БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.3. Филогенез растений и история растительности
11.3.1. Обзор филогенеза организмов
Самые древние следы жизни можно обнаружить уже примерно через 1 млрд лет после возникновения Земли, т. е. примерно 3,5 млрд лет т. н. Атмосфера нашей планеты состояла тогда преимущественно из водяного пара, углекислого газа, азота (N2) и следов водорода (Н2), серы, хлора и фтора, метана, аммиака и т. п. Столь распространенный в настоящее время свободный кислород почти полностью отсутствовал, и озоновый слой, защищающий от жесткого космического и ультрафиолетового излучения, в первичной атмосфере еще не образовался. При возникновении жизни главную роль играли, очевидно, высокие температуры (например, вследствие вулканической деятельности) и электрические разряды.
Как многократно удалось показать экспериментально, при таких условиях могли возникать многочисленные органические соединения. К ним относятся аминокислоты, нуклеотиды, сахара, жирные кислоты и спирты или их предшественники — важнейшие «кирпичики» жизни. До сих пор не ясно, в каких точно условиях возникли эти молекулы и соответственно их полимеры: был ли это их водный раствор («первобытный суп»), или электрически заряженные поверхности («первобытная пицца») как, к примеру, пирит. Решающим тогда стало возникновение системы, способной к идентичной редупликации и кодированию. Предпосылкой для этого, очевидно, были полинуклеотиды типа РНК с автокаталитической активностью, способные поэтому катализировать собственную репликацию. Две или более подобные системы РНК могли включиться в так называемый «гиперцикл», в котором компоненты способствуют взаимной репликации. Благодаря связи такой кодирующей системы с липидными мембранами («компартментализация») и протеинами смогли, наконец, возникнуть какие-то пока еще гипотетические прогеноты. Приходится постулировать, что в них произошли постепенное усовершенствование генетической информации путем передачи этой функции от информационной РНК к кодирующей системе ДНК (сначала основанной на двух, затем — на трех нуклеотидах) с транскрипцией и трансляцией, а также оптимизация системы обмена веществ на основе все более многочисленных и лучше интегрированных управляемых ферментами процессов.
Самыми древними достоверными ископаемыми живыми существами были уже имевшие клеточную организацию прокариоты.
Они могли покрывать свои потребности в веществах и энергии либо за счет сложных органических соединений (первичные гетеротрофы, анаэробное брожение), либо за счет анаэробного хемосинтеза (например, используя S и Н2 при высоких температурах, см. 6.9). Сокращение легко доступных органических источников энергии, очевидно, сделало необходимым выработку окислительно-восстановительнои системы (цитохромы и т. п.) и затем постепенный переход к автотрофности. Сначала возникли, скорее всего, различные формы анаэробного фотосинтеза (см. 6.4.10) на основе циклического фотофосфорилирования, особенно с фотосистемой I и чаще всего с H2S в качестве донора электронов. Аэробный фотосинтез с использованием воды как источника электронов и восстановлением СО2 возник у цианобактерий путем эволюции фотосистемы II (см. 6.4.5). Это привело к увеличению содержания в атмосфере кислорода и создало предпосылки для возникновения значительно более экономичного обмена веществ, основанного на аэробном дыхании. Диверсифицирование органической жизни создало возможность эволюции самых разных форм вторичных гетеротрофов, таких как, например, сапрофиты и паразиты в разных группах прокариотических организмов.
Сегодня некоторые ставят под сомнение пока еще общепринятое деление прокариотических организмов на эубактерий и архебактерий (Bacteria и Archaea). Возможно, что первые гетеротрофные эукариотические клетки с клеточным ядром и хромосомами, эндоплазматической сетью и другими внутриклеточными мембранными системами, микротрубочками и актиновыми филаментами, иногда со жгутиками (типа 9 + 2) и, вероятно, также с митозом и мейозом происходят от общего предка с архебактериями. Впрочем, вполне допустимо, что первые эукариотические клетки возникли путем клеточного симбиоза архебактерий и эубактерий (гидрогенная гипотеза, см. 2.4.2). Если возникновение многочисленных внутриклеточных мембранных систем эукариотических клеток можно объяснить вероятным впячиванием (инвагинацией) клеточной мембраны прокариотического предка, то возникновение хромосом и клеточного ядра остается непонятным. Также не известно точно, когда определенные бактерии (Negibacteriota) попали путем первичного эндоцитоза в формирующуюся эукариотную клетку, где они стали митохондриями. Возможно, что это произошло перед или сразу после возникновения первых эукариотических организмов (Агсhаеzоа). Последнее часто принимают более вероятным, так как известны очень примитивные эукариоты без митохондрий. Сейчас, правда, считают, что эти организмы утратили свои митохондрии вторично, приспосабливаясь, например, к эндопаразитическому образу жизни, а, следовательно, митохондрии или соответственно гидрогеносомы принадлежат к базовому компоненту всех эукариот (см. 2.4.2) .
У эукариот количество ДНК увеличено, и она организована с гистонами в нуклеосомы, хромосомы и клеточные ядра. Распределение и дальнейшая передача наследственного материала стали более точными благодаря митозу, а возможности генетических рекомбинаций значительно возросли вследствие эволюции полового процесса с сингамией и мейозом.
Существует невероятное множество одноклеточных гетеротрофных эукариот с митохондриями. Различные представители такого организационного уровня стали исходным пунктом для дальнейшей эволюции. Вероятно, однократный симбиоз одной из этих линий развития с фотоавтотрофными бактериями (Суanobacteriotа) в ходе второго первичного эндоцитоза (см. 2.4.2) привел к возникновению первых растительных клеток, в которых эндоцитобионты стали пластидами. От этих первых ра
стительных клеток происходят все растительные организмы с простыми пластидами с двойной мембраной. Эти растения предстают перед нами тремя линиями: Glaucobionta, Rhodobionta (красные водоросли) и Chlorobionta (включая зеленые водоросли), из которых Rhodobionta и Chlorobionta более близки друг к другу. В группе Chlorobionta зеленые водоросли (Chlorophyta) являются сестринской группой Stre- ptophyta, которые состоят преимущественно из первично наземных растений (Embryophyta).
Все остальные группы водорослей, имеющих пластиды с 3 или 4 мембранами, возникали многократно путем вторичного эндоцитобиоза, когда различные гетеротрофные эукариотические одноклеточные включали разные водоросли. Зеленые водоросли являются вторичными эндоцитобионтами Chlorarachniophyta и Euglenophyta, а красные водоросли — Haptophyta, Cryptophyta, Dinophyta и Heterokontophyta.
Не совсем ясны отношения Dinophyta. Хотя вполне вероятно, что у них вторичными эндоцитобионтами являются красные водоросли, но в этой группе известны случаи, по-видимому, третичного эндоцитобиоза после утраты вторичных эндоцитобионтов.
В дальнейшей эволюции разных полифилетически возникших групп водорослей неоднократно параллельно шла эволюция от голых монадных или амебоидных форм к преимущественно неподвижным, имеющим слизистые чехлы или клеточные стенки кокковидным формам, затем к нитчатым и многоклеточным трихальным и, наконец, к ветвящимся в двух или трех измерениях, сложным талломным плектенхимным или паренхимным формам. Помимо этого, среди водорослей есть и сифональные формы. Это развитие коррелирует с переходом от свободного к сидячему образу жизни и увеличением размеров. Одновременно формировалась смена поколений, происходил переход от изогамии к анизогамии и оогамии и часто к преобладанию диплофазы над гаплофазой.
Организмы, известные как грибы, имеют разное происхождение и представляют четыре линии развития одноклеточных гетеротрофных эукариот. Эти линии привели к возникновению Acrasiobionta (клеточные слизевики), Myxobionta (слизевики), Oomycota (целлюлозосодержащие грибы) и Mycobionta (хитинсодержащие грибы). Предок Oomycota был также и предком выше названных Heterokontophyta, a Mycobionta, — пожалуй, ближайшие родственники многоклеточных животных.
У Oomycota и Mycobionta, как и у разных групп водорослей, можно наблюдать развитие от одноклеточных форм к сифональным нитчатым и сложным плектенхимным талломам. Заселение грибами суши и связанная с этим дальнейшая их эволюция, к примеру появление плодовых тел, тесно связаны с возникновением наземных растений.
Наконец, благодаря эктосимбиозу различных микобионтов с цианобатериями или зелеными водорослями многократно и независимо возникали разные лишайники (Lichenes).
Появившиеся в ордовике около 450 млн лет назад наземные растения (Embryophyta: Bryophytina, Pteridophytina, Spermatophytina1) находятся в тесном родстве с водорослями Streptophytina и вместе с ними составляют отдел Streptophyta, который характеризуется, к примеру, латерально расположенными (если имеются) жгутиками, клеточным делением обычно с образованием фрагмопласта, а также особой структурой целлюлозы. К Streptophytina относят, например, Charophyceae (харовые водоросли) с родом Chara и Coleochaetophyceae с родом Coleochaele, которых как близких родственников Embryophyta следует поместить в один подотдел. Streptophyta вместе с зелеными водорослями (Chlorophyta) составляют группу хлоробионтов (Chlorobionta).
1 Остатки семенных растений достоверно известны значительно позже — с верхнего девона. — Примеч. ред.
Наземные растения отличаются прежде всего тем, что их гаметангии (антеридии и архегонии) имеют оболочку из стерильных клеток и спорофит питается вначале как эмбрион за счет гаметофита.
Эволюция наземных растений проходила, вероятно, по мелководьям пресноводных водоемов с сезонными колебаниями уровня воды. В пользу этого свидетельствуют наличие и ныне в пресных водах близких родственников наземных растений, а также умозаключения о том, что условия здесь для возникновения наземных ра
стений были значительно благоприятнее, чем на морских побережьях. Важными предпосылками для заселения суши были возникновение кутикулы, устьиц, проводящих и механических тканей, межклетников и органов поглощения воды. Почти все эти структуры могли возникать как приспособление к лимитированной на суше доступности воды.
Первоначальной группой наземных растений можно считать мохообразные (Вгуорhуtinа), у которых более сложным анатомически и морфологически поколением является гаметофит, а неразветвленный спорофит всегда остается связанным с гаметофитом, быстро становится гетеротрофным и питается за счет гаметофита. Самая древняя подгруппа Вгуophytina, пожалуй, — талломные печеночники (Мarchantiopsida).
Папоротникообразные (Рterchantiopsida) и семенные растения (Spermatophytina) имеют морфолого-анатомически сложный, разветвленный спорофит, дифференцированный на основные органы: корень, стебель и лист1, а также ксилему и флоэму. Соответственно можно объединить папоротникообразные и семенные растения в группу побеговых, или сосудистых, растений (Коrmophyta, или Тracheophytа). Всегда талломное гаметофитное поколение у них постепенно редуцируется.
Пока недостаточно ясно, возникли ли Pteridophytina из в таком случае парафилетических по отношению к ним Bryophytina, поскольку они близкородственны антоцеротовым (Аnthocerotopsida), или же наземные растения сразу развивались двумя линиями — Вryophytina и Pteridophytina.
1 Такая деффиренциация тела свойственна не всем Pheridophytina. — Примеч.ред.
Pteridophytina очень давно разделились на две ветви — плаунообразные (Lycopodiopsidа), с одной стороны, и хвощеобразные (Equisetopsida), псилофитообразные (Psilotopsida) и папоротникообразные (Рteridopsidaа) — с другой; три последние группы являются сестринскими по отношению к Spermatophytina. Поэтому Рteridophytina по отношению к Spermatophytina парафилетичны и, соответственно, отличаются от семенных растений только отсутствием семян.
В ныне почти исключительно травянистой группе папоротников, значительно различающихся характером облиствения, структурой спорофиллов, а также анатомией стеблей, в прошлом не раз возникали древовидные формы. Точно также многократно параллельно происходит переход от равноспоровости с чаще всего обоеполыми гаметофитами к разноспоровости с мужскими гаметофитами, образующимися из микроспор, и женскими гаметофитами, образующимися из мегаспор.
Возникшие, по-видимому, в самом конце девона, примерно 370 млн лет тому назад, разноспоровые семенные растения отличаются тем, что женский гаметофит (зародышевый мешок) остается на спорофите в невскрывающемся мегаспорангии (нуцеллюсе) со стерильной оболочкой (интегументном), по крайней мере, до опыления. Эта структура из интегумента, нуцеллюса и зародышевого мешка, называемая семязачатком, после оплодотворения развивается в семя с семенной кожурой, питательной тканью эндоспермом и многоклеточным зародышем.
Возникновение семязачатка позволило преодолеть зависимость оплодотворения яйцеклеток в архегониях, развивавшихся на самостоятельном женском гаметофите, от капельножидкой воды.
Среди семенных растений, по-видимому, монофилетической группе из хвойных (Сniferopsida), саговниковых (Суcadopsida), гинкговых (Ginkgopsida) и гнетовых (Gnetopsida) (голосеменные) можно противопоставить цветковые растения (Мagnoliopsida) (покрытосеменные). Таким образом, приходится пересмотреть недавно еще общепринятое представление, что покрытосеменные наиболее близки к гнетовым и, следовательно, голосеменные парафилетич- ны относительно покрытосеменых. Родственные отношения четырех групп голосеменных неясны.
Самая богатая видами (около 250000) современная группа растений — покрытосеменные — в ископаемом состоянии впервые появляется в раннем мелу, примерно 140 млн лет назад. Она демонстрирует удивительное многообразие в экологии, в образовании травянистых и древесных жизненных форм, в биологии опыления и распространения, а также в обилии вторичных метаболитов. Поэтому сегодня покрытосеменные — доминирующий элемент почти всех типов растительности, они смогли заселить также различные экстремальные местообитания.
Вероятные родственные связи между различными группами растений и грибов обобщенно представлены на рис. 11.9.
11.3.2. История растительности
Как ясно показал обзор филогенеза организмов, флора и соответственно растительность Земли постоянно изменялись с момента возникновения жизни, вероятно, более 3,5 млрд лет назад и уж точно — с возникновения эукариотических автотрофных растений около 1,4 млрд лет назад. Однако вымирание организмов не было непрерывным (бокс 11.11, с. 517). Современный растительный покров Земли можно понять только как результат длительного исторического развития. Реконструкцией этого процесса занимаются исторические флористика и фитоценология. Изменение флоры и растительности Земли обусловлено эволюцией растений, изменением земной поверхности (расположения материков и океанов, горообразования), например благодаря тектонике плит; изменением общего состава атмосферы (концентрации кислорода и углекислого газа, появлении озонового слоя, поглощающего ультрафиолетовое излучение, изменении температуры, количества осадков и т. д.), например благодаря возникновению фотосинтеза и аэробного дыхания; изменением всех взаимодействий растений с другими, также изменяющимися организмами. Все эти различные факторы тесно связаны друг с другом.
Для реконструкции истории флор и растительности привлекают данные палеонтологии (соответственно палеоботаники), геологии, филогенетики и систематики и исторической биогеографии.
11.3.2.1. Методы
Фоссилизация растительных остатков происходит лишь в очень специфичных условиях, прежде всего в морских и озерных осадках, в торфе и образующемся из него со временем каменном угле. Лучше всего в ископаемом состоянии сохранились некоторые группы водорослей, а также фрагменты побегов, листья, споры, пыльца, семена и плоды сосудистых растений. Ископаемые растения относят, насколько возможно, к современным таксонам или описывают как вымершие таксоны. Для определения систематического положения ископаемых чаще всего можно использовать только морфологические, а в случае окаменелое тей — и анатомические признаки.
Бокс 11.11. Массовые вымирания
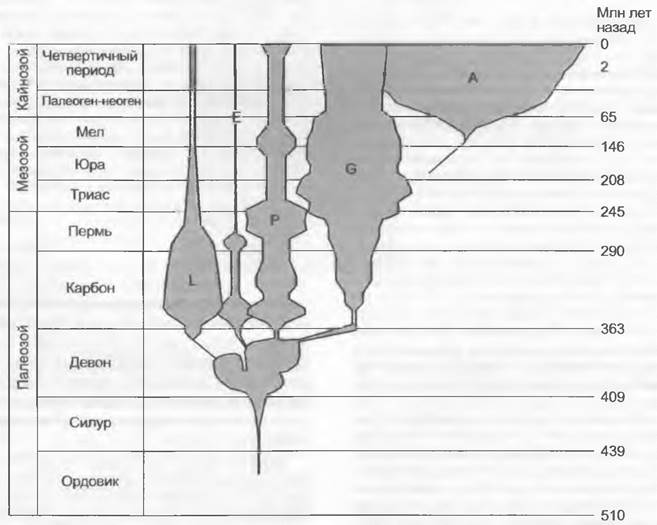
В истории Земли флора и растительность постоянно изменялись, одни растительные группы вымирали, появлялись новые (рис. А). Вымирание организмов, впрочем, не было непрерывным процессом. Возможно обнаружить периоды повышенной скорости вымирания, так называемые массовые вымирания. Очевидно, что массовые вымирания сосудистых растений, наземных позвоночных и морских беспозвоночных лишь в малой мере происходили одновременно. Некоторые авторы признают девять массовых вымираний сосудистых растений (девон — 391, 378 и 363 млн лет; карбон — 290 млн лет; триас — 241 млн лет; юра — 152 —155 млн лет; мел — 132 млн лет; третичный период - 29 и 16 млн лет) Возможно, самое известное (но не совпадающее с вышеназванными 9 периодами) благодаря вымиранию динозавров массовое вымирание на границе мела и третичного периода, как полагают, произошло из-за столкновения с Землей астероида. Этот удар привел к выбросу в атмосферу огромного количества пыли. В результате на долгие годы сократилось количество достигающей Земли солнечной радиации, и соответственно произошло заметное похолодание климата. Недавно на континентальном шельфе у побережья Юкатана (Мексика) был найден кратер, появившийся в результате этого удара, первоначально предполагаемого на основании повсеместно распространенного слоя, обогащенного иридием (иридий редок на Земле, но обычен в астероидах и кометах). Предполагают, что в различных областях Земли при этой катастрофе должно было вымереть от 20 до 75% сосудистых растений (по крайней мере, локально). И хотя эта внеземная причина массового вымирания на границе мела и третичного периода вполне вероятна, вряд ли таким путем можно объяснить все массовые вымирания в истории Земли.
Рис. А. Относительное многообразие групп наземных растений с начала ордовика: L — Lycopodiopsida; Е — Equisetopsida; Р — Pteridopsida; G — голосеменные; А — покрытосеменные
Непосредственно в ископаемом состоянии обычно сохраняются только скелетные элементы некоторых групп водорослей: кремнеземные диатомовые (Diatomеае), известковые Соccolithophorales (см. рис. 11.71), Соrallinасеае, Dasycladасеае. В других случаях ископаемые возникают путем окаменения, углефикации или в виде отпечатков и соответственно отливок.
При окаменении (см. рис. 11.130; 11.131;1. 139) органический материал клеточных стенок и клеточного содержимого заменяется минеральными веществами (например, кремнеземом, карбонатом кальция), и возникают ископаемые, в значительной мере передающие прижизненную структуру. Углефикация происходит в ходе потери газов и воды при механическом сдавлении, вследствие чего возникают сильно деформированные остатки. Отпечатки и отливки сохраняют только форму поверхности соответствующей части растения. Заключенные в янтаре ископаемые являются чаще всего отпечатками или отливками полостей, но могут содержать также и органический растительный материал. Важный метод анализа ископаемых — получение тонких шлифов (см. рис. 11.142), с которых послойно снимают отпечатки на пластиковой пленке. На некоторых образцах хорошо различимы анатомические или даже электронно-микроскопические структуры (например, строение гранхлоропластов в листьях из миоцена).
Обычная проблема при реконструкции ископаемых находок — определить принадлежность отдельно найденных и описанных органов. Так, например, к всеобщему изумлению в 1960 г. было показано, что ранее описанные как Саllixylon стволы, анатомически похожие на древесину голосеменных, и папоротникообразные листья рода Archaeopteris (см. 11.2, рис. II.288) на самом деле принадлежали одному растению.
Особое значение, в первую очередь для более молодых отложений, имеет прекрасно сохраняющиеся благодаря устойчивой к всякого рода воздействиям экзине споры и пыльцевые зерна сосудистых растений. Анализом спор и пыльцы занимается палинология (см. 11.2 — Микроспорофиллы).
Споры и пыльцевые зерна можно легко классифицировать на основе их глубокой структурной дифференцировки (см. 11.2 — Пыльца). Прежде всего, споры и пыльца ветроопыляемых, а также некоторых насекомоопыляемых растений рассеиваются в огромном количестве. В Центральной Европе ежегодно на квадратный сантиметр почвы оседает несколько тысяч пыльцевых зерен и спор, которые захораниваются в отложениях (к примеру, в морском известняке, торфе, грубогумусной почве и т. п.). По первому появлению споровых тетрад можно определить время возникновения мейоза и, соответственно, полового процесса, а самые древние трикольпатные пыльцевые зерна (см. рис. 11.268) позволяют датировать появление «Rosidae». Для изучения развития флоры и растительности в четвертичном периоде можно из подходящих отложений получить буровую колонку, послойно обработать и количественно проанализировать ее. Графическое представление пыльцевой диаграммы (см. рис. 11.306) показывает наличие и меняющееся соотношение спор и пыльцевых зерен разных видов в охваченном буровым профилем временном срезе. При знании современных количественных показателей осаждения пыльцы в различных типах растительности возможна даже реконструкция изменения количественного состава растительности вблизи от места бурения.
Знание возраста ископаемых растительных остатков имеет центральное значение для истории флоры и растительности. На смену сравнительной хронологии истории Земли (рис. 11.293), которая опирается на наличие в исследуемых слоях определенных животных и растительных руководящих остатков, сегодня пришли различные методы определения абсолютного возраста.
Рис. 11.293. Хронология истории Земли
Возраст горных пород определяют с помощью радиометрических методов. При этом используют постоянство периода полураспада радиоактивных элементов в минералах. Например, период полураспада урана (236U —> 206Рb) составляет 4,5 млрд лет. По относительной концентрации радиоактивного элемента и продукта его рас
пада можно определить возраст горной породы. Для датировки ископаемых остатков особое значение имеет радиоактивный калий (40К —> 40Са или 40Аг) как важная составная часть большинства осадочных пород. При использовании этого элемента хорошо удается определять возраст ископаемых находок более 100 000 лет. Для определения более молодых ископаемых (< 50 000 лет) чаще всего используют радиокарбоновый метод. Он основан на том, что в органических соединениях первоначальное отношение изотопов углерода 12С:14С, равное таковому в воздухе, благодаря распаду 14С в 14N (период полураспада 5 730 ± 40 лет) сдвигается в сторону 12С. Другие методы абсолютной датировки ископаемых еще более молодого возраста базируются на различных процессах с выраженным годовым ритмом. Это — определение возраста по годовым кольцам прироста древесины у деревьев (дендрохронология), посредством чего иногда удается точно определить возраст до 8 000 лет. Так называемые ленточные глины и подобные им отложения, равно как и ледники, показывают слоистость, которую можно использовать для определения абсолютного возраста. Далеко разносящийся при извержениях вулканов пепел также может стать важной точкой отсчета для определения возраста. Относительно грубые палеомагнитные датировки основаны на том, что магнитное поле Земли со временем периодически меняет полюса, причем ориентация магнитных полюсов «фоссилизируется» в железосодержащих горных породах. Правда, временные границы отрезков с различной полярностью приходится устанавливать изначально другими методами. На протяжении третичного и четвертичного периодов кайнозоя эпохи противоположной полярности имели длительность от 20 тыс. до 730 тыс. лет.
Для последнего тысячелетия можно установить возраст растительных остатков, если вместе с ними были найдены предметы до или ранне исторической культуры известного возраста. Если таким образом установлена одновременность некоего доисторического периода и определенного «лесного времени», то можно, наоборот, определить возраст археологических находок неизвестного возраста по пыльцевому анализу, если в раскопе сохранилась подходящая ископаемая пыльца.
Не только ископаемые находки, но и родственные связи, и распространение современных таксонов позволяют сделать косвенные выводы об истории флоры и растительности определенной области. Многие таксоны имеют дизъюнктивные ареалы, промежутки между частями, которых обычно очень велики и естественным образом в ходе обычного расселения непреодолимы. К примеру, существует дизъюнкция ареалов многих групп растений между Восточной Азией и восточной частью Северной Америки (см. рис. 14.24). Если исключить дальнее расселение в качестве объяснения такого примера ареалов, то понять его можно только при допущении существования в прошлом сплошной области распространения таксона. Это подтверждают данные о климате и растительности прошлого в областях, расположенных между Восточной Азией и востоком Северной Америки. В настоящее время многочисленные исследования родства, основанные на последовательностях ДНК, позволяют в известной мере проводить абсолютное датирование, опираясь на концепцию молекулярных часов. Если удается датировать филогенетическое расхождение тех же восточноазиатских и североамериканских форм, то, исключив дальнее расселение, возможно утверждать, что по крайней мере до этого времени должна была
существовать единая общая область распространения.
Так как молекулярные часы приходится калибровать по ископаемым остаткам или геологическим событиям, существует опасность попасть в порочный «круг доказательств».
Успешные реконструкции истории флор и растительности Земли требуют географически и систематически репрезентативного и легко интерпретируемого материала. Добротную картину истории флоры и растительности, соответственно, получают главным образом для молодых геологических отложений и для областей с давними научными традициями (например, Европа, Северная Америка и во все большей степени Китай) и в основном для таких организмов, которые имеют структуры, хорошо сохраняющиеся при фоссилизации.
11.3.2.2. Докембрий и палеозой (4 млрд — 245 млн лет)
Со времени возникновения жизни более 3,5 млрд лет назад и до заселения суши многоклеточными растениями, вероятно, в ордовике (около 450 млн лет назад) теплые моря, по-видимому, населяли одноклеточные и многоклеточные организмы, сильно различавшиеся организацией, образом жизни и формами получения энергии.
В этот промежуток времени возникли эубактерии и архебактерии как прокариотические организмы, гетеротрофные эукариоты, вначале без митохондрий, а затем с приобретенными путем эндоцитобиоза митохондриями и, наконец, фотоавтотрофные эукариоты — благодаря эндоцитобиозу между гетеротрофными эукариотами и фотоавтотрофными цианобактериями.
Несомненные ископаемые клетки прокариот, напоминающие различных бактерий, известны из формаций Fig-Tree Южной Африки возрастом 3,1 млрд лет. Организмы, по образу жизни, вероятно, напоминавшие архебактерий, недавно были найдены в окаменелых морских глубоководных вулканических породах возрастом около 3,2 млрд лет. В слоях возрастом 3 — 1,6 млрд лет из Африки и Северной Америки многообразие прокариотических форм увеличивается. Так, находят, к примеру, похожие на Oscillatoria цианобактерии, образовавшие ископаемые строматолиты. Первые эукариотические водоросли появились ранее 1,4 млрд лет назад, а в древних формациях Австралии возрастом от 1,5 до 0,9 млрд лет найдены на удивление хорошо сохранившиеся остатки одноклеточных водорослей (Caryosphaeroides, напоминающие СЫогосос- cales), у которых можно распознать разные стадии деления клетки и даже остатки клеточного ядра. Эти биоценозы содержали также бактерии, водные грибы и одноклеточных животных (Protozoa). Ранее ордовика дифференцировались две главные линии ранней эволюции растений, приведшие к зеленым и красным водорослям.
Изначальное отсутствие кислорода в атмосфере требовало получения энергии путем самых разных форм анаэробной гетеро- и автотрофии. Ранние прокариоты вели, вероятно, образ жизни, отчасти сходный с таковым современных крайне термофильных видов архебактерий. Аноксигенный и оксигенный фотосинтез появился, вероятно, уже более 3 млрд лет назад. С тех пор концентрация свободного кислорода в атмосфере постепенно нарастала. К периоду 2,8 — 2,4 млрд лет назад концентрация атмосферного кислорода достигла примерно 1 — 2 % от современной концентрации (21 %). При таких условиях сократилось число доступных для анаэробных организмов местообитаний и началась дифференциация аэробных организмов и эволюция более эффективного дыхания. Ко времени возникновения эукариотических водорослей концентрация кислорода в атмосфере достигла примерно 10%. С увеличением концентрации кислорода в атмосфере связано и возникновение задерживающего ультрафиолетовое излучение озонового слоя, ставшего важной предпосылкой заселения суши.
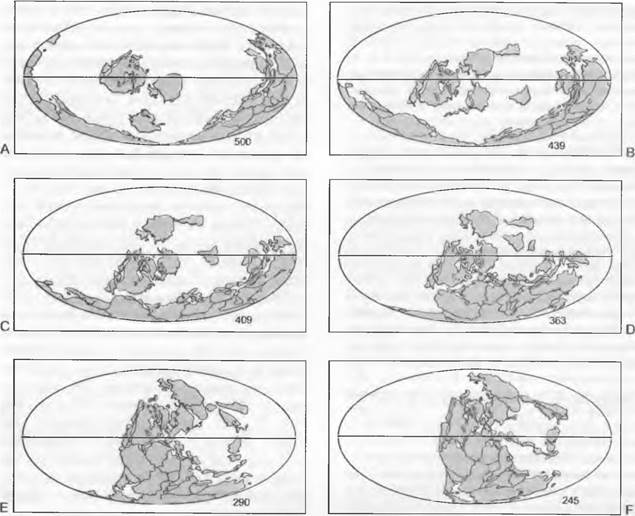
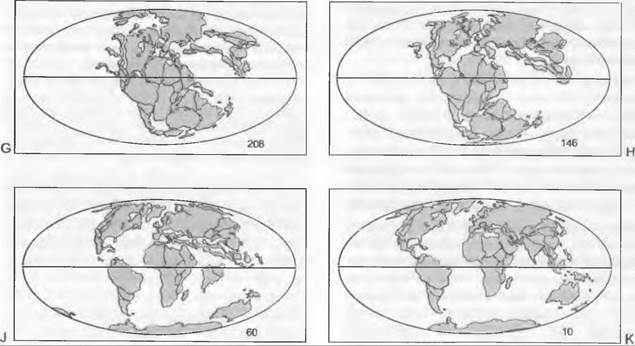
Ко времени заселения суши (приблизительно 450 млн лет назад) в ордовике (510 — 439 млн лет; здесь и ниже цифры означают число лет до нашей эры) центры современных материков располагались преимущественно в Южном полушарии (рис. 11.294), и климат Земли был теплым. Из этого времени происходят таксономически не определимые первые ископаемые находки до некоторой степени засухоустойчивых растений, представленные спорами, фрагментами кутикулы с отпечатками клеток и трубковидными клеточными элементами с поясковыми утолщениями стенок. В силуре (439 — 409 млн лет) встречаются остатки растений из вымершей группы Homeophytae (например, Caía, Tortilicaulis), а также род Cooksonia из линии развития Lycopodiopsida. В девоне (409 — 363 млн лет) с обильными осадками и наводнениями, но также и с более выраженной засушливостью климата континенты занимали явно более северное положение (рис. 11.294). В этот период выявлена сильная дифференциация наземных растений, из-за чего девон рассматривают как начало палеофита, сменившего протерофит. Один из известных представителей девонских растений — род Rhynia (396 млн лет; см. рис. 11.130), относящийся к ныне вымершей линии развития. Почти все ныне существующие эволюционные линии растений впервые встречаются в девоне. Из Lycopodiopsida это, к примеру, Drepanophycus (примерно 410 млн лет) и Zosterophyllum (396 млн лет; см. рис. 11.130). Разделение Lycopodiales на Isoetales и Lycopodiales выявлено в верхнем девоне. Линия развития, ведущая к папоротникам, хвощам и семенным растениям, представлена, например, Psilophyton (390 млн лет; рис. 11.288) и Tetraxylopteris (381 млн лет; рис. 11.288). Ранний представитель хвощей— Ibyca (381 млн лет), а Rhacophyton (377 млн лет) считают представителем папоротников. Семенные растения встречаются также уже в верхнем девоне — Elkinsia (367 млн лет).
Ожидаемых различных ископаемых представителей Bryophyta, этой первоначальной группы ныне живущих наземных растений, в девоне очень мало, вероятно, из-за их очень плохой сохранности в ископаемом состоянии. Только Pallavicinites (370 млн лет) из Jungermanniopsida достоверно известен из девона. Впрочем, предполагали, что некоторые ископаемые из ордовика следует относить к печеночникам. Достоверно ископаемые антоцеротовые и лиственные мхи впервые встречаются в мелу.
С изменением систематического состава флоры в силуре и девоне началось и изменение растительности. Самые ранние наземные сообщества были разреженными, низкими (<50 см) и чаще всего представляли, по-видимому, земноводные заросли по берегам водоемов или в заболоченных понижениях рельефа. Позднее встречались уже и более высокие и полностью наземные сообщества. Эти наземные биоценозы в силуре и девоне были, очевидно, распространены по всему свету (рис. 11.294; 11 295).
Рис. 11.294. Расположение континентов с начала ордовика (числа — миллионы лет)
Наличие редуцентов (бактерий и грибов) на растениях показывает, что первые наземные экосистемы функционировали в сущности так же, как и современные. Вероятно, что уже самые ранние наземные растения имели везикулярно-арбускулярную микоризу (см. 9.2.3: Микориза). Вполне возможно, что этот симбиоз сыграл решающую роль в заселении суши.
В карбоне (363 — 290 млн лет) материки еще дальше сдвинулись на север (см. рис. 11.294).
В этом периоде появились первые обширные леса с деревьями высотой более 30 м. Из остатков этих лесов образовался каменный уголь.
Центральная область этих лесов охватывала Европу и восточную часть Северной Америки, они известны также в Сибири и Восточной Азии. Во всех этих областях господствовал влажный теплый (суб-) тропический климат (рис. 11.296). Растения каменноугольных лесов не имели годовых колец древесины или покоящихся почек, анатомия их листьев подобна анатомии листьев растений современных дождевых тропических лесов.
Состав атмосферы почти достиг современных показателей. В таких условиях на сырых и переувлажненных торфяных почвах росли мощные болотные леса (рис. 11.295), в которых доминировали древовидные хвощи и плауны, а также кордаиты. Эти биоценозы были многовидовыми и сильно дифференцированными на ярусы и зоны.
Зональность определяла доступность воды. Заросли хвощей (Archaeocalamites, Calamites; см. рис. 11.142; 11.143), очевидно, занимали мелководья. На незалитых местах обитали древовидные плаунообразные высотой до 40 м и с диаметром стволов в 2 м (Lepidodendron, Sigillaria', см. рис. 11.138; 11.139) с богатой сопровождающей флорой из менее высоких древовидных папоротников, кустарниковых или лиановидных семенных растений из ныне вымерших таксонов и с нижним ярусом из травянистых клинолистов (см. рис. 11.142), лиственных мхов и печеночников. Более сухие местообитания бьли населены, очевидно, кордаитами (см. рис. 11.206). Благодаря мошной вторичной древесине они, вероятно, лучше поддерживали водный баланс, чем имевшие толстую кору и мало древесины лепидодендроновые.
Рис. 11.295. Реконструкция каменноугольного леса (Музей естественной истории, Чикаго). Слева вверху ветвь с листьями и колосками из спорофиллов лепидодендрона (Lepidodendron), правее стволы лепидодендрона и сигиллярии (Sigillaria), между ними вайя Neuropterís с образующимися семенами, а также тонкие побеги Lyginopteris (оба — Pteridospermae); в центре на переднем плане клинолист Sphenophyllum, сзади папоротник с гигантской древней стрекозой, а также другие древовидные плауновидные, справа каламит (Calamites)

Рис. 11.296. Glossoptens, цельный лист с сетчатым жилкованием (1/Зх)

В каменноугольных лесах встречались и животные, например, амфибии, первые рептилии, пауки, многоножки и примитивные насекомые (стрекозы, тараканы). Можно обнаружить здесь и паразитические, симбиотические и сапрофитные грибы.
Одновременно с каменноугольными лесами современного Северного полушария в Южной Африке, Индии, Австралии, Антарктиде и на юге Южной Америки развилась совершенно другого типа, более скудная так называемая гондванская флора. Руководящими формами здесь считают кустарниковые семенные растения, например, род Glossopteris (рис. 11.296), разные папоротники и хвойные. Соответственно более прохладному умеренному климату этих регионов, составлявших южный континент Гондвану, древесина здешних растений иногда была с годовыми кольцами. Похолодание и аридизация климата в перми (290 — 245 млн лет) привели к исчезновению каменноугольных лесов и вымиранию древовидных плаунообразных и клинолистов. Зато можно наблюдать диверсификацию голосеменных, к примеру, Voltziales (см. 11.2; рис. 11.207). Это уже начало мезофита. В карбоне и еще сильнее в перми Южный континент подвергся оледенению.
11.3.2.3. Мезозой (245 — 65 млн лет)
По крайней мере с начала аридного или полуаридного триаса (245 — 208 млн лет) в течение всей достаточно влажной юры (208 — 146 млн лет) и до умеренного или теплого нижнего мела (146 — 65 млн лет) флоры по всей Земле были однообразны вследствие однообразия климата, без каких- либо следов оледенений.
В этот огромный промежуток времени продолжалось движение континентов, и в мелу возникло море Тетис, разделявшее Северный и Южный материки (см. рис. 11.294).
Главную роль во флорах в это время играли папоротники, хвощи и особенно разные группы голосеменных вроде Ginkgoopsida, Coniferopsida и Cycadopsida, а также вымершие беннеттиты (см. рис. 11.212).
Обилие видов сосудистых растений и их биотопическое разнообразие сильно возросли по сравнению с карбоном, что можно объяснить экспансией растений в более сухие местообитания и коэволюцией с наземными группами животных (например, опыление насекомыми беннеттитов, распространение семян рептилиями).
Мезофит северной континентальной области начался с наступившего в верхней перми (цехиггейна) и продолжавшегося до среднего триаса длительного аридного периода, который привел к формированию ксероморфных групп растений вроде Voltziales (см. рис. 11.207) и унификации флор Северного и Южного континентов. В более поздних флорах мезофита среди папоротников преобладали эвспорангиатные и первичные лептоспорангиатные. Ginkgoopsida были представлены примерно 11 разнообразными родами (например, Baiera, см. рис. 11.202), а представленный только одним современным видом род Ginkgo с нижней юры до верхнего мела распространен почти по всей Голарктике (см. рис. 14.16). Ареал рода Araucaria из хвойных был в юре космополитным, но с третичного периода ограничивался Южным полушарием и сегодня сократился до разрозненных фрагментов в Западной Пацифике, Австралии и Южной Америке. Среди хвойных в юре и нижнем мелу были предшественники современных Podocarpaceae Южного полушария, а также Taxaceae, Cephalotaxaceae и Cupressaceae, и, возможно, также Рinaceae. Особенно типичны для флор мезофита Сycadopsida с существующими и поныне и уже вымершими таксонами (рис. 11.297).
Рис. 11.297. Последовательность появления типов пыльцы и листьев древнейших покрытосеменных от нижнего мела (баррем, апт, альб) до начала верхнего мела (сеноман) на востоке Северной Америки (и в других местах). Типы пыльцы: А, В — дистально-однобороздная (А — Clavatipollenites, В — Liliacidites и др.); С — трехбороздная (Tricolpites и др.); D— F — трехборозднооровая (Tricolporoidites). Типы листьев: G — обратнояйцевидный, с нерегулярным жилкованием, сходный с листьями однодольных (Acaciaephyllum); Н — почковидный (Proteaephyllum); J — яйцевидный зубчатый, с перистым жилкованием (Quercophyllum)\ К— узкообратно-яйцевидный цельнокрайный (Rogersia); L— широкоэллиптический (Ficophyllum); М — пальчатолопастной (Vitiphyllum); N — обратнояйцевидный (Celastrophyllum); О — щитовидный (Nelumbites, Menispermites); Р — яйцевидно-сердцевидный (Populophyllum); Q — эллиптический, зубчатый (Celastrophyllum); R, Т — перистый (Sapindopsis); S, U — трехлопастной, возрастание регулярности жилкования (Araliaephyllum и Araliopsoides)
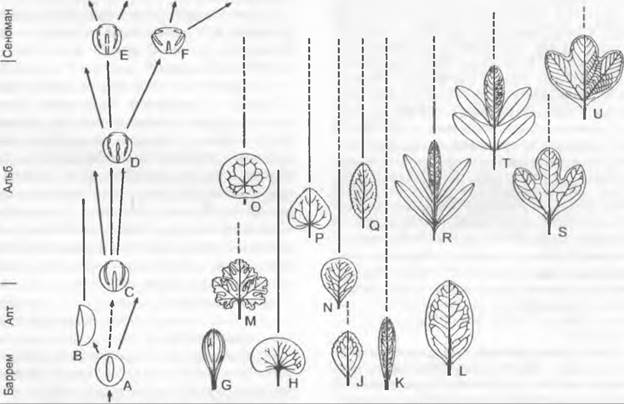
Важным этапом в истории флор и растительности Земли стало возникновение покрытосеменных (примерно 140 млн лет) в нижнем мелу. Покрытосеменные появились, вероятно, как травы и кустарники нарушенных местообитаний в низких географических широтах в полуаридных или аридных условиях. Вскоре после возникновения покрытосеменных произошло разделение Северного (Лавразии) и Южного (Гондваны) материков морем Тетис. Сравнительное единообразие мирового климата уступило место все более усиливающемуся градиенту температур от полюсов к экватору. Причину быстрой диверсификации и высокого видового разнообразия покрытосеменных следует искать не только во многих особенностях покрытосеменных, повышающих темпы их видообразования, но и в этих тектонических и климатических изменениях Земли. Уже в среднем мелу покрытосеменные стали доминирующей группой растений в большинстве типов растительности по всему миру. Это время отмечает начало неофита, ознаменовавшееся вымиранием или сильным сокращением таксонов, доминировавших в мезофите.
Более или менее непрерывные ряды ископаемых форм (рис. 11.297) показывают увеличение разнообразия пыльцы и листьев. Из этого можно сделать вывод о ранней диверсификации жизненных форм (от древесных до травянистых) и заселенных биотопов (от сухих до влажных).
Можно думать, что современные ареалы по крайней мере некоторых голарктических, пантропических или распространенных в Южном полушарии таксонов обусловлены положением континентов в мелу и раннем третичном периоде. Так, Северная Америка и Евразия сравнительно поздно отделились друг от друга в области Северной Атлантики; Африка, Аравия и Индия были долгое время связаны друг с другом, тогда как Африка и Южная Америка, напротив, рано разделились (примерно 90 млн лет), а сухопутная связь и возможность миграции между Южной Америкой, Антарктидой и Австралией с Новой Зеландией существовала еще примерно 40 млн лет назад. Пространственное сближение или соединение Индии с Южной Азией, Австралии и Новой Гвинеи с Юго-Восточной Азией, а также Северной и Южной Америк (примерно 3 млн лет) имело место лишь в самые поздние геологические эпохи. В следующих примерах звездочками отмечены достоверно известные уже из мела ископаемые остатки групп, демонстрирующие соответствующие типы распространения. Голарктическое: Magnoliaceae* (см.рис. 14.24), Сеrcidiphyllaceae*, Platanaceae*, Fagaceae* (см. рис. 14.17), Вetulaceae*, Раеоniасеае. Пантропическое: Аnnоnасеае, Моnimiасеае, Сhloranthасеае*, Myrtaceae*, Flacourtiасеае, Sapotaceae*, Rubiасеае, Агесасеае*. Африканско-южноамериканское: Vellozizceze. В Южном полушарии: Winteraceae*, Protеасеае*, Nothofagасеае*.
Эти и другие примеры возможного мелового или раннего третичного происхождения ряда ареалов требуют критической перепроверки в свете накапливающихся данных геносистематических работ. Использование молекулярных часов, например, в исследованиях Gentianасеае и Melastomataceae показало, что миграции между тропическими континентами, вероятно, имели место и спустя долгое время после разделения континентов либо путем заноса, либо путем миграции через сегодняшние области с умеренным климатом. Для пантропических Lauraceae удалось показать, что пантропический ареал некоторых групп родов можно объяснить дрейфом континентов, тогда как другие группы родов с подобным же распространением слишком молоды для этого.
11.3.2.4. Кайнозой (65 млн лет — по настоящее время)
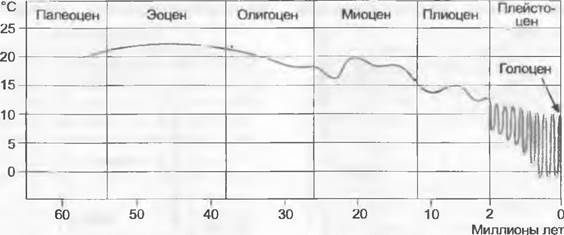
Палеоген-неогеновый период (65 — ок. 2 млн лет) — время очень глубоких климатических изменений. В то время как в первой половине периода, в палеогене (палеоцен 65 — 54 млн лет, эоцен 54 — 38 млн лет, олигоцен 38 — 26 млн лет), был широко распространен мягкий или даже субтропический климат, в более позднее время, в неогене (миоцен 26 — 12 млн лет, плиоцен 12 — около 2 млн лет; начало плиоцена иногда датируют 5,2 млн. лет), произошли похолодание и аридизация климата, которые продолжилось в оледенениях четвертичного периода, или плейстоцена.
Существенной причиной этих климатических изменений стало изменение поверхности Земли. Поднятие в миоцене и плиоцене Гималаев, Альп, Анд и т. п. привело к образованию обширных засушливых областей в «дождевой тени» этих новых горных систем, смещение огромных масс суши в более высокие географические широты способствовало образованию полярных ледяных «шапок» (рис. 11.294), а сокращение или высыхание больших морей вроде Тетиса привело к появлению континентального климата в удаленных от морей областях. Поэтому климат Земли в конце палеоген-неогена был значительно сложнее, чем в начале его. Эти климатические и геоморфологические измерения сильно повлияли на развитие флоры и растительности (рис. 11.298).
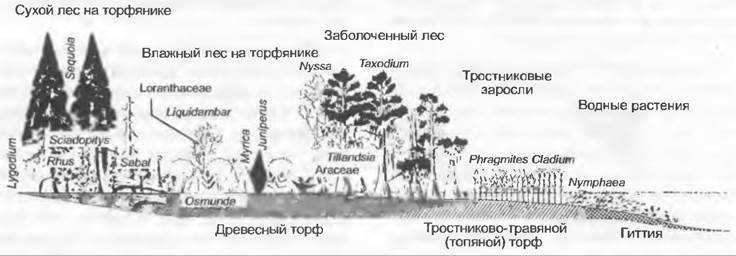
Рис. 11.298. Реконструкция поясов растительности среднетретичного болота Центральной Европы
В палеогене даже в областях современного умеренного климата были широко распространены тропическо-субтропические флоры вечнозеленых дождевых лесов с Lauraceae (например, Cinnamomurri), Moraceae (Artocarpus, Ficus), Juglandaceae (Engelhardtia), пальмами (Sabal. Elaeis, Nypa) и тропическими папоротниками (к примеру, Matonid). «Языки» этих флор достигали даже ныне арктических регионов, например, Аляски и Гренландии. В палеогене Голарктики значительно дальше к северу были распространены богатые видами листопадные лиственные и смешанные леса. Такие флоры встречались до Шпицбергена и Канадского архипелага (81°45' северной широты, в настоящее время среднегодовая температура там -20°С). Они состояли из видов Pinus, Picea, Platamis, Fagus, Quercus, Corylus, Betula, Ainus, Jugions, Ulmus, Acer, Vitis, Tilia, Populas, Salix, Fraxinus и др. родов, которые еще и сегодня встречаются, к примеру, в умеренном климате Европы. Другие роды в современной Европе уже давно вымерли, но встречаются в более теплых областях Северной Америки (Taxodium, Sequoia), Восточной Азии (Ginkgo, Cercidiphyllum) или в обеих областях (Tsuga, Magnolia, Liriodendron, Sassafras, Carya, Diospyros). Так как Северные материки тогда еще менее разошлись друг от друга, чем в настоящее время (см. рис. 11.294), в третичном периоде в циркумполярном регионе, вероятно, происходит более оживленный флористический обмен. В результате возникла так называемая аркто-третичная флора — ядро современной флоры Голарктиса.
Для Центральной Европы принимают среднегодовую температуру в эоцене 22 °С. В Северном полушарии граница леса была смещена к северу на 20 — 30°, а северная граница распространения пальм — на 10 — 15° по сравнению с современными. Ископаемые остатки этих палеогеновых флор найдены в Европе, например, в Экфельде — Эйфеле и Месселе под Дармштадтом, а в Гайзельтале — под Халле. Балтийская флора янтарей относится к этому же времени. В ней удалось найти дополнительно к вышеназванным родам представителей Согпасеае (Mastixia), Annonaceae, Theaceae (Stewartia), Stre- culiaceae, Sapotaceae, Symplocaceae, Pandanaceae
и Cyatheaceae, которые в настоящее время распространены главным образом в тропиках и являются реликтовыми таксонами Юго-Восточной Азии. Тогдашняя растительность, вероятно, была похожа на современные, богатые представителями Lauraceae горные леса Юго-Восточной Азии. Три европейских рода (Ramonda, Jankea. Haberlea) пантропических Gesnerlaceae являются, вероятно, реликтами этой тропической третичной флоры.
С эоцена до миоцена в Центральной Европе из органических отложений мелеющих озер и граничащих с ними заболоченных лесов формировались отложения бурого угля. Руководящими формами третичных лесов (рис. 11.298) были обитающие в настоящее время в теплоумеренном поясе Северной Америки реликтовые роды хвойных Taxodium и Séquoia, а также общий для Америки и Азии род Nyssa (Согnасеае).
В зарастающих водоемах (рис. 11.298) на глубине наряду с Nymphaea росли и другие нимфейные, например, Brasenia (современное распространение: Америка, Азия). Мелководье занимал пояс осок и тростника с Dulichium (Сурегасеае; Америка). Далее следовал пояс переувлажненного заболоченного леса с Nyssa, Taxodium и эпифитными Tillandsia(Bromeliaceae; Америка). Выше нормального положения водного зеркала росли болотистые леса. В более влажных местах встречались Myrica, Liquidambar (Америка, Азия), Cyrilla(Америка) и Osmunda claytoniana (Америка, Азия), а на более сухих — Séquoia (Америка), Sciadopitys (Азия). Sabal (Америка. Азия) и вьющийся пантропический папоротник Lygodium.Из названных родов сегодня в Европе еще растут только Nymphaea и Myrica. Характерные для торфяных лесов особенности в настоящее время сохранились в субтропической Юго-Восточной Америке (например, во Флориде).
Похолодание и частичная аридизация климата в неогене привели к смещению флор и растительных зон к югу Земли, вымиранию в Европе почти всех тропических и многих теплолюбивых арктотретичных таксонов и появлению обширных дизъюнкций в ареалах многих видов голарктических лиственных лесов в континентальных областях Азии и на западе Северной Америки.
С эоцена ло плиоцена в Северном полушарии уменьшилась доля ранее доминировавших тропических таксонов, и стали господствовать арктотретичные представители. В Европе серьезным препятствием для миграций третичных (а также и четвертичных) флор были высокие горы, протянувшиеся с запада на восток, и Средиземное море. Отсюда понятно, почему сегодня Европа значительно беднее аркготретичными видами, чем климатически сравнимые области Восточной Азии и Северной Америки.
Характерные дизъюнкции: Восточная Азия— Европа— восточная Северная Америка (Fagus — рис. 14.17; Carpinus, Hepatica) и Восточная Азия — восточная Северная Америка (Tsuga, Magnoliaceae — рис. 14.24, IHicium, Sassafras, Hamamelis, Nyssa, Catalpa) и соответственно реликтовый эндемизм юго-востока Северной Америки (Taxodum) или Восточной Азии (Ginkgo. Metasequoia, Cercidiphyllum) следует понимать как этапы продолжающегося сокращения ареалов арктотретичных таксонов. Среднегодовые температуры в Центральной Европе снизились от миоцена до нынешнего времени примерно с 16 °С до 8 — 9°С.
Особенно важные рефутиумы арктотретичных таксонов (третичных реликтов) в Европе и Юго-Западной Азии — Балканы (например, Picea omorica, как близкородственная восточноазиатской P. jezoensis, и Aesculus hippocastanum — Aesculus встречается еще в Гималаях, Восточной Азии, а также в Северной Америке), влажные биотопы восточного Средиземноморья (например, Platanus orientalis, этот род встречается только еще в Северной Америке, а в Средней и Восточной Азии вымер; Ulmaceae — Zelkova, есть еще в Восточной Азии, так же как Liquidambar и Styrax) и лесные области по восточному побережью Черного и южному побережью Каспийского морей (например, Plerocarya из Juglandaceae есть еще в Восточной Азии, а в Северной Америке вымер; Albizia (Mimosaceae) и Diospyros (Ebenaсеае) с субтропическими или даже тропическими связями).
Возникновение теплого континентального, с сухим летом климата в субмеридиональных и меридиональных зонах (например, в Средиземноморье, на западе Северной Америки, а также и в Южном полушарии, например, в Чили) привело в середине палеоген — неогена к преобразованию тогдашних вечнозеленых дождевых лесов в жестколистные леса. В Европе этот процесс ускорило многократное высыхание Средиземного моря в миоцене. Примеры этих изменений флор — появление Myrtis communis и Smilax aspera из преимущественно тропических семейств, а также Quercus ilex, Nerium oleander и Olea europaea.
Возникновение жестколистных лесов хорошо документировано, особенно в олигоиеновых и миоценовых ископаемых флорах Юго-Восточной Европы с предками современных видов Laurus, Arbutus, Ceratonia, Pistacia, Phyllyrea и т. п., а также на западе Северной Америки. Леса из лавровишни на Канарских островах можно рассматривать как по меньшей мере частичный реликт этой фазы. Для некоторых обитателей лавровишневых лесов (например, Ixanthus viscosas (Gentianaceae), ближайшего родича средиземноморской Blackstonia) с помощью молекулярных методов удалось показать более молодой возраст. Впоследствии в дальнейшей эволюции средиземноморской флоры особенно важную роль сыграли геологически стабильные области Иберийского полуострова (рис. 11.299) и северо-западной Африки, а также юго-западной Азии.
Рис. 11.299. Колебания климата в кайнозое. Среднегодовые температуры реконструированы для Западной и Центральной Европы. Временной масштаб для плейстоцена и голоцена растянут, а число холодных и теплых периодов меньше, чем их было на самом деле
С возрастанием сухости и континентальности удаленных от морей областей в неогене, очевидно, непосредственно связано возникновение и прогрессирующее развитие ксерофильных безлесных флор саванн, степей, полупустынь и пустынь, а также их широкое распространение по Земле. С распространением саванн и степей повсюду была связана эволюция стадных травоядных животных.
Решающее значение для возникновения альпийской флоры Голарктики имело горообразование в миоцене. Расположение центров многообразия характерных высокогорных родов (например, Saxifraga, Draba, Primula, Gentiana, Pedicularis, Leontopodium, Crépis) показывает, что эволюция этих родов началась в центрально-азиатских горах (например, в восточных Гималаях, Западном Китае, на Алтае). Оттуда многие таксоны расселились, очевидно, через Берингийскую сушу, заселив циркумполярно-арктическую область, а часть их даже добралась через Центральную Америку до Южной Америки (например, Gentianella).
Несмотря на это, нельзя недооценивать и значение европейско-средиземноморских гор как центров образования альпийских таксонов. Это относится, например, к Sempervivum, Helianthemim, Rhodolhanmus, Phyteuma. Achillea, Globularia и Sesleria. Здесь нередко и на примере современных представителей можно проследить последовательную дифференциацию из горных видов субальпийских и далее — альпийских.
Начавшееся в миоцене третичного периода похолодание (рис. 11.299) проявилось сильными колебаниями климата и связанными с ними оледенениями четвертичного периода(примерно от 2 млн лет до настоящего времени; разные авторы датируют начало четвертичного периода, или плейстоцена, 2,5 — 1,64 млн лет). Причинами резких колебаний климата в четвертичном периоде, с одной стороны, являются регулярные изменения вращения Земли и, с другой стороны, особенности земной поверхности.
Характер вращения Земли закономерно изменяется в связи с эксцентричностью ее эллиптической орбиты, с наклоном земной оси и с так называемой прецессией, которой определяется дата перигелия, самого близкого к Солнцу положения Земли. Однако одних этих регулярных изменений вращения Земли недостаточно для объяснения климатических колебаний четвертичного периода, так как они имели место, очевидно, в течение всей истории Земли. Эти колебания могли возникнуть лишь потому, что благодаря тектонике континентальных плит массивы суши оказывались у полюса Морские течения как важные переносчики тепловой энергии также повлияли таким образом, что стало возможным образование полярных массивов льда. Для начала четвертичного периода, очевидно, особенно важным было появление льдов в Арктике, так как Антарктида уже давно была покрыта льдом. Другим важным фактором для четвертичных колебаний климата было, по-видимому, изменение воздушных потоков и глобальных направлений ветров из-за продолжавшегося в четвертичном периоде поднятия гор.
В течение четвертичного периода было много (рис. 11.300) оледенений (криомер) и теплых межледниковий (термомер). В периоды оледенений на северо-западе Европы, в граничащей с ней Северо-Западной Сибири и в обширных областях Северной Америки (на юг примерно до 40° северной широты) образовывались мощные материковые ледники (до 3 000 м толщиной). Альпы также были почти полностью покрыты сплошным ледником (рис. 11.301), в то время как в горах Южной Европы, Азии, Аляски и тропиков были сравнительно небольшие ледники. Во время теплых межледниковий температуры иногда были выше, чем в настоящее время. Оледенениям в высоких географических широтах в более теплых и сухих областях на юге (например, в Средиземноморье, Сахаре) соответствовали дождливые периоды (плювиальные времена), в то время как в межледниковьях там обострялись засухи. На тропических равнинах климат в периоды оледенений был холоднее и заметно суше, что приводило к отчетливому сокращению и фрагментации тропических дождевых лесов.
Рис. 11.300. Климатостратиграфическое деление и реконструированные средние июльские температуры всего четвертичного периода (слева), а также среднего и позднего плейстоцена и голоцена (справа) на примере Нидерландов (по W.H.Zagwijn (слева) и J.De Jong (справа), взято из G Lang, слегка изменено).
Цифры обозначают межледниковые периоды: 3 — Cromer IV, 4 — Cromer III, 5 — Cromer II, 6 — Cromer 1,7 — Leerdam, 8 — Bavel, 9 — Waal C, 10 — Waal A, 11 — Tegelen TC5, 12 — Tegelen TC3, 13 — Tegelen A

Среднегодовые температуры во время похолоданий в Центральной Европе опускались на 8 — 12°С, в удаленных от ледников и тропических областях — на 4 — 6°С. Снежная граница в Альпах проходила на 1 200 — 1 400 м ниже современной (в Центральных Альпах уже на высоте 1 600 — 2 000 м, в Северных Альпах — 900 — 1200 м). Альпийские ледники спускались в предгорья и приближались к северному леднику почти на 500 км. Но климат ледниковых периодов был враждебным для растительности и за пределами оледеневших областей. Здесь широкой каймой вокруг материкового ледника откладывалась принесенная ветром пыль. К северу от Средиземноморья на некоторой глубине в течение всего года сохранялась вечная мерзлота. Связывание огромных масс воды в леднике обусловило понижение уровня моря (примерно на 120 м) и расширение площади материков. Так, Британские острова и южная часть Северного моря во время последнего оледенения были частью материка.
Многочисленные климатические колебания четвертичного периода группируют в шесть ледниковых периодов (см. рис. 11.300), которым в разных (см. рис. 11.300) странах Европы и в Северной Америке даны разные названия. Похолодание началось на границе третичного и четвертичного периодов, было незначительным в претегелине (= брюгге; примерно 2,3 млн лет) и эбуроне (= донау; примерно 1,6 млн лет), усилилось в менапе (= гюнце; примерно 1,1 млн лет) и эльстере (= минделе; около 0,35 млн лет), достигло высшей точки в заале (= риссе; около 0,23 млн лет) и закончились в вейхзеле (= вюрме; около 0,11 млн лет). Между этими холодными периодами были потепления, которые в начале четвертичного периода были более продолжительными, чем похолодания, но в позднем чет
вертичном периоде стали существенно короче, чем периоды оледенений (голынтейнекое и эмское межледниковья продолжались всего по 15 000 лет, а вюрмское оледенение, напротив, около 100000 лет). Сами периоды оледенений прерывались короткими фазами потеплений (интерстадиалами; в вюрмское оледенение их было около 24).
Четвертичные холодные, влажные и соответственно сухие периоды сильно влияли на растительность Земли и приводили к характерным изменениям ареалов и смещениям растительных зон. Многие третичные таксоны вымерли, а новые возникли как следствие географической изоляции, гибридизации и полиплоидизации вследствие постоянных изменений ареалов. Особенно интенсивному воздействию подвергались приледниковые области в Европе и Северной Америке.
Реконструкция растительности Европы во время максимального развития последнего оледенения показана на рис. 11 .301. За исключением лесостепей и лесотундр с березами, соснами и другими холодостойкими древесными породами, распространенными локально, например, у относительно более теплой восточной оконечности Альп, Центральная Европа была тогда безлесной. По ее ведущему виду — арктоальпийской дриаде восьмилепестной (Dryas осtopetala) эти ископаемые флоры были названы дриадовыми. На территории дриадовой флоры были широко распространены кустарничковые тундры и холодные степи, часто на лёссовых отложениях, заку- старенные луга, осоковые болота и бедные видами сообщества водных растений.
Рис. 11.301. Растительность Европы около 20000 лет назад (Вейхзельское оледенение к моменту максимального распространения ледника). Зубчатая линия означает край ледника, белый цвет — тундру, редкий пунктир — тундростепь частый пунктир — степи с изолированными популяциями древесных пород
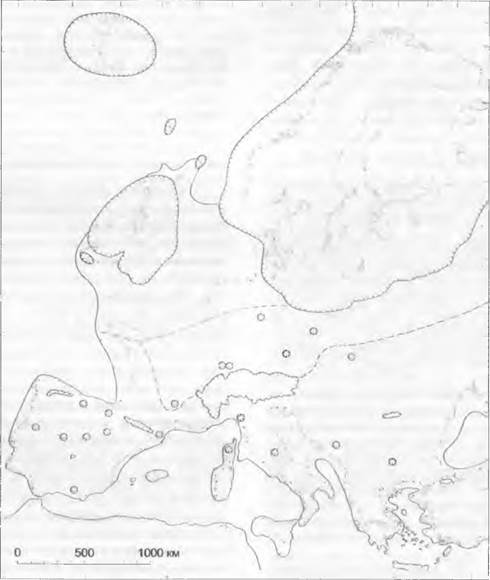
Из видов дриадовой флоры в настоящее время Dryas octopetala, Salix herbácea. Loiseleuria procumbens, Saxífraga oppositifolia, Silene acaulis, Polygonum viviparum, Oxyria digyna иEriophorum scheuchzeri имеют аркго-альпийские ареалы, Salix polaris и Ranunculus hyperboreus сегодня встречаются только в Арктике, a Polenlilla аигеа и Salix retusa— только вальпийском поясе. Вместе с этими видами обитают и такие, которые в настоящее время встречаются еще и между Арктикой и высокогорьями, например, в среднегорье (Betula папа, Empetrum nigrum), или широко распространены, так как проявляют меньшие требования к климату (Filipéndula ulmaria, Ме- nyanthes trifoliata. Potamogetón). В более сухих местообитаниях растут виды, характерные для холодных степей, которые сегодня распространены преимущественно восточнее (Artemisia, Helianthemum, Ephedra). На сырых почвах сегодня как сорняки растут, к примеру, Chenopodium album и Centaurea cyanus Типичными животными этих холодных степей были мамонт, северный олень, овцебык, сурок и лемминг (см. рис. 11.301).
На рис. 11.301 видно далекое отступление в Южной Европе более прихотливых древесных пород. Галерейные и долинные леса сохранялись в области более южных холодных степей, широко распространены были открытые лесостепи и лесотундры, а рефугиумы летнезеленых лиственных или более прихотливых хвойных пород были небольшими, разрозненными и обычно находились вблизи побережий. Вечнозеленая растительность могла сохраниться, вероятно, только за пределами Европы — в СевероЗападной Африке и Юго-Западной Азии.
Идентификация четвертичных рефугиумов с помощью пыльцевого анализа сегодня дополнена применением методов анализа ДНК. При этом принято, что при миграции из рефугиума генетическая изменчивость уменьшается, соответственно в рефугиумах можно встретить больше генетических вариаций, чем в заселенных из них областях (см. 10.2.2). Широкое исследование изменчивости, к примеру, пластидной ДНК в разных частях ареалов некоторых видов деревьев показало, что Quercus robur и Q. petraea, Abies alba и Alnu, glutinosaимели рефугиумы в Юго-Восточной (Балканы), Южной (Италия) и Юго-Западной Европе (Иберийский полуостров), Fagus sylvatica— только в Юго-Восточной и Южной Европе, Picea abies— только в Юго-Восточной Европе. Эти результаты чаще всего подтверждают данные пыльцевого анализа, но иногда приводят и к выявлению новых рефугиумов.
В холодные времена многие виды отступали из гор и Арктики в более низкие или южные места. Они имели хорошие возможности для широких миграций по обширным взаимосвязанным равнинам. Следствием этого стал интенсивный обмен флорами. Это затрагивало не только флоры Альп, Пиренеев, Карпат и других европейских высокогорий. Например, исходно азиатские горные таксоны смогли попасть в Северную Америку через сухопутный Берингийский мост или в Альпы, а альпийские формы — в Арктику и наоборот. В теплые периоды эти виды, с одной стороны, снова заселяли горы и Арктику, с другой стороны, их сплошные во время оледенений ареалы оказывались разорванными распространяющейся лесной растительностью. Эти процессы объясняют многочисленные альпийские, арктоальпийские и азиатскоальпийские дизъюкции в современной флоре, а также наличие арктоальпийских и бореальных видов как ледниковых реликтов за пределами их основного ареала.
Рис. 11.302. Распространение Salix herbácea в Европе в настоящее время (•) и, по данным ископаемых находок, в послеледниковый период вюрмского похолодания (о), а также в более ранние холодные периоды (х)
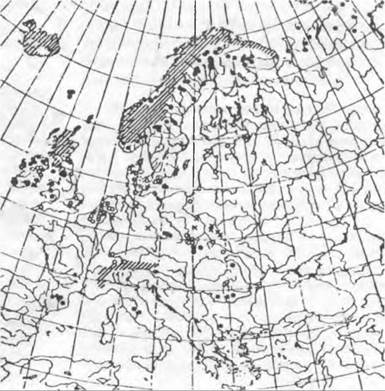
Многие виды Saxifraga, Gentiana, Androsace, Soldanella, Primula, Potentilla и т. п. имеют европейские горные дизъюнктивные ареалы с удаленными частными ареалами в Альпах, Абруцце, Пиренеях, Сьерра-Неваде, Карпатах и Балканах. Арк- то-альпийские виды чаще встречаются во многих изолированных более высоких местах среднегорий (например, в Шварцвальде, Судетах: Saxifraga oppositijblia, Veronica alpina, Gnaphalium supinum). Расстояния между разными частями ареалов достигают часто более 1000 км. Известны, например, очень удаленные находки чисто арктических видов Saxifraga nivalis и Pedicularis sudetica в Исполиновых горах, причем Pedicularis удален от основной части ареала примерно на 2 000 км.
Ива травянистая (Salix herbácea, рис. 11.302) в четвертичном периоде имела арктический ареал (северо-восток Северной Америки, Гренландия, Исландия, Шпицберген, Северная Европа) и во время ледниковых периодов проникла оттуда через Центральную Европу в Альпы, Карпаты, Пиренеи, Абруццу и Балканские горы. Ископаемые находки показывают сплошной ареал, который сегодня сильно разорван, причем между его арктической и альпийской главными частями еще можно обнаружить несколько очень изолированных реликтовых местонахождений (например, в Судетах). Подобное распространение свойственно также Betula папа, имеющей центр ареала в Северной Америке, но достигшей Азии Loiseleuria procumbens, Dryas octopetala и родственным видам, а также циркумполярной Eriophorum scheuchzeri. Из азиатских центров заселили Арктику и европейские высокогорьяSaussurea alpina и Lloydia serótina, тогда как Pinus cembra и Leontopodium alpinum добрались только до европейских высокогорий Alchemilla alpina и Nigritella nigra — примерыальпийских таксонов, которые в ледниковые периоды смогли проникнуть в Арктику.
Сохранившиеся от ледниковых периодов реликтовые местонахождения арктоальпийских или альпийских видов на равнине приурочены к болотам (например, Betula папа, Empetrum nigrum, Trichophorum caespitosum) или топям, оврагам (Saxifraga paniculata, Arabis alpina). В таких местообитаниях локальные микроклиматические условия компенсируют неблагоприятный для этих видов местный климат и сдерживают более сильные конкурирующие виды.
Горные или альпийские сосудистые растения смогли пережить ледниковые периоды как за пределами покровного оледенения, так и иногда в области оледенения на свободных от льда утесах или вершинах («нунатаках»). О положении этих рефугиумов до сих пор судили по сгущениям реликтовых или разорванных ареалов разных видов и соответственно по совпадению расположения ареалов и геоморфологически опознаваемых нунатаков. Часто в ископаемом состоянии не сохраняется ничего кроме пыльцы ветроопыляемых лесных деревьев, поэтому для определения положения рефугиумов все большее значение получает исследование ДНК.
Можно ожидать разных пространственных проявлений генетической изменчивости современных растений в зависимости от того, пережили ли они оледенение вне области покровного оледенения или внутри нее. Таким образом можно показать, что альпийский Eritrichium попит переживал ледниковый период на трех географически изолированных нунатаках, в то время как арктически-субарктический (рис. 11.303) ареал Saxifraga caespitosa и арктическая часть ареала Saxifraga oppostifblia сформировались в результате заселения освободившихся из-под ледника территорий из рефугиумов за пределами оледенения, а не с нунатаков. С помощью молекулярных методов было показано, что рефугиумом Papaver alpinum, вероятно, были западные, северо-западные, восточные и южные отроги Альп вне области покровного оледенения. Сходно расположенные рефугиумы предполагаются для очень многих других таксонов.
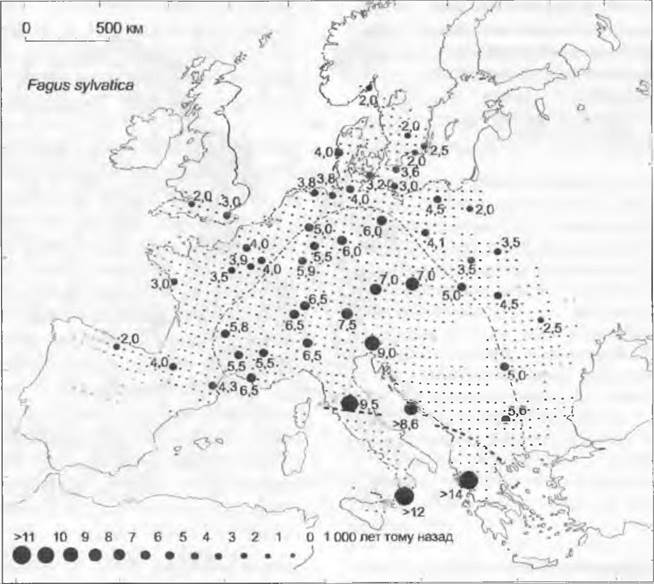
Рис. 11.303. Современный ареал и послеледниковое и голоценовое расселение бука (Fagus sylvatica). Большие черные точки с цифрами означают время в тысячах лет по радиоуглеродным датировкам; маленькие точки — современный ареал; штриховая линия — приблизительные границы ареала 10 000 (жирно) и 5 000 лет назад
Похолодания обусловили, вероятно, и расширение ареалов голарктических видов и родов через экватор в Южное полушарие (например, Quercos, Fagus: см. рис. 14.17), а также некоторые примечательные биполярные дизъюнкции (иногда с промежуточными местонахождениями в тропических высокогорьях).
Биполярно распространенные таксоны находят у лишайников (например. Cetraria islándica), мхов (например, Sphagnum) и покрытосеменных (например, Carex, Epilobium, Empetrum, Primula farinosa agg). Эти таксоны в четвертичном периоде, продвигаясь по Кордильерам и Андам, попали из Северной Америки в Анды Южной Америки и антарктическую Южную Америку. Другие через горы Юго-Восточной Азии и Новой Гвинеи достигли Австралии и Новой Зеландии (которую заселяли также из Южной Америки в результате дальнего заноса, например, Gentianellá), а третья миграция проходила через восточноафриканские горы на юг Африки.
В теплые эпохи четвертичного периода растительный покров был похож на современный. Правда, в ранних межледниковьях в Центральной Европе еще встречались некоторые третичные виды, которые сейчас уже вымерли.
До начала вюрмского оледенения, с которого начался поздний плейстоцен, в Центральной Европе еще были широко распространены некоторые третичные виды: обитающая в настоящее время в Северной Америке и Восточной Азии Brasenia schreberi, эндемичная для Балкан Picea отопка, ограниченные сегодня Северной Африкой и Юго-Западной Азией кедры (Cedrus) и крупноцветковые рододендроны группы Rhododendron ponticum (см. рис. 11.306), которые сейчас встречаются еще только на Кавказе, в Северной Анатолии, Ливане, на Балканском полуострове и в юго-западной части Иберийского полуострова, т. е. в далеко отстоящих частных ареалах.
Начавшийся 10 тыс. лет назад последний отрезок четвертичного периода называют голоценом, или послеледниковьем. После последнего максимума оледенения около 20 тыс. лет назад климат постепенно становился все теплее, несмотря на возвратные похолодания. В последующие 10 000 лет ледниковые массы большей частью растаяли. Благодаря этому многие виды лесов и других климатически более требовательных растительных сообществ заселили до того безлесные или покрытые ледниками области Европы и Северной Америки. В горах это привело к поднятию границы леса, к соответствующему развитию растительных зон и к возврату альпийских тундр на большие высоты.
Обратная миграция лесообразующих пород (рис. 11.303) в позднем плейстоцене и голоцене происходила разновременно. Причина этого кроется в различиях их экологии, конкурентоспособности и скорости распространения, а также в различном расположении рефугиумов. Средняя скорость распространения древесных видов достигала примерно 400 м/год.
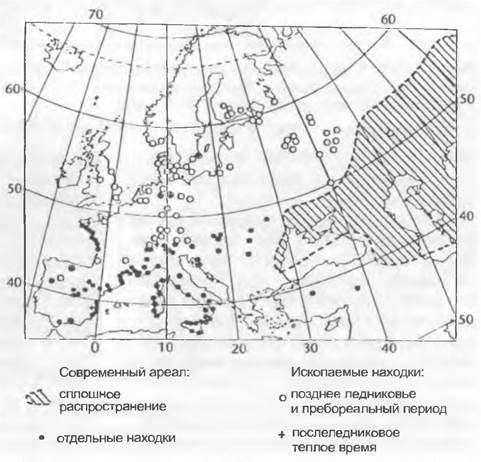
В холодных степях позднего плейстоцена преобладали злаки, Сурегасеае и Artemisia. Ephedra distachya (рис. 11.304) и родственные виды широко распространились по Европе с востока, но были позднее вытеснены развивавшимися лесами в засушливые внутриконтинентальные биотопы и на открытые побережья. Древесные породы позднего плейстоцена — Salix, Betula и Pinas. Тогдашние леса походили на леса современной Лапландии.
Рис. 11.304. Распространение Ephedra distachya agg. в настоящее время и в период от позднего ледниковья до послеледниковья
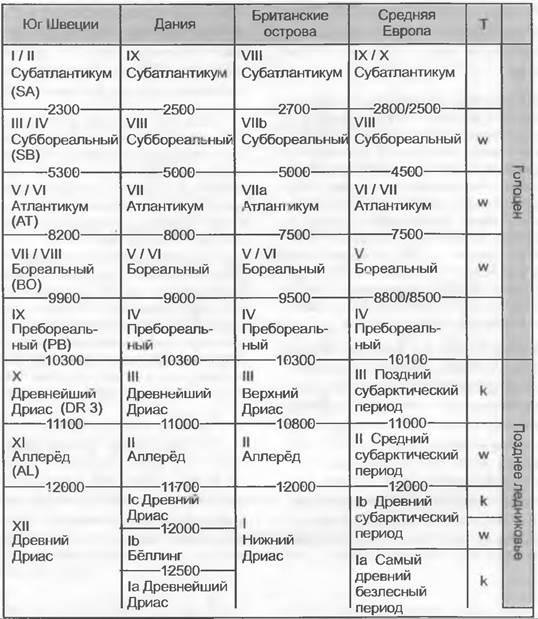
Голоцен начался с потепления климата в пребореальное время (около 10 000 — 8 500 лет), через бореальное время (примерно 8 500 — 7 500 лет) перешел в атлантическое (примерно 7 500 — 4500 лет) и суббореальное время (примерно 4 500 — 2 500 лет), когда был достигнут температурный максимум; в субатлантическое время (2 500 — по настоящее время) началось новое похолодание (рис. 11.305).
Рис. 11.305. Климатостратиграфическое деление позднего ледниковья и голоцена в Северной, Западной и Средней Европе. Временные границы обозначены в годах до нашего времени по данным радиоуглеродных исследований: Т — температура; w — тепло или теплее; к — холод или холоднее
Этому соответствует развитие растительности (рис. 11.306). После короткого похолодания в конце пребореального времени началось бореальное потепление и новое распространение берез и сосен. В атлантическое время начинает массово расселяться лещина, что привело сначала к возникновению сосново-орешниковых лесов. Затем из-за сокращения встречаемости березы и сосны (рис. 11.307), а также усиленного распространения вяза и дуба появились дубово-орешниковые смешанные леса.
С распространением более прихотливых широколиственных деревьев — липы, клена и ясеня — в атлантическое и суббореальное время возникают дубово-смешанные леса. На все сильнее заболачивающихся низменностях распространяются болотные ольшанники, а ели покрывают восточные Средние горы до Гарца, а также восточные Альпы и Карпаты. Сосновые леса, как и современные (рис. 11.307), встречались преимущественно в более сухих и теплых биотопах. В еще более сухих и непригодных для деревьев биотопах развились богатые видами ксерофильные низкотравные луга и степи.
Рис. 11.306. Позднечетвертичная пыльцевая диаграмма с конца ледникового периода до настоящего времени (Luttersee, 160 м, восточнее Геттингена; пыльцевые зоны I — XII по F = Firbas, О = Overbeck). Доля пыльцы деревьев (без клена, Acer) показана черным цветом, пыльцы Corylus и не древесных пород (только важнейшие типы) — белым (и те, и другие даны относительно суммы пыльцы деревьев, принятой за 100 %)
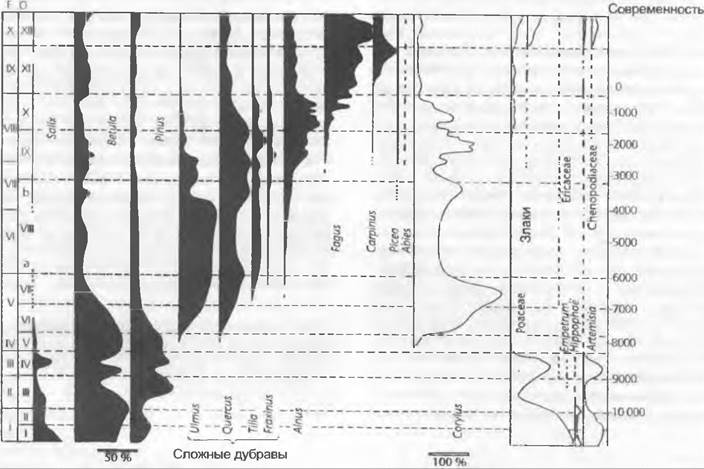
Рис. 11.307. Расширение и сужение ареала сосны обыкновенной в постледниковой Центральной Европе: области одинакового содержания захороненной пыльцы (< 5%; 5 — 50%; 50 — 75%; >75%) в бореальном и суббореальном периодах
Остается неясным, возникли ли подходящие условия для распространения в Центральную Европу теплолюбивых видов каменистых пустошей, сухих низкотравных лугов и степей с юга, и востока лишь в атлантическое и суббореальное время или они господствовали здесь еще до начала повторного развития лесов в голоцене. Исходно более или менее сплошные ареалы этих присредиземноморских или южно-иберийско- понтических видов сегодня чаще всего разорваны, так как распространение лесов очень сузило их жизненное пространство. Такое дизъюнктивное распространение правомерно рассматривать как реликтовое более теплых времен.
Дизъюнктивные ареалы теперь имеют мигрировавшие в теплые периоды голоцена из Средиземноморья в Центральную Европу Quercus pubescens, Comus mas, Buxus sempervirens, Cytisus nigricans, Anthericum liliago и разные виды Ophrys. Вероятно, раньше, может быть в конце ледникового периода, в Европу попали южносибирские и южнорусские степные растения Stipa ser. Capillata, Adonis vernalis, Lathyrus pannonicus, Scorzonera purpurea; богатый видами круг форм Linum flavum
и Astragalus exscapus, а также Eryngium campestre, Salvia pratensis и Odontites luteus попали в ЮгоВосточную Европу из югозападно-азиатского Средиземноморья. Эти виды растут чаше всего на богатых известью почвах в редких зарослях кустарников на скалах, сухих низкотравных лугах и в степных сообществах, иногда — в светлых сосновых и дубовых лесах. Они заселяли с юга Центральную Европу по долинам Роны и Рейна, через южные склоны Альп, вдоль восточных отрогов Альп и по долине Дуная. С востока теплолюбивые виды двигались севернее и южнее Карпат и через Венгрию проникли более или менее далеко в жаркие и сухие, удаленные от моря ландшафты Европы, такие, например, как континентальные долины Центральных Альп, Восточная Австрия, Внутренняя Богемия, Тюрингия и бассейн Майниа.
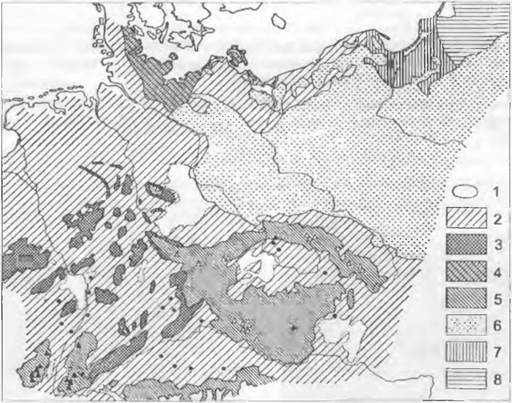
В субатлантическое время заметно снизились температуры и увеличилось количество осадков. Впервые появляются бук европейский, граб и пихта, вытесняя дуб и лещину. Наконец, в историческое время сначала на равнинах, а затем в низких северозападных Средних горах начинает господствовать бук европейский, а на востоке — граб. Горные леса становятся все больше смешанными лесами с буком, пихтой и елью. Так возникла современная растительность, показанная на рис. 11.308.
Рис. 11.308. Реконструкция естественной растительности в Центральной Европе к началу исторического времени (0 г. н. э.) на основании палинологических данных:
1 — сухие разреженные дубово-смешанные леса (без бука, годовые осадки около 500 мм); 2 — буково-смешанные леса глубоких долин (иногда с преобладанием дуба); 3 — буковые горные леса; 4 — область буковых лесов с редкими сосняками; 5 — горные леса с буком, пихтой и елью (▲ — субальпийское редколесье, ∆ — леса с доминированием ели); 6 — сосняки с дубом на песчаных почвах; 7 — грабовые смешанные леса; 8 — грабовые смешанные леса с елью (• — области с локальным доминированием сосны)
Современный человек был современником изменений климата и растительности в Европе с периода примерно 40 тыс. лет тому назад, неандерталец — 200 тыс. лет, но влияние людей на флору и растительность стало заметным в пыльцевом профиле лишь с позднего неолита (около 3 000 лет до н. э.) вследствие перехода к оседлости и начала земледелия.
В почвенных профилях о земледелии свидетельствуют слои золы от выжигания леса и пыльца хлебных злаков и сорняков (Plantago, Rumex, Centaurea cyanus). Увеличение пыльцы хлебных злаков и трав означает растущие пастбищное хозяйство и луговодство. Сорняки как спутники человеческой культуры распространились по всему миру. Археофитами называют такие виды, как Agroslemma githago и Papaver rhoeas, которые попали в Центральную Европу вместе с зерновыми культурами еще в неолите. Sinapis arvensis и Anagallisarvensis как археофиты достигли Центральной Европы лишь в бронзовом веке. Неофиты — виды, вселившиеся в историческое время. К ним относятся, например. Impatiens glandulifera из Гималаев, Senecio vemalis из ЮгоЗападной Азии, Elodea cunadensis, Conyza canadensis и некоторые виды Aster и SoUdago из Северной Америки, a Galinsoga parvifloraи С. ciliata из Южной Америки. Из засушливых областей Нового Света происходит Opuntia ficus-indica и Agave americana, иногда определяющие ландшафты в Средиземноморье. Heracleum mantegazzianum (Кавказ) и Senecio inaequidens (Южная Африка) — также неофиты, которые натурализовались в Европе лишь во второй половине прошлого столетия.
Влияние людей на окружающий мир достигло сегодня масштабов, далеко превосходящих любое естественное изменение в истории Земли. Так, например, в Европе, давно и все более плотно населенной, сохранились лишь последние остатки первобытного леса, небольшие участки тундровой и высокогорной растительности, сообщества неприступных скальных расселин, некоторые низинные и верховые болота, почти нет нетронутых участков водной и прибрежной растительности. Засоление лугов и появление дюн вряд ли прямо зависит от деятельности человека. Эти области (рис. 11.308) занимают сейчас очень маленькие площади и также подвержены непрямому воздействию человека, например, вследствие попадания вредных веществ из воздуха. Считается, что на территории Германии около 1,6 % видов сосудистых растений вымерло, а 12,2% угрожает вымирание («существуют только маленькие, едва ли способные выжить остаточные популяции») или они сильно угнетены («сокращают численность, во многих местах уже исчезли»). Для тропических дождевых лесов, родины около 2/3 всех существующих видов грибов, животных и растений, принимают, что при нынешней скорости (около 1 млн км2 за каждые 5 — 10 лет) продолжающегося вырубания (без особой охраны известных центров разнообразия) в ближайшие 60 лет вымрет приблизительно 5 % всех видов. Подобную угрозу фауне и флоре со стороны человека можно признать и для других областей Земли.