Примеч. ред.
Если лиственница — дерево листопадное, то хвоинки пихт, елей и сосен живут от 3 до 9 лет, изредка дольше. Хвоинка дифференцирована на пластинку и основание. В отличие от пихты, которая сбрасывает весь лист, у ели отпадает только листовая пластинка. Основание листа срастается со стеблем в так называемую листовую подушку.
Представители родов Pinaceae имеют либо только удлиненные побеги (например, Abies, Picea; рис. 11.205; 11.208), либо удлиненные и укороченные побеги. У вечнозеленых кедров (род Cedrus) и у лиственниц (рис. 11.208) каждый удлиненный побег в первый год несет зеленую хвою. На второй год на них вырастают укороченные побеги с пучками хвоинок; эти побеги могут нарастать несколько лет. У взрослых деревьев сосен хвоя располагается только на укороченных побегах. Молодые растения в первый или второй год дают и удлиненные побеги с зеленой хвоей, но после этого на них образуются только коричневые чешуевидные листья, в пазухах которых возникают укороченные побеги с немногочисленными чешуевидными листьями и пятью, тремя, двумя или даже только одной хвоинкой.
Когда пыльца попадает на семязачаток, женский гаметофит в нем еще не развит, а иногда даже не возникла и мегаспора. Соответственно между оплодотворением и опылением бывает продолжительный промежуток времени, в течение которого замыкается микропиле и скрывает прорастающую пылинку в семязачатке. У большинства видов Pinus между опылением и оплодотворением проходит один год, а семена из зрелых шишек высвобождаются только на третий год. В отличие от Pinus у ели от оплодотворения до созревания семени проходит один вегетационный период. При созревании семян шишка либо распадается (например, Abies), либо семена
высвобождаются, а шишка остается интактной (например. Picea, Pinus). Семена часто имеют кожистое крылышко, которое, однако, образуется не из семенной кожуры, а из отслаивающейся части семенной чешуи.
Рис. 11.208. Pinaceae: А — F — Picea abies: А — С — побеги с ♂ цветками, ♀ соцветиями и шишками (уменьшено); D — тычинка; Е— цветок с кроющей (d) и семенной (s) чешуями, а также семязачатком (sa) (увеличено); F — крылатое семя (в натур, вел.); G — Н — Larix decidua: G — удлиненный побег с несущими хвоинки короткими побегами (к) летом; Н — удлиненный побег с ♂ цветками, ♀ соцветиями и распускающимися почками укороченных побегов (к) весной (примерно в натур, величину)
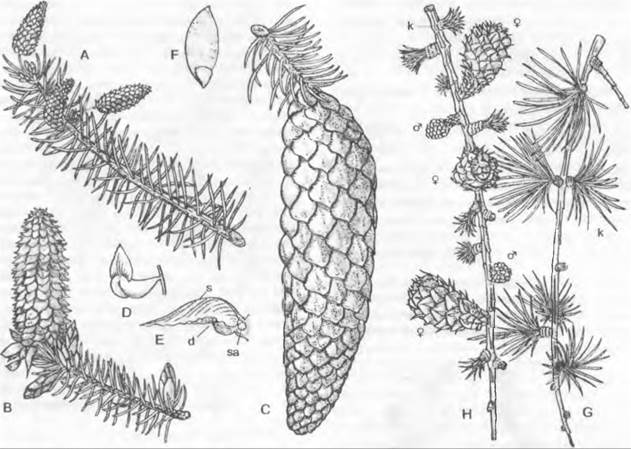
Экология среднеевропейских Pinaceae весьма различна. Если пихта белая (Abies alba) — дерево, достаточно требовательное в отношении почвы и климата, образует смешанные леса с буком, то формирующая однородные леса на северо-востоке Европы ель европейская (Picea abies) в Средней Европе в естественных условиях приурочена к среднегорному и субальпийскому поясам. Правда, из-за большой ценности для лесного хозяйства ель распространена гораздо шире в искусственных посадках. Лиственница европейская (Larix decidua) — светолюбивое дерево, которое часто встречается прежде всего в местах с континентальным климатом в Центральных Альпах вблизи верхней границы леса. Сосна обыкновенная (Pinus sylvestris) — светолюбивое, однако в остальном неприхотливое дерево с низкой конкурентоспособностью, легко выносящее как сухое и жаркое лето, так и сильные зимние морозы. Она встречается от равнин до верхней границы леса в горах и растет как на очень сухих, так и на очень сырых почвах. Чаще всего сосну обыкновенную можно встретить там, где более требовательные породы расти не могут. Два других вида этого рода — сосна горная (Р. mugo) и сосна «кедровая» европейская (Р. cembra)1 — обитают преимущественно в высокогорьях Средней Европы, а сосна черная (Р. nigra), распространенная преимущественно в Средиземноморье, доходит до Нижней Австрии.
1 В России этот вид и близкие к нему (например, Р. sibrica, Р. koraiensis) традиционно известны под названием «кедры». — Примеч. пер.
Araucariaceae (3/около 23. Южное полушарие, кроме Африки) имеют чаще всего широкие листья, сидящие спирально или накрест супротивно, а в древесине — трахеиды с расположенными в виде пчелиных сот окаймленными порами (араукариоидная поровость). Это однодомные либо двудомные растения. Тычинки несут от 4 до 20 микроспорангиев, а прочно срастающиеся с кроющими чешуями семенные чешуи — лишь один семязачаток. Описанный только в 1995 г. род Wollemia из Австралии явно наиболее примитивный род семейства — имеет большое сходство с его представителями, вымершими уже в третичный период. Соответственно его можно рассматривать как «живое ископаемое».
Некоторые представители Podocarpaceae (18/около 130, Южное полушарие, от тропиков до умеренных широт) имеют очень крупные листья. Виды этого семейства, преимущественно двудомные, отличаются в первую очередь тем, что женские шишки часто редуцированы до нескольких или иногда только одного комплекса из кроющей и семенной чешуи, в котором семенная чешуя окружает семя в виде мясистого покрова (эпиматия). Кроме того, стерильные семенные чешуи могут срастаться с осью шишки в мясистую «ножку»1. Относящийся к этому семейству Parasitaxus ustus из Новой Каледонии — единственный паразит среди хвойных. Его хозяин (Falcatifolium taxoides) также относится к Podoсаграсеае. В роде Phyllocladus (иногда выделяемом в самостоятельное семейство) функцию органов ассимиляции выполняют листоподобные, лопастные по краю филлокладии.
1 Рецептакулум. — Примеч. ред.
Cupressaceae (включая Taxodiaceae; 29/около 140; по всему земному шару, чаще всего в районах с умеренным климатом) имеют игольчатые или чешуевидные листья, расположенные по спирали, накрест супротивно или в мутовках. Растения эти однодомные или двудомные. Тычинки имеют от 2 до 6 микроспорангиев, а семенные чешуи, срастающиеся с кроющими чешуями, — от 2 (редко 1) до 20 семязачатков.
В Средней Европе Cupressaceae представлены двудомным можжевельником обыкновенным (Juniperus communis). Его женские шишки (рис. 11.209) состоят из многочисленных стерильных чешуевидных листьев, а также трех семенных чешуй, каждая с одним семязачатком. Эти семенные чешуи при созревании шишки становятся мясистыми и образуют шаровидные можжевеловые «ягоды». Можжевельник обыкновенный — неприхотливое растение, типичное для выгонов и кустарничковых пустошей. Выше альпийского пояса в горах и за полярной границей леса он представлен стелющимся подвидом — subsp. alpina. Мамонтово дерево (Sequoiadendron calífomicum) в Калифорнии имеет ствол диаметром более 8 м; его возраст свыше 3 000 лет. Sequoia sempervirens из прибрежных горных местностей в Калифор нии достигает высоты более 100 м. Metasequoia glyptostroboides, найденная в I940 г. в Китае, ранее была известна только в ископаемом состоянии в третичных отложениях и является еще одним примером «живого ископаемого». Этот листопадный вид осенью сбрасывает целиком укороченные олиственные побеги. Также веткопадным и имеющим реликтовый ареал является болотный кипарис (Taxodium disrichum), который образует обширные леса по болотам северного побережья Мексиканского залива. Он имеет выступающие из воды или ила выросты корней (пневматофоры), которые снабжают корневую систему воздухом.
Рис. 11.209. Cupressaceae, Juniperus communis: А — побег ♀ растения с соцветиями (D — с опылительными каплями, а также одно- и двулетними «шишкоягодами» — Е); В — побег ♂растения с цветками (С) (А, В — ок. 2/Зх, С — Е увеличено)
Основанное преимущественно на форме и положении листьев разделение Taxodiaceae (листья чаще всего игловидные, расположены спирально) и Cupressaceae (листья чаще всего чешуевидные, расположены накрест супротивно или в мутовках) необоснованно. Cupressaceae, вероятно, — монофилетическая группа, возникшая из Taxodiacae. Так как Cupressaceae — название более раннее, оно должно быть оставлено при объединении этих двух семейств.
Sciadopityaceae (1/1, Япония) с единственным видом Sciadopirys verlicillata отличаются наличием мутовчато расположенных игловидных филлокладиев1.
1 Это не филлокладии, а сросшиеся боками попарно игловидные листья укороченных побегов, развивающихся ярусами в пазухах чешуевидных листьев удлиненных побегов. — Примеч. ред.
У Cephalotaxaceae и особенно Тахасеае строение женских шишек существенно отличается от характерного для вышеописанных семейств. Женские шишки двудомных Cephalotaxaceae (1/около 6, районы умеренного климата Восточной Азии) состоят из немногих пар накрест супротивных чешуевидных листьев. В пазухе каждого из них находятся два прямостоячих семязачатка, из которых чаще всего развивается только один. Зрелое семя окружено мясистой тканью, которая возникает из ножки семязачатка. Семенные чешуи у Cephalotaxaceae неразличимы.
У также двудомных Taxaceae (5/около 25, районы с умеренным климатом Северного полушария), имеющих древесину без смоляных ходов, единственный семязачаток находится на уплощенных коротких боковых веточках над одной из накрест супротивных пар чешуевидных листьев (рис. 11.210). Положение семязачатков на боковых веточках трактуют как конечное или боковое. Как и у Cephalotaxaceae, у Taxaceae зрелое семя окружено мясистой тканью, которая возникает из ножки семязачатка.
Рис. 11.210. Taxaceae, Taxus baccata: А, В — цветущий ♂ и семяносящий ♀ побеги (с одним незрелым и двумя зрелыми семенами; 3/4х); С, D — ♂ и ♀ цветоносные побеги, каждый в пазухе хвоинки (2,5х); Е — ♀ цветоносный побег в продольном разрезе с микропиле (m), интегуменгом (¡), нуцеллусом (n), зародышевым мешком (е), зачатком ариллуса (а) и конусом нарастания первичного пазушного побега (v) (9х); F — семя в продольном разрезе, с ариллусом, семенной кожурой, эндоспермом и зародышем (2х)
Разделение Cephalotaxaceae и Taxaceae на два разных семейства крайне проблематично. Это можно видеть, например, из того, что род Атето- taxus относят то к Cephalotaxaceae, то к Taxaceae.
Интерпретация женских структур Taxaceae неоднозначна. Исходя главным образом из очевидного близкого родства Cephalotaxaceae и Taxaceae, а также из ближайшего родства с Cupressaceae/Sciadopityaceae, хорошо подтверждаемого данными по последовательности нуклеотидов ДНК, можно считать вполне вероятным, что женские структуры Taxaceae производны от шишки с кроющими и семенными чешуями, их строение не может служить основанием для часто практиковавшегося в прошлом выделения Taxaceae в отдельный порядок (Taxales). Тычинки Taxaceae также своеобразны. Так, у Taxus многочисленные микроспорангии располагаются по радиусам вокруг ножки тычинки, а у Pseudotaxus мутовки тычинок отделены друг от друга чешуевидными листьями.
В Средней Европе Taxaceae представлены тиссом ягодным (Taxus baccata), который встречается в местах с мягкой зимой, по осветленным участкам леса. Все части этого дерева кроме мясистого ариллуса ядовиты из-за наличия производных таксана, а древесина тисса — самая плотная из среднеевропейских деревьев. Таксол — вещество, содержащееся в Taxus brevifolia, — из-за его подавляющего митозы действия применяют при лечении рака.
Семейства Coniîeropsida можно объединить в три группы. Первую составляют Pinaceae, вторую Araucariaceae и Podocarpaceae, а третью Cupressaceae, Sciadopityaceae и Cephalotaxaceae/Taxaceae. Эта группировка, основанная на молекулярных данных, подтверждается структурой мужских гаметофитов. Более тесное родство существует, по-видимому, между второй и третьей группами.
4. Класс: Gnetopsida
Сохранившиеся доныне три рода Gnetopsida — Ephedra (35 — 45 видов, сухие области Евразии и Америки), Gnetum (около 30 видов, тропики всего земного шара) и Welwitschia (один вид, Юго-Западная Африка) — представляют собой древовидные растения, различающиеся в морфологическом плане. Древесина Gnetopsida имеет узкие лучи и у всех трех родов содержит сосуды1.
Гомология этих сосудов с сосудами Magnoliopsida спорна1. Виды рода Ephedra — кустарники, небольшие деревья или изредка даже лианы. Чаще всего лианы, но также и кустарники, и небольшие деревья встречаются в роде Gnetum (рис. 11.211). Welwitschia своим неразветвленным стволом, имеющим диаметр до 1 м, глубоко погружена в почву (рис. 11.211). Ephedra имеет очень мелкие чешуевидные листья, располагающиеся накрест-супротивно или в мутовках по три. У Gnetum листья простые, с перистым жилкованием, напоминающие листья многих двудольных, накрест супротивные. У взрослых растений Welwitschia имеется только одна пара супротивных лентовидных листьев с параллельным жилкованием (из двух пар, возникающих в процессе развития), которые в течение всей жизни растения нарастают у основания и отмирают на конце. Gnetopsida преимущественно двудомные.
1 Членики сосудов Gnetopsida имеют эфедроидные перфорационные пластинки, нигде за пределами класса более не известные. — Примеч. ред.
Мужские цветки1Gnetopsida имеют покров из одной или двух пар брактей и от одной до шести тычинок2 (рис. 11.211).
1 Цветков у Gnetopsida нет. — Примеч. peд.).
2 Это микроспорангиофоры. — Примеч. ред.
Рис. 11.211. Gnetopsida:
A — внешний вид молодого растения Welwitschia mirabilis с ♀ соцветиями (ок. 1/20х); В — ♂ цветок с кроющим листом (d) и двумя парами брактей (b), сросшимися между собой тычинками и стерильным семязачатком (ок. 7х); С, D — диаграммы ♂ и ♀ цветков; Е— J — Gnetum (Е, F, J — G. gnemon, G — G. costatum, H — G. monta- num); E — побег c ♂ соцветиями (3/8x); F — мутовчатые частные соцветия, снаружи с фертильными ♂, внутри со стерильными ♀ цветками (1/5х); G, Н — ♂ цветки с парой брактей (b); J — продольный срез ♀ цветка с двумя парами брактей, удлиненным интегументом, нуцеллусом и зародышевым мешком (увеличено); К — М — диаграммы ♂ и ♀ соцветий и цветков Gnetum; N — R — Ephedra altissima, N — ♂ побег (2/3x); O, P — ♂ частное соцветие и ♂ цветок (7,5х); Q, R — ♀ побег с незрелыми семенами (2/Зх) и терминальный ♀ цветок (2х); S, Т — диатоаммы ♂ и ♀ цветков Ephedra

У Ephedra мужской цветок имеет покров, состоящий из двух сросшихся основаниями друг с другом чешуевидных листьев, расположенных в медианной плоскости. Из этого покрова чаще всего выходит ножка, на конце которой находится несколько сросшихся между собой микроспорангиев. Поскольку эта ножка может быть на конце вильчато разветвленной или же один цветок содержит две ножки, иногда считают, что мужской цветок Ephedra состоит из двух чаще всего сросшихся между собой тычинок. Также состоящий из двух медианных брактей, но часто сросшихся в трубку покров мужских цветков Gnetum окружает ножку с одним или двумя микроспорангиями на конце. Эта структура вполне соответствует тычинке. У Welwitschia мужской цветок имеет покров из двух накрест супротивных пар брактей и мутовки шести тычинок, сросшихся основаниями между собой. На вершине каждая тычинка несет три сросшихся между собой микроспорангия. Иногда считают, что здесь мы имеет дело не с шестью, а скорее с двумя трехчленными тычинками. Мужские цветки Welwitschia всегда содержат на вершине рудиментарный семязачаток над рудиментарной парой трансверсальных брактей.
Микроспорангии вскрываются короткими клапанами благодаря экзотецию. Вследствие того, что утолщения стенок клеток экзотеция у Welwitschia инвертированы, пыльца при высыхании стенки микроспорангия выдавливается из него.
Мужской гаметофит содержит кроме сперматогенной клетки еще либо четыре (Ephedra), либо только две (Gnetum, Welwitschia) дополнительные клетки. Деление сперматогенной клетки приводит к образованию двух спермиев, а оплодотворение осуществляется посредством пыльцевой трубки (сифоногамия).
Женские цветки содержат прямостоячий семязачаток с интегументом, вытянутым в длинное микропиле1. Из микропиле выделяется опылительная капля. Семязачаток окружен одной трансверсальной (Ephedra) или двумя — медианной и трансверсальной (Gnetum, Welwitschia) — парами брактей, чаще всего сросшихся между собой (рис. 11.211).
1 Микропилярная трубка. — Примеч. peд.).
При созревании семени у Ephedra брактей цветка отвердевают и окутывают семена, а брактеи соцветия могут становиться мясистыми. У Gnetum наружная пара брактей становитсямясистой, а внутренняя — жесткой; у Welwitschia наружная пара брактей крыловидно разрастается, тогда как внутренняя твердеет. Женский гаметофит формируется путем свободных делений ядер, за которым следует образование клеточных стенок. Он состоит либо из одной мегаспоры (моноспорический: Ephedra), либо из всех четырех дочерних ядер, образующихся в результате мейоза в материнской клетке мегаспоры (тетраспорический: Gnetum, Welwitschia). Женский гаметофит может насчитывать более чем 1 000 клеток. У Ephedra развиваются типичные ахегонии, у Gnetum и Welwitschia архегониев нет, а яйцеклетки у видов обоих родов не отличимы от остальных клеток женского гаметофита. У Welwitschia от женского гаметофита навстречу пыльцевым трубкам отрастают трубковидные структуры.
У Ephedra и Gnetum происходит двойное оплодотворение. Слияние одного спермия с яйцеклеткой, а другого с брюшной канальцевой клеткой у Ephedra или же с любыми двумя клетками у Gnetum дает две зиготы. Которая из этих двух зигот разовьется в единственный зародыш семени, по-видимому, однозначно не предопределено.
Хотя некоторые виды Ephedra, вероятно, ветроопыляемые, по-видимому, у Gnetopsida в целом преобладает опыление различными насекомыми. Насекомых привлекают как запах, так и содержащие сахар опылительные капли.
Ископаемые Gnetopsida малоизвестны. Ископаемая пыльца, сходная с пыльцой современных гнетовых, известна с верхнего триаса, но принадлежность к Gnetopsida ископаемых макроостатков из нижней юры (например, Piroconites) или мела (Drewia, Eoanthus) сомнительна.
Предполагалось, что растущие только в пустыне Намиб растения Welwitschia mirabilis, отдельные экземпляры которых имеют возраст до 2 000 лет, являются реликтами с тех времен, когда климат там был более влажным.
С покрытосеменными часто сближали ископаемые Bennetitales (рис. 11.212). Стволы их были либо короткие неразветвленные и сильно узловатые (например, Williamsonia), либо относительно тонкие и разветвленные (например, WieIandieIIa). Одинождыперистые или простые листья располагались по спирали. Цветки были или однополыми, или — очевидно, у более продвинутых родов — обоеполыми1. У большинства видов цветки имели обертку из спирально расположенных кроющих листьев. Тычинки2 располагались в мутовках и были либо перисто-разветвленными и завернутыми на верхнюю сторону, либо нерасчлененными и листовидно уплощенными. Микроспорангии имели экзотеций и были по нескольку в виде синангия окружены двухлопастным покровом. Каждая тычинка несла несколько таких синангиев на адаксиальной стороне.
1 Гомология этих структур цветкам покрытосеменных не доказана. — Примеч. ред.
2 Микроспорофиллы. — Примеч. ред.
Рис. 11.212. Bennettitales:
А — Wielandiella angustifolia (верхний триас), реконструкция с ветвями, листьями и цветками или плодами (1/5х), В — ♀/♂ цветки Williamsoniella coronata (средняя юра) с листочками обвертки и тычинками, а также гинецеем (примерно натур, величина); С — лепестковидная тычинка (2х); D — клубневидный ствол, крона перистых листьев и цветочные почки Cycadeoidea sp. (реконструкция, сильно уменьшено); Е — продольный срез выступающего из женского цветка плода1Bennettites gibsonianus (нижний мел) с листочками обвертки, межсеменными чешуями и расположенными на ножках семенами (примерно 1/Зх); F — продольный срез группы микроспорангиев с экзотецием у Cycadeoidea dacotensis (ок. 16х)
1 Плодов у Bennettitales не было. — Примеч. ред.
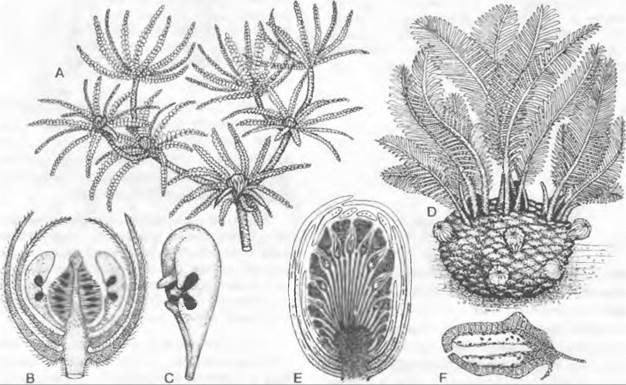
Вертикальные семязачатки имели стерильную обертку. В женском цветке или в верхней части обоеполого цветка чаще всего были многочисленные (до нескольких сотен) семязачатки, располагавшиеся по спирали вдоль удлиненной оси цветка и отделенные друг от друга стерильными межсемянными чешуями. Семязачатки с плюской и ножкой можно интерпретировать как сильно редуцированные мегаспорофиллы, а межсемянные чешуи — как стерильные мегаспорофиллы. Вероятно, Bennettiatales — по крайней мере, некоторые — опылялись насекомыми. Bennettiatales появились впервые в верхнем триасе и вымерли в мелу.
5. Класс: цветковые растения, покрытосеменные (Маgnoliopsida; Аngiospermoe)
Как показывает сравнение разных изданий учебника Э. Страсбургера, представления о родственных связях среди покрытосеменных (естественно, и среди других групп растений), а отсюда — и сама система цветковых растений постоянно изменяются. Есть три причины такого продолжающего и, конечно, еще далекого от завершения изменения системы. В ранний период систематики, после К. Линнея, особенно резко возросло число вновь открываемых видов вследствие интенсивной деятельности по сбору и изучению растений, прежде всего в ходе колонизации новых земель. Вместе с общим развитием технических средств постоянно увеличивались количество и характер изучаемых признаков растений. Если первые систематики имели в своем распоряжении только лупу, то развитие техники от простых микроскопов до современных электронных позволило проводить наблюдения со все большим разрешением, все более качественно и быстро характеризовать химический состав частей растений с помощью хроматографии и анализа ультраструктуры. В наше время для систематиков привычным средством стала уже разнообразная техника для анализа ДНК. Наконец, в ходе истории сильные изменения претерпели и сами подходы к интерпретации наблюдаемых данных. Если в начале систематики постулировали особую важность лишь некоторых признаков, то впоследствии возникла необходимость в тщательном пересмотре всех признаков с целью по возможности объективного выявления общего сходства и повторной кладистической обработки. При изучении последовательностей нуклеотидов участков ДНК применяют такие различные методы анализа, как методы объединения соседей, максимальной парсимонии (максимальной экономии) и максимальной вероятности (см. 11.1).
Карл Линней основал применяющийся и сейчас способ наименования растений, включающий название рода и дополняющее его название вида (видовой эпитет)1. Его система, основанная в первую очередь на числе и расположении тычинок, была очень искусственной. После К. Линнея были опубликованы различные системы, принимающие во внимание все признаки, учитывавшиеся в то время, например, системы М. Дцансона, А. Л. де Жюссьё, А. П. Декандоля, Дж. Линдлея, С. Л. Эндлихера, Г. Бентама и Дж. Д. Гукера, А. Брауна, А.Энглера, С. Е. Бесси, Х. Халлира, Дж. Б. Хатчинсона и Р. фон Веттштейна. До начала 90-х годов прошлого столетия были разработаны системы Артура Кронквиста (1981), Р. М.Т.Дальгрена (1975), А. Л.Тахтад жяна (1980, 1997) и Р.Ф. Торна (1992). Примечательно, что опубликование дарвиновской теории эволюции очень мало повлияло на развитие самой систематики. То, что до Ч. Дарвина считали сходством, впоследствии интерпретировали как родственную близость.
1 К. Линней широко использовал этот принцип бинарной номенклатуры, ставший впоследствии общепринятым, но не был его основателем. — Примеч. ред.
Представляемая здесь система в общем плане следует опубликованной в 1998 г. и основанной на признаках ДНК (rbcL, atpВ, 18SrДНК) системе «группы филогении покрытосеменных» (АРG) К. Бремера, М. В. Чейза, Р. А. Стивенса и др.
Эта система, доведенная только до уровня порядка, была модифицирована, главным образом в основании филогенетического древа, с учетом данных новых публикаций, в особенности последних, касающихся филогении в пределах порядков.
Главным основанием для представленной здесь системы являются результаты анализа ДНК. С постоянным увеличением числа проанализированных таксонов и секвенированных участков ДНК (гbсL, аtрВ, 18SrДНК, митохондриальные аtp1 и matR, фитохромы А и С) они приводят к очевидному согласию о родственных взаимоотношениях разных таксонов. Добавим, что полученные результаты в большинстве случаев были подтверждены и классическими признаками. Тем не менее понятно, что представленная здесь система будет изменяться и далее.
Данная система в значительной степени основана на признании строгой монофилии широко известных таксонов. Вследствие этого некоторые семейства, несмотря на их фенетическую отграниченноcть, теперь признаны не более, чем формальными группировками в составе других семейств (например, Асегасеае будут включены в Sарindасеае, Lеmnасеае — в Агасеае, Сhenopodiасеае — в Аmaranthасеае и т. д.). Признание их в качестве отдельных семейств привело бы к признанию парафилии остальных таксонов. Если бы, к примеру, считать и далее Lemnасеае самостоятельным семейством, то Агасеае было бы парафилетическим (см. 11.1). При этом сходство как окончательный субъективный критерий классификации не принимают во вни мание.
В значительной мере мы избегали употреблять формальные названия на уровнях выше порядка. Это обусловлено тем, что весьма непростая формализованная процедура присвоения названий должна применяться только тогда, когда достигнута существенная стабильность системы.
Признаваемые здесь порядки и неформальные группы более высокого уровня в неравной степени сходны по классическим признакам. Трудности, которые возникают при попытке найти классические признаки для определенных молекулярными методами родственных групп, могут в принципе иметь три основные причины. Так, возможно, что выделение какой-либо группы по молекулярным признакам окажется несостоятельным в ходе дальнейших исследований. Далее может оказаться, что общие признаки, возникшие в ходе быстрой фенотипической эволюции, становятся неразличимыми или же при дивергентной эволюции утрачиваются общие признаки в больших и старых родственных группах. Наконец, признаки многих групп не очень хорошо изучены. Здесь молекулярные гипотезы родства также имеют преимущество по сравнению с классической систематикой. Используемые в них признаки порядков и групп порядков далеко не всегда являются синапоморфиями.
Области распространения всех семейств, число их родов и видов даны в соответствии с различными обработками различных семейств и с очень различной точностью и достоверностью.
Эволюция и филогения покрытосеменных (обзор)
Покрытосеменные, включающие около 250 тыс. видов, в настоящее время — самая богатая видами группа растений, намного превосходящая все остальные группы. К важнейшим синапоморфиям покрытосеменных относятся: происходящие из одной материнской клетки членики ситовидных трубок и сопровождающие клетки флоэмы, тычинки с двумя латеральными парами гнезд пыльника, пыльники с гиподермальным эндотецием, пыльцевые зерна почти всегда без ламеллятной эндэкзины, трехклеточный мужской гаметофит, семяпочки, расположенные в замкнутых плодолистиках с рыльцем, стенки мегаспор без спорополленина, 8-ядерный женский гаметофит, а также двойное оплодотворение и образование вторичного эндосперма.
Вообще видовое богатство таксонов является результатом более высокой скорости видообразования и (или) более низкой скорости вымирания. Видовое богатство покрытосеменных имеет, вероятно, различные причины. На первое место здесь можно поставить возникшее уже у первых покрытосеменных опыление насекомыми (шире — зоофилию). Определенная специализация опылителей приводит к репродуктивной изоляции, результатом которой, в свою очередь, может быть высокая скорости видообразования. Кроме того, возможно, что опыление насекомыми и другими животными вообще повышает степень перекрестного опыления и тем самым ограничивает генетическое обеднение, угрожающее мелким популяциям. В результате могла бы снизиться степень вымирания. Очень богатые видами орхидные (около 18 500 видов) — несомненный пример тесной связи ярко выраженной специализации по биологии опыления с видовыми разнообразиями.
Репродуктивной изоляции и повышению таким образом скорости видообразования, по- видимому, содействует расположение семяпочек в замкнутом плодолистике, благодаря чему возможно распознавание чужой пыльцы не в момент оплодотворения, а еще при опылении. Таким образом снижается вероятность оплодотворения чужеродной пыльцой и межвидового скрещивания. Неоднократно наблюдалось, что скорость роста пыльцевой трубки коррелирует с генетическим сходством производителей пыльцы и семяпочки. Поэтому относительное подавление роста пыльцевой трубки при опылении растений из генетически расходящихся популяций также может приводить к репродуктивной изоляции в пределах вида и этим способствовать повышению скорости видообразования.
Участие плодолистиков в образовании единиц расселения (плодов) в общем, пожалуй, повышает дальность расселения видов и таким образом способствует репродуктивной изоляции путем географической изоляции, а тем самым — и повышению скорости видообразования. В то же время из-за увеличения дальности расселения, естественно, преодолевается пространственная изоляция популяций. Сочные плоды, распространяющиеся эндозоохорно. в эволюции покрытосеменных возникли, по-видимому, достаточно поздно. Эпизоохорно распространяющиеся плоды, напротив, известны уже у очень ранних ископаемых покрытосеменных.
Поскольку вторичный эндосперм покрытосеменных формируется только после успешного оплодотворения, их размножение эффективнее, чем у голосеменных. Данный факт, а также сокращение длительности гаметофитной фазы жизненного цикла способствовали, возможно, превосходству покрытосеменных в их конкуренции с разными группами голосеменных растений.
Чрезвычайное разнообразие вторичных метаболитов у покрытосеменных, очевидно, также влияет на скорость видообразования. Поскольку эти вещества большей частью обеспечивают защиту растений от травоядных животных и патогенных организмов, эволюция новых вторичных метаболитов может привести к возникновению относительно свободных от повреждающих факторов ниш, в которых возможны дивергентная эволюция и возникновение новых видов. Видовое разнообразие Аsterасеае (ок. 22 750 видов), вероятно, можно объяснить богатством у них вторичных метаболитов.
Наконец, покрытосеменные, выработав разнообразные жизненные формы, смогли занять жизненное пространство, не освоенное древесными и кустарниковыми растениями. Так, травянистость в сочетании с укороченным жизненным циклом позволяет заселять относительно нестабильные местообитания. Многообразие морфологии и, по-видимому, анатомии вегетативных органов также может способствовать ускорению видообразования.
Первые ископаемые покрытосеменные появляются в отложениях раннего мела, около 140 млн лет назад. Ископаемые остатки их пыльцы и листьев обнаружены в низких широтах с семиаридным или сезонно аридным климатом. Преобладающими элементами наземной растительности они впервые становятся в среднем, и верхнем мелу.
Датировка возраста покрытосеменных по «молекулярным часам» и исследования значительно более древних Gnetopsida, из которых до недавнего времени часто выводили покрытосеменные, допуская ближайшее родство этих таксонов, неоднократно приводили к предположению, что возраст покрытосеменных не менее 300 млн лет, т.е. они возникли значительно раньше мела. Хотя это и не исключено, но столь раннее появление покрытосеменных не подтверждено ископаемыми остатками. Правда, недавно описанные ископаемые остатки цветков относятся к самым верхам юры. Есть все основания думать, что заметная диверсификация покрытосеменных, представляющих собой явно более древнюю линию эволюции, впервые произошла в начале мела.
В основание эволюционного древа покрытосеменных (рис. 11.213) ставят несколько семейств, часто включаемых в Маgnoliidae, к которым относится только 4 % всех ныне живущих видов. Представители этих семейств — главным образом древесные формы, но есть и травы, как правило, с простыми листьями и эфирными маслами в шаровидных идиобластах. Цветки спиральные или циклические из немногих или многочисленных элементов, пыльцевые зерна однобороздные (моносулькатные), плодолистики большей частью свободные или гинецей состоит только из одного плодолистика. Базальной линией современных покрытосеменных недавно признали, на основании различных молекулярных данных, семейство Amborellaceae, представленное единственным видом, произрастающим в Новой Каледонии. Внешний вид очень мелких (< 5 мм) цветков Amborellaceae. состоящих из немногих элементов, хорошо соответствует палеоботаническим данным, показывающим все большее число очень мелких древнейших цветков. Таким образом, весьма вероятно, что первые покрытосеменные были мелкоцветковыми. Поскольку в древесине Amborella нет сосудов, то возможно, что эти столь характерные для огромного большинства покрытосеменных элементы появились уже после возникновения покрытосеменных. Вплотную к основанию эволюционного древа покрытосеменных находится также семейство Nymphaeaceae с распространенными в Центральной Европе кувшинкой (Nymphaea) и кубышкой (Nuphar). Представители более крупных семейств Magnoliidae — магнолия (Magnolia), лавр (Laurus) и перец (Piper). Относящийся к Magnoliidae и также произрастающий в Центральной Европе роголистник (Ceratophyllum), который несколько лет назад рассматривали как представителя базальной линии покрытосеменных, вероятнее является ближайшим родственником однодольных или так называемых настоящих двудольных (Eudicotyledonae).
Рис. 11.213. Предполагаемая филогения покрытосеменных по результатам анализа семи нукпеарных (18SrDNA, фитохромы: PHYA, PHYC), пластидных (rbcL, atpB) и митохондриальных (atp1, matR) последовательностей ДНК методом максимальной экономии
Magnoliidae— парафилетическая базальная группа, из которой возникли остальные покрытосеменные. Явным признаком того, что это остаток некогда более богатой видами группы, считается их фрагментированный ареал — преимущественно тропические и субтропические области всех континентов.
Однодольные (Liliidae, Monocotyledonae), первый большой блок остальных покрытосеменных, содержат около 22 % от общего числа их видов. Самые древние несомненные ископаемые остатки этой группы датируются возрастом около 90 млн лет. Благодаря своим характерным признакам — преобладанию травянистых жизненных форм, наличию только одной семядоли, большей частью рассеянно расположенными на поперечном срезе стебля закрытыми проводящими пучками, обычно параллельному жилкованию листьев, трехчленным цветкам и однобороздным пыльцевым зернам — однодольные образуют четко обособленные, издавна распознанные таксоны. Базальная линия однодольных представлена семейством Асогасеае с занесенным в Центральную Европу аиром (Acorns) и порядком Alismatales с такими растениями, как, например, сусак (Butomus), частуха (Alisma), рдест (Potamogetón), аронник (Arum) и ряска (Lemna). Преимущественно водный образ жизни организмов обеих линий часто давал основание предполагать, что все однодольные происходят от водных предков. Остальные однодольные более или менее ясно делятся на два больших блока. К «Lilianae» в центральноевропейской флоре, например, принадлежат лилия (Lilium), вороний глаз (Paris), орхидеи, спаржа (Asparagus), ландыш (Convallaria) и тамус (Tamus), а из тропических видов, например, панданусы (Pandanus). «Со- mmelinoidae», как вторая большая группа, представлена, например, семействами пальм (Агесасеае), злаков (Роасеае) и осоковых (Сурегасеае), а также бананами (Musa) и имбирем (Zingiber).
Остальные 74% покрытосеменных относятся к Rosidae (высшие, или настоящие двудольные, от англ. eudicots), которые отличаются от Magnoliidae и однодольных пыльцевыми зернами с тремя или более апертурами и, вероятно, имеют примерно тот же возраст, что и однодольные.
К базальным линиям Rosidae принадлежат такие различающиеся таксоны, как лотос (Nelumbo), п татан (Plot anus), барбарис (Berberis), лютик (Ranunculus) или мак (Papaver). В то время как в этих линиях еще можно наблюдать вариации в количестве и расположении элементов цветка, при формировании основной группы Rosidae («core eudicots») закрепились пятимерия цветка и двойной околоцветник, состоящий из чашечки и венчика. К базальной линии этой главной группы относят порядок Caryophyllales, который в центральноевропейской флоре представлен семейством гвоздичных (Caryophyllaceae) и к которому относятся также семейства кактусовых (Cactaceae) и аизооновых (Aizoaceae) с его «живыми камнями» — литопсами (Lithops). Большая часть Rosidae делится на два крупных блока: собственно, «Rosidae» и «Asteridae».
К собственно «Rosidae», характеризующимся большей частью свободнолепестным венчиком, двумя кругами тычинок, крассинуцеллятной семяпочкой с двумя интегументами и нуклеарным типом развития эндосперма, в Центральной Европе относятся, например, герань (Geranium), зверобой (Hypericum), молочай (Euphorbia), ива (Salix), фиалка ( Viola),клевер (Trifolium), крапива (Urtica), роза (Rosa), переступень (Bryonia), бук (Fagus), береза (Betula), дербенник (Lythrum), пастушья сумка (Capsella), липа (Tiliа) и клен (Acer). У «Asteridae» лепестки, как правило, сросшиеся, тычинки часто только в одном круге, семяпочка тенуинуцеллятная с одним интетументом, целлюлярный тип развития эндосперма. К ним в Центральной Европе относятся, например, дерен (Согnus), вереск (Calluna), первоцвет (Primula), горечавка (Gentiana), подмаренник (Galium), мята (Mentha), коровяк (Verbascum), подорожник (Plantago), красавка (Atropa), вьюнок (Convolvulus), незабудка (Myosotis), падуб (Ilех), тмин (Carum), ворсянка (Dipsacus), колокольчик (Campanula) и василек (Centaurea).
Хотя среднеевропейские представители «Rosidae» и «Asteridае» большей частью травянистые растения, в обеих больших группах Rosidae имеются и многочисленные древесные таксоны. Тем не менее травянистость среди «Rosidae» и «Аsteridae» встречается чаще, чем среди Мagnoliidae.
Краткий обзор эволюции покрытосеменных ясно показывает, что в их вегетативной сфере наблюдается отчетливая тенденция к травянистым жизненным формам. Эволюция цветка направлена к сокращению числа циклически расположенных органов, а в эволюции семяпочки происходили уменьшение и упрощение. Таким образом, у покрытосеменных продолжается та же тенденция, которую можно было наблюдать уже у разных голосеменных растений.