БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
11.2. Бактерии, грибы, растения
Понятия «животные» и «растения» исходно приравнивали к двум основным систематическим группам (таксонам) живых организмов («regnum vegetabile» и «regnum animale» соответственно). В настоящее время известно, что мы имеем здесь дело с типами организации, различающимися по физиологии питания, а не с естественными группами, связанными общим родством. При этом «растительное царство» даже не представляет собой определенной эволюционной линии и соответственно не является каким бы то ни было таксоном. Растения можно определить, как фотоавтотрофные организмы. Соответственно ботаника — это биология фотоавтотрофов. В рамках ботаники рассматриваются все фотоавтотрофные организмы, однако также и те гетеротрофные группы, которые происходят от автотрофов или важны для понимания филогении автотрофов.
В самом широком смысле, следовательно, и грибы (лишайниковый симбиоз!), и прокариоты (гипотеза эндосимбионтов!) являются предметом ботаники и соответственно этой книги. Грибы имеют с типичными растениями некоторые конвер- гснтно возникшие общие признаки (см. ниже).
Ультраструктурнос исследование строения клетки выявило у живых организмов два принципиально различных, в настоящее время не связанных между собой переходными формами плана строения: протоциты и эуциты. Эти два плана строения клетки сравнительно недавно были положены в основу подразделения всего живого на две основные группы (царства): Procaryota (прокариоты) и Eucaryota (эукариоты). Недавние исследования с привлечением методов молекулярной биологии свидетельствуют в пользу факта очень раннего расхождения на три основные группы: археи (Archaea), бактерии (Bacteria) и эукарии (Eucarya), причем Archaea на филогенетическом древе (рис. 11.9) оказываются ближе к Eucarya, чем к Bacteria. Однако на основе строения клетки Archaea и Bacteria как прокариоты противопоставляются Eucarya (эукариотам). Все три основные группы (Bacteria, Archaea, Eucarya) рассматриваются здесь в ранге царств. Чтобы избежать путаницы с применявшимся ранее понятием «царство» («растительное царство», противопоставляемое «животному царству»), вместо понятия «царство» при филогенетической классификации живых организмов все больше используется категория «домен» (домен Bacteria и т. д.).
В приводимой ниже системе рассмотрим Archaea и Bacteria, а из Eucarya — все автотрофные и произошедшие от них вторично гетеротрофные растения (в узком смысле слова), а также гетеротрофные грибы (в широком смысле слова). Как правило, это организмы, ведущие прикрепленный образ жизни, с жесткими клеточными стенками; они могут получать питательные вещества только в растворенном (или же газообразном) виде. Таким образом, сюда не включаются одно- и многоклеточные животные (Arehaeozoa, Protozoa и Metazoa= Zoobionta). При этом необходимо учитывать, что разграничение растений, грибов и животных у низкоорганизованных эукариот (= Protobionta, например, слизевики и эвглены, рис. 11.9) связано с немалыми трудностями.
Рис. 11.9. Филогенетическое древо растений и грибов: cтрелками указаны случаи возникновения эндосимбиозов и как результат этого —возникновение митохондрий и пластид. Стрелки, идущие к лишайникам (Lichenes), показывают установление эктосимбиоза с цианобактериями и Chlorophyta: 1 — возникновение; 2 — приобретение митохондрий в ходе эндосимбиоза; 3 — цианобактерии как первичные эндосимбионты, приобретение «первичных пластид»; 4 — красные водоросли как вторичные симбионты, приобретение «вторичных пластид»; 5 — зеленые водоросли как вторичные симбионы, приобретение «вторичных пластид»; 6 — фотоавтотрофность вследствие эктосимбиоза
В пределах рассматриваемых царств (régna, домены) обширные, однако однозначно произошедшие от одной предковой группы (монофилетические) таксоны трактуются в ранге подцарств (subregna, субдомены) и отделов (phyla). Их названия в случае подцарств оканчиваются на -bionta, в случае отделов автотрофных эукариот на -phyta, а в случае грибов — на -mycota.
Взаимоотношения и возможное родство большинства описываемых в этой книге групп суммируются в кратком обзоре, приводимом после соответствующего раздела. Общий обзор принятой авторами книги системы можно видеть в оглавлении и на рис. 11.9.
Более ранние названия крупных групп (например, водоросли, грибы, семенные растения) в свете современных исследований родственных связей в значительной мере не годятся для обозначения монофи- летических эволюционных линий. В крайнем случае такие названия соответствуют типам организации. Однако для соответствующего большим группам плана строения (типа организации) или одного уровня развития (уровня организации) эти понятия все еще сохраняют определенное значение при обозначении конкретных общностей и для легкого охвата многообразия форм. Поэтому белее ранние обозначения типов организации объяснены в этой книге в специальных вставках— боксах (11.1 — 11.3; 11.5; 11.8 — 11.10), хотя они и не составляют ныне основу для систематического подразделения.
К одному типу организации (любого объема) относят группы организмов, которые по особенностям своей внешней (т.е. морфологической) или также и внутренней (т. е. анатомической и цитологической) организации в значительной мере совпадают. Типы организации часто соответствуют уровням развития и как таковые являются выражением неоднократно возникавших приспособлений к определенным условиям жизни или же общего прогрессивного развития организации.
Таким образом, типы организации охватывают и явно гетерогенные группы. Однако, с другой стороны, они нередко разделяют родственные группы, которые с точки зрения эволюции и систематики должны быть помещены близко друг от друга.
История системы «растительного царства» связана со сменой лежащих в ее основе фундаментальных принципов. Наиболее известная из искусственных систем та, которую предложил К.Линней (1735) на основе половых структур. Он противопоставил 23 классам цветковых растений 24-й класс — Ciyptogamia («тайнобрачные»), к которым отнес не только тогда еще малоизученные папоротники, мхи, водоросли и грибы, но и некоторые высшие растения с трудно распознаваемыми цветками (Ficus, Lemna) и даже кораллы и губки. Подразделения (таксоны) цветковых растений (Phanerogamia, «явнобрачных») Линней различал прежде всего по распределению пола в цветках и по числу, срастанию, расположению и соотношениям длин тычинок. Тайнобрачные ныне можно называть споровыми растениями, так как у них развитие новых особей происходит чаще всего из одноклеточных диаспор (например, спор), а явнобрачные — цветковыми, или, лучше, семенными растениями.
Уже Линней попытался создать естественную систему растений, однако только А.Л. де Жюссье (1789), О. П.Декандоль (1819), Ст.Эндлихер (1836) идр. могут считаться создателями важнейших формальных систем. Даже после появления эволюционного учения в системах А. Брауна (1864), Дж. Бентама и Дж. Д. Гукера (1862 — 1883), А. В. Эйхлера (1883) и особенно в широко используемой и сейчас системе А. Энглера для выделения таксонов сохраняется использование уровней организации и уровней развития.
Первая попытка создания филогенетической системы была сделана Р. фон Веттштейном (1901 — 1908). Современные общепринятые системы представляют собой различные этапы на пути от формальной к филогенетической и синтетической группировкам.
Даже при самом общем рассмотрении всевозможных ныне конкурирующих между собой систем можно увидеть многие, часто глубокие различия. Это показывает, в какой степени систематика все еще находится в состоянии становления. Правда, в последние годы вследствие сравнения и кладистической обработки данных по последовательностям различных гомологичных участков ДНК из хлоропластного, митохондриального и ядерного генома сделан существенный скачок в выяснении родственных связей всех групп организмов.
Таким образом, можно надеяться достичь в обозримом будущем общепризнанной приемлемой естественной группировки.
Точно так же и приводимая здесь система представляет собой попытку в какой-то мере представить в общих чертах родственные связи между большими группами. Однако учитывая учебный характер данной книги, мы сделали в описании определенные упрощения.
К настоящему времени известно около 500000 ныне живущих растений. Из них более двух третей относятся к семенным растениям (около 700 голосеменных и 240000 покрытосеменных), около 10000 — к папоротникообразным и 24 000 — ко мхам. Среди Рrotobionta число описанных водорослей оценивается приблизительно в 23 000, грибов — около 100000, лишайников — примерно 20 000. Наконец, следует учитывать еще 3 000 видов бактерий и 2 000 — синезеленых водорослей. Каждый год описываются все новые виды (особенно грибов и покрытосеменных!), поэтому вполне можно предположить, что еще далеко не законченная инвентаризация растительного царства приведет к тому, что число его видов значительно превысит полмиллиона!
I. Царство (Домен): Bacteria
Царство Bacteria (бактерии, цианобактерии. прохлоробактерии) включает прокариотные организмы (бокс 11.1) с особой химической структурой клеточной стенки (муреиновый мешок).
Два рассматриваемых далее отдела — Posibacteriota и Negibacteriota — образуют группу бактерий в узком смысле слова, они называются также эубактериями (Eubacteria; в противоположность цианобактериям и т. д.).
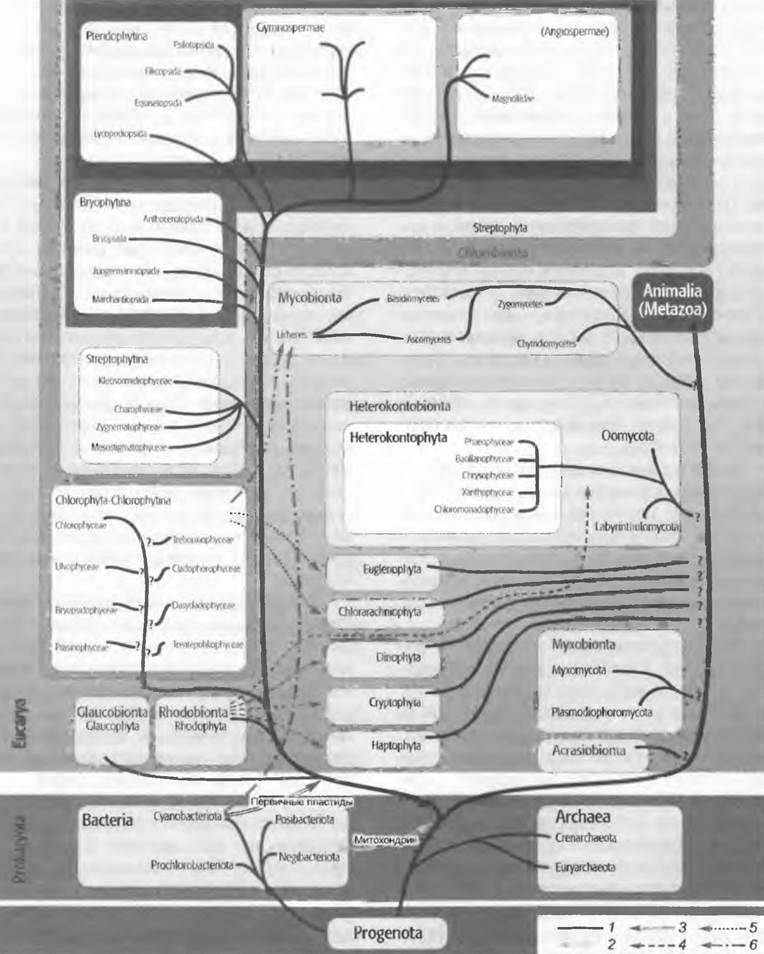
Эубактерии (см. рис. 2.93) — это прокариоты, подавляющее большинство видов которых гетеротрофны (см. 9.1); кроме того, они очень мелкие (см. рис. 1.1) и морфологически малодифференцированны. Различные формы клеток бактерий могут быть выведены из исходной формы шара, а также прямого или изогнутого цилиндра (см. рис. 11.10).
Различают; шаровидные кокки, которые могут объединяться в простые группы типа колоний; палочки, спорообразующие формы которых называют бациллами; формы от изогнутых (вибрионы) до винтообразно извитых палочек (спириллы).
У некоторых бактерий клетки после деления остаются соединенными между собой, образуя скопления из клеток, пакеты (сарцины, рис. 11.10, F), нити (рис. 11.10, Н) или сети. Уже у бактерий можно, таким образом, констатировать тенденцию к развитию более сложных структур. Однако у таких многоклеточных форм принцип разделения функций осуществляется редко (ср. Сhlorochromatium).
Состоящие из суммы клеток нити могут быть простыми или разветвленными; некоторые из них погружены в слизистые влагалища или имеют жгутики. Аналогичную конвергенцию с мицелием эукариотных грибов представляют разветвленные, многоклеточные нити актиномицегов (рис. 11.10, К). Миксобактерии — это гибкие, ползающие по поверхности субстрата прокариоты. Аналогично некоторым эукариотным миксомицетам, отдельные клетки некоторых из таких форм сползаются в плодовые тела размером до 1 мм (рис. 11.10, L — N).
Бокс 11.1. Типы организации прокариот
Прокариотные клетки (протоциты) не имеют настоящего, т. е. ограниченного оболочкой, ядра (отсюда возникло их прежнее название — Аnucleobionta. безъядерные), однако у них есть от одного до нескольких эквивалентов ядра — нуклеоид(ы). ДНК свободно располагается как генофор в так называемой нуклеоплазме. Митоз и мейоз отсутствуют. Подразделение клетки на реакционные пространства (компартменты) выражено меньше, чем у эукариот: отсутствуют хлоропласты и митохондрии. У некоторых прокариот имеются двигательные органеллы, но их структура существенно отличается от структуры аналогичных органелл у эукариот. Стенка прокариотной клетки состоит из гетерополимерных веществ, которые
пока не удалось выявить ни у каких эукариотных организмов. Клеточная стенка представляет собой гигантскую мешковидную полисахаридную молекулу различного химического состава, за счет основных валентностей связанную в сетевидное образование (см. 2.3.3)
Если эукариоты очень зависимы в своей жизни от кислорода, отношение к этому веществу у разных прокариот разное. У них наблюдается переход от полной непереносимости кислорода до обязательной потребности в нем. Только у прокариот имеется широко встречающаяся здесь способность связывать атмосферный азот.
К прокариотам относятся бактерии, цианобактерии, прохлоробактерии и археи.
Рис. 11.10. Bacteria. Формы бактерий: А — Staphylococcus; В — Lactobacillus', С — Bdellovibrio-, D — Spirillum; Е — Caulobacter (4 000х); F — Sarcina; G — J — Sphaerotilus'. G — подвижная стадия (700x), H — форма Sphaerotilus (330x), J — начало разделения клеток (800x); К — Streptomyces; L—N — Chondromyces: L — палочки (200x), M, N — плодовые тела (З0х)
Морфологическая дифференцировка свидетельствует об эволюции в сторону более комплексных структур. Хотя здесь ни в коей мере не достигается комплексность, свойственная эукариотам, все же встречаются уже соответствующие эукариотам формы, выработавшиеся в связи с приспособлением к определенным условиям жизни (колонии, нити, разветвленные нити, мицелий, плодовые тела, споры, жгутики). Однако встречаются и явления редукции, вплоть до размеров вирусов.
Нуклеоид и плазмиды. ДНК бактерий распределена в цитоплазме не диффузно, а локализована в определенной части клетки — нуклеоплазме. Нуклеоид представляет собой клубок из тонких нитей, граничащий непосредственно с цитоплазмой; ядерная оболочка отсутствует. Из-за деления генофора, опережающего клеточное деление, в бактериальной клетке часто встречаются 2 — 4 нуклеоида. У Escherichia coli генофор состоит из одной-единственной кольцевидно замкнутой нити ДНК длиной 1,4 мм. Делению генофора у бактерий, очевидно, предшествует прикрепление его к клеточной мембране (никаких митоза и мейоза!) Кроме генофора в бактериальных клетках встречаются кольца ДНК меньшего размера, способные к самостоятельной репликации — так называемые плазмиды.
Ввиду многих особенностей у бактерий, в отличие от эукариот, ни в коем случае не следует говорить о «клеточных ядрах» и «хромосомах», что, к сожалению, сейчас еще часто делается. Напротив, ДНК бактерий по строению и функциям в значительной мере соответствует ДНК всех других организмов (см. 1.2). При быстрой репликации скорость новообразования ДНК достигает 33 мкм (длина цепочки) в минуту. Бактерии — это те организмы, на которых были получены наиболее существенные данные молекулярной генетики. Для Е. coli, Salmonella typhimurium и других бактерий уже созданы генетические карты.
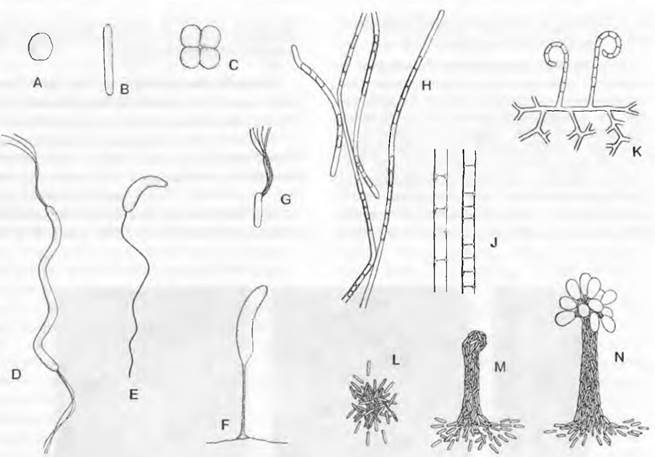
Рис. 11.11. Bacteria. Расположение жгутиков у бактерий: А — Монотрих (Vibrio metchnikovii, 7 000х); В — часть жгутика (Bordetella bronchiseptica, 60 000х); С — базальное зерно у места прикрепления жгутика (Rhizobium radicicoia, 20 000х); D — лофотрих (Spirillum undula, 8 000х); Е — перитих (Proteus vulgaris, содержимое клетки частично автолизировано, 10 000х)
Цитоплазма отграничена от клеточной стенки цитоплазматической мембраной, которая, как и у всех других организмов, многослойная (= плазмалемма). В цитоплазме находятся нуклеоид (иногда несколько), различные мембранные системы и клеточные включения. Рибосомы бактерий, размер которых составляет 16x18 нм, состоят примерно на 60% из РНК и на 40% из белка. Они содержатся в клетке в числе примерно 5 000 — 50 000. Рибосомы осаждаются в ультрацентрифуге при 70S (S — единица Сведберга, коэффициент для определения молекулярной массы). Напротив, эукариоты имеют в цитоплазме 80S-рибосомы, а в митохондриях и хлоропластах — 70S-рибосомы. Мембраны внутри плазмы бактерий образуют (насколько это было исследовано) сеть.
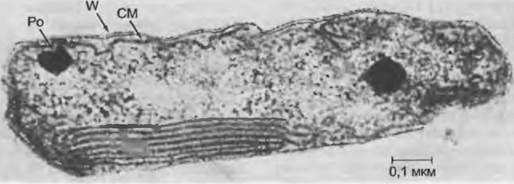
Мезосомы возникают из впячиваний цитоплазматической мембраны. Они были описаны, в частности, как мезосомы и тубулярные тельца. У фототрофных бактерий отмечены также фотосинтетически активные пузырьки, аналогичные соответствующим структурам в хлоропластах зеленых растений — тилакоидам (рис. 11.12); у некоторых видов они даже сходным образом сгруппированы в стопки. Мембраны тилакоидов — носители поглощающих свет пигментов (бак- териохлорофиллов и каротиноидов), а также компонентов систем транспорта электронов и фосфорилирования при фотосинтезе. Правда, тилакоиды у бактерий никогда не бывают окружены общей оболочкой; таким образом, здесь мы имеем дело не с настоящими пластидами. У некоторых бактерий встречаются также газовые пузырьки (например, Chromatiaceae).
Рис. 11.12. Bacteria. Фотоавтотрофная бактерия Rhodopseudomonas с тилакоидами: СМ — цитоплазматическая мембрана; Ро — полифосфатное тельце; W — клеточная стенка
Компоненты клеточной стенки. Откладывающиеся в клетке вещества отчасти являются запасными. Многие бактерии запасают полисахариды типа гликогена. Липофильные граны состоят из поли-β- гидроксимасляной кислоты. Mycobacterium и Actinomyces накапливают преимущественно нейтральные жиры и воски. Фосфорная кислота накапливается в форме полифосфатных гранул («волютина») (см. рис. 2.93).
Клеточная стенка бактерий (см. рис. 2.98) имеет толщину примерно 20 нм. У нее нет фибриллярной структуры, как у целлюлозной стенки клеток высших растений. Ее механическая прочность достигается, как правило, покровом — мешком. Последний состоит преимущественно из полимера муреина, образованного из субъединиц N-aцeтилмураминовой кислоты и N-ацетилглюкозамина; они в чередующейся β-1,4-гликозидной последовательности соединены в полигликановые тяжи. За счет соединения тяжей с короткими пептидами (тетра- или пентапептидами, содержащими D- или L-аминокислоты) возникает сетевидная макромолекула, «муреиновый мешок». Пептидоглюкан у всех бактерий (в том числе и цианобактерий) в более чем 100 вариантах (типы пептидоглюкана) является компонентом клеточной оболочки. У некоторых бактерий на поверхности образуется мощный слой слизи (зооглеи) или «капсулы» различного состава (больше! частью это полисахариды или полипептиды). Клетки Acetobacter xylinum соединяются целлюлозой в пленку; у Sarcina ventriculi клетки также связаны между собой целлюлозой.
Движение бактерий осуществляете» за счет очень нежных плазматических жгутиков (см. 2.3.2; рис. 2.96), которые встречаются на определенных стадиях развития многих бактерий и обеспечивают активное движение клетки в том или ином направлении. При электронной микроскопии эти бактериальные жгутики показывают винтообразную поверхностную структуру (рис. 11.11, В); они составлены из нескольких крайне тонких продольных фибрилл, скрученных между собой. Структуры «2 + 9», как у настоящих жгутиков эукариот, у них нет (см. рис. 2.16). Способность к движению обеспечивает сократительный белок (флагеллин), сходный с миозином мышечных клеток. Диаметр жгутиков составляет чаще всего 10 — 20 нм. длина — до 20 мкм. Они расположены в клетке по-разному: в единственном числе на конце клетки (монотрихи. рис. 11.11, А) или пучками (лофотрихи, как Spirillum, рис. 11.11, D); они могут быть равномерно распределены по всей поверхности клетки (перитрихи, рис. 11.11, Е). Прикрепление к клетке может быть полярным (рис. 11.11, С), биполярным, латеральным (сбоку) или немного ниже конца клетки (субполярным). Каждый жгутик отходит (насколько известно) от базального тельца (рис. 11.11, С), погруженного в оболочку клетки (см. рис. 2.97). Число жгутиков может зависеть от внешних условий: так, Proteus vulgaris при скудном питании имеет 2 субполярных жгутика вместо многих распределенных по всей поверхности клетки жгутиков в обычных условиях. В пучках насчитывается 2 — 50 жгутиков (политрихи).
Движение. Скорость движения с помощью жгутиков составляет, например, у Bacillus megatherium до 200 мкм/с — это примерно 50-кратная длина самой клетки. Spirillum за одну секунду может 13 раз обернуться вокруг своей оси. При этом жгутики совершают 40 оборотов, что соответствует числу оборотов электромотора. Движение происходит, как правило, за счет толчка, как у корабельного винта, однако может заменяться подтягивающим движением по типу самолетного пропеллера. Оно осуществляется чаще всего внутри жидких сред, реже по влажным поверхностям (так движется по агару имеющий перитрихиальное жгутование Proteus vulgaris). В зависимости от действующего фактора движение представляет собой хемотаксис (см. 8.2.1.1), аэротаксис, фототаксис (см. 8.2.1.2) и магнитотаксис (см. 8.2.1.3). Движения в ответ на то или иное раздражение позволяют подвижным формам скапливаться в местах с оптимальным наличием полезных для них веществ. Подвижность имеющей жгутики прокариотной клетки можно в плане строения и эволюции рассматривать как пример конвергенции с аналогичными клетками у эукариот. К скользящему движению способны лишенные жгутиков, похожие на цианобактерии, но гетеротрофные бактерии. Ползающее движение очень медленное (примерно 250 мкм/мин) и связано с выделением слизистых чехлов. О способности к движению миксобактерий по типу голых протопластов упоминалось выше.
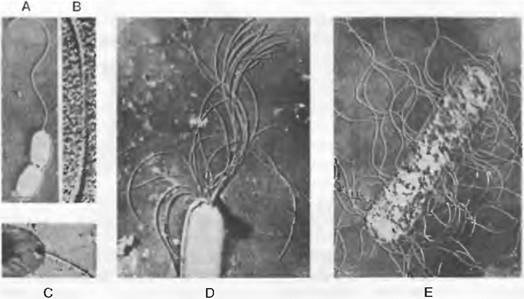
Помимо жгутиков у некоторых бактерий встречаются многочисленные более тонкие нити («фимбрии», или «pili»), функция которых в значительной степени пока неизвестна. У Escherichia coli так называемые F-, или половые, фимбрии обеспечивают парасексуальную конъюгацию (рис. 11.13, А, В).
Физиология. Питание бактерий может различаться в зависимости от источника энергии, донора электронов и источника углерода. Получение энергии происходит
либо путем разложения веществ в субстрате (хемотрофия, см. 6.9), либо за счет использования световой энергии (фототрофия, см.
6.4). В качестве донора электронов служат органические (органотрофия) или неорганические вещества, например, NH3, H2S или Fe+4 (автотрофия). Автотрофные бактерии в зависимости от донора электронов и источника энергии бывают хемолитотрофами или фототрофами. Строго анаэробные виды в присутствии кислорода не могут ни расти, ни размножаться. Факультативные анаэробы могут существовать и без кислорода; микроаэрофильные формы выдерживают лишь незначительные концентрации кислорода. Для облигатно аэробных бактерий кислород совершенно необходим. Распространена — и встречается только у прокариот— способность к фиксации атмосферного азота (см. 9.2.1).
Размножение происходит, как правило, путем деления клеток на две, у вытянутых форм всегда перпендикулярно продольной оси. При этом сначала образуется от края клетки к середине (центрипетально) поперечная перегородка, по которой клетки отделяются друг от друга (отсюда старое название «дробянки» — Schizophyta). У почти всех исследованных до настоящего времени бактерий пептидоглюкановая капсула (насколько она имеется) с самого начала принимает участие в образовании перегородки. Клетки могут после деления оставаться соединенными в рыхлые цепочки (например. Streptococcus).
Для переживания неблагоприятных условий некоторые формы образуют покоящиеся клетки, или споры. У некоторых палочковидных бактерий споры закладываются внутри клеток как эндоспоры; они отличаются от вегетативных клеток меньшей способностью к окрашиванию и сильным преломлением света. Значение эндоспор состоит прежде всего в их устойчивости к нагреванию, благодаря чему они могут сохраняться неповрежденными, например, при длительном кипячении. Вегетативные клетки спорообразующих форм, наоборот, погибают уже при пастеризации (нагревании в течение 10 мин до 80 °С). Спорообразование внутри бактериальной клетки начинается с различных химических превращений в материнской клетке, причем разлагается 75 % ее белков. Затем следует деление материнской клетки на две дочерние клетки неравной величины. Спорообразование завершается формированием вокруг меньшей клетки, становящейся спорой, толстой стенки, которая может составлять до 50 % ее объема и сухой массы. У терморезистентных спор накапливается специфичная для них дипиколиновая кислота. При этом усиливаются терморезистентность и светопреломление.
Спорообразование определяется внешними условиями и осуществляется, например, при недостатке питательных веществ. Готовность к прорастанию спор повышается при хранении и нагревании. Из датированных частиц почвы, которые содержали бактериальные споры (например, почва на гербарных растениях) можно было еще прорастить споры после 200 — 320 лет хранения в сухих условиях. Правда, при хранении в сухом состоянии почвенного образца 90 % спор теряли жизнеспособность в течение 50 лет.
У бактерий возможен частичный перенос генетического материала («парасексуальность»): фрагменты ДНК могут передаваться от клетки-донора к клетке-реципиенту непосредственно путем конъюгации, с помощью бактериофагов или в экстрагированной форме путем трансформации (рис. 11.13, В; относительно функции половых волосков, или пилей: рис. 11.13, А).
Рис. 11.13. Escherichia coii. Перенос ДНК от клетки-донора к клетке-реципиенту (А, В). Бактериофаги (С — Е): А — электронная микрофотография. Клетка-донор и клетка-реципиент (последняя слева) посредством полового волоска (копуляционной фимбрии) соединены между собой (3 500х); В — схема переноса ДНК. Вверху — половой волосок, внизу — реципиент. Донорная ДНК (показана черным цветом) разделена на две цепи, из которых одна проникает в клетку-реципиент. Репликация (светлая линия) одноцепочечной ДНК (темная линия) происходит в половом волоске и реципиенте. Некоторые локусы генов отмечены кружками и буквами (Cat, Lac, Т, Az′, L); С, D — размножение бактериофагов в клетке хозяина с ее разрушением (лизогенный цикл). При смене назад к литическому циклу может происходить трансдукция, т. е. перенос бактериальной ДНК на освободившиеся фаги; Е — отдельные фаги Т2 (40 ОООх)
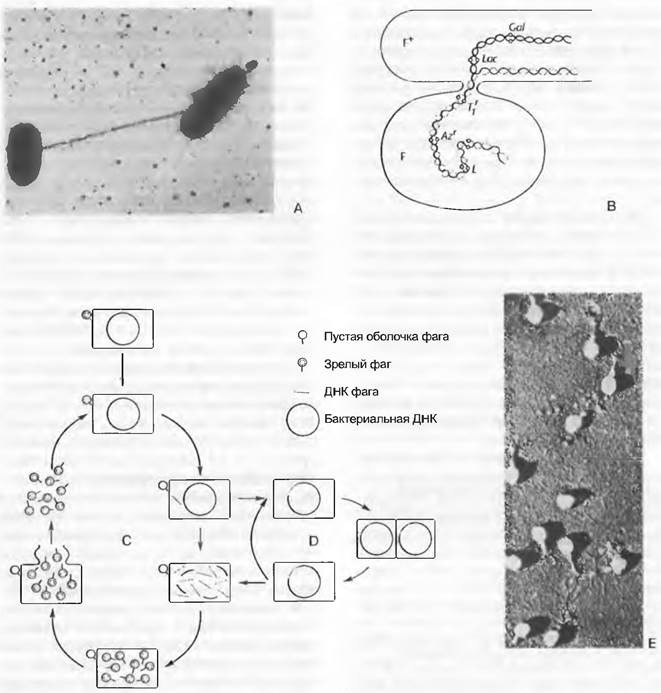
В противоположность чаще всего более крупным бактериям (см. 1.2.5), вирусы, имеющие значительно меньшие размеры и проходящие через бактериальные фильтры, не являются самостоятельными организмами. Они развиваются из генетического материала клеток. Вирусы представляют собой как бы ставшие самостоятельными гены, которые взяли на себя функцию управлять обменом веществ клетки-хозяина для собственного синтеза. Возможно, хотя бы некоторые вирусы даже могли возникнуть путем крайней редукции из патогенных бактерий. Если бактерии содержат ДНК и РНК в соотношении примерно 1: 3,5 %, то вирусы всегда содержат только один тип нуклеиновых кислот — либо ДНК, либо РНК. Вирусы могут воспроизводиться только в живых клетках; они не показывают ни роста, ни деления и нечувствительны к пенициллину и сульфонамидам. На электронной микрофотографии отсутствуют (несмотря на иногда весьма высокую морфологическую организацию) все структуры, характерные для бактерий. Известные примеры болезней, вызываемых вирусами: мозаика табака (ср. рис. 3, т. 1), ящур, бешенство, желтая лихорадка, гепатит, грипп, оспа, герпес. Ретровирусы — РНК- вирусы, у которых одноцепочечная РНК транскрибируется на двойной спирали ДНК (т.е. противоположно обычной транскрипции с ДНК на РНК). К этой группе относятся, в частности, возбудители иммунодефицита (СПИД, ВИЧ) и вызывающие опухоли онковирусы.
Бактериофаги представляют собой особенно высокоорганизованные, относительно крупные вирусы (длина 150 — 110 мкм). Они состоят из «головки», содержащей ДНК. оболочки и белкового «хвоста» (рис. 11.13, Е). Кончик хвоста прикрепляется к поверхности бактериальной клетки, и только после этого содержащаяся в головке ДНК внедряется в тело бактерии через полый хвост. Спустя несколько минут становятся заметны первые признаки образуемых заново частей фага, а еще примерно через такое же время несколько сотен новых фагов высвобождаются из бактериальной клетки в результате ее разрушения (лизиса). Они возникают не путем деления, а путем образования заново из плазмы бактерий. Это явление основано на том, что ДНК фага включается в обмен веществ хозяина, воздействует на его генетический аппарат, и вместо нормальных компонентов бактерии синтезируются специфические компоненты фага. Фаги могут внутри клетки-хозяина изменять за счет мутаций свои биохимические свойства; их можно скрещивать и рекомбинировать. Прежде думали, что фаги, возможно, являются начальным этапом жизни. Однако у них нет собственного обмена веществ и энергии (например, отсутствует дыхание), и поэтому в настоящее время их считают обособившимися частями бактериальной ДНК, которые приобрели способность к самовоспроизведению и даже к сохранению вне клетки в полностью неактивном (латентном) состоянии, пока опять не включатся в обмен веществ хозяина. Существенным подтверждением такого предположения было, в частности, открытие того, что не все бактериофаги смертельны для бактерий, а ДНК так называемых «умеренных» фагов без вреда для хозяина может длительное время реплицироваться бактериальной ДНК (рис. 11.13, С, D). «Генетическое вещество» фагов и бактерий частично очень сходно с таковыми бактериями.
Систематическое подразделение эубактерий. При преобладающей бедности морфологических признаков для подразделения бактерий согласно предполагаемым родственным связям наряду с доступными морфологическими данными особенно большое значение имеют также биохимические и физиологические критерии. Важную роль играет сравнение последовательностей нуклеиновых кислот (см. 1.2).
Бактерии подразделяются на грамотрицательные и грамположительные — это группы, которые могут рассматриваться в ранге отдела. Для их идентификации используется так называемая окраска по Граму (предложена X. К. И. Грамом, 1853 — 1938). Грамположительными называются такие бактерии, у которых анилиновый краситель сохраняется в клетке, а у грамотрицательных бактерий он может вымываться. Для лишенных клеточной стенки бактерий, которые могут возникать как спонтанно, так и в эксперименте (ср. микоплазмы), окраска по Граму не применяется.
Первый отдел: грамположительные бактерии (Рosibacteriota)
Из многослойного муреинового покрова грамположительных эубактерий при окраске по Граму краситель не вымывается. Их клеточная оболочка (см. рис. 2.98, А)
отличается следующими особенностями. Многослойная муреиновая сеть составляет до 30 — 70 % сухой массы клеточной стенки. Особенностью аминокислотного состава является то, что диаминопимелиновая кислота часто заменяется лизином. Полисахариды отсутствуют или ковалентно связаны. Содержание белка меньше. Часто встречаются тейхоевые кислоты — полимеры рибитфосфорной кислоты или глицерофосфорной кислоты, которые через фосфодиэфирные связи соединены с мурамовой кислотой. Среди грамположительных бактерий широко представлены всевозможные морфологические и физиологические типы, известные у бактерий. Наиболее сложной морфологической организации достигли актиномицеты, вегетативное тело которых представляет собой ветвящийся мицелий. Однако образование плодовых тел, как у миксобактерий, а также способность к фотосинтезу отсутствуют. Образование эндоспор встречается только у некоторых палочковидных грамположительных бактерий. Представление о многообразии относящихся к этому отделу форм дает подразделение их на следующие искусственные группы.
1. Кокки. Грамположительные кокки (аналогично рис. 11.10, А, С) могут представлять собой либо одиночные клетки, либо сгруппированные попарно, тетрадами, а также образуют скопления в виде пакетов (у Sarcina ventriculi до 64 клеток, скрепленных вместе целлюлозой). Это облигатные или факультативные анаэробы, иногда аэробы. Пример: молочнокислые бактерии (такие, как Streptococcus lactis) из семейства Streptococсасеае. Сюда же относится Deinococcus (см. 1.1.23).
2. Неспорообразующие палочки. Сюда относятся палочковидные (см. рис. 11.10, В) анаэробные или факультативно аэробные молочнокислые бактерии из семейства Lactobacillaceae (Lactobacillus).
3. Спорообразующие палочки могут образовывать эндоспоры. Они относятся к одному- единственному семейству — Clostridiaceae (Bacillus, Clostridium). Это либо неподвижные, либо (за счет наличия латерально или же перитрихиально расположенных жгутиков) подвижные, аэробные, живущие в почве, или факультативно анаэробные бактерии. Многие из них образуют также цепочки из клеток или нити.
4. Коринеформные бактерии (включая Actinomycetales). Коринеформными называют грамположительные бактерии с сильно меняющимся обликом, т. е. палочки могут становиться булавовидными, короткими, кокками или слабо разветвленными формами. Образование эндоспор полностью отсутствует. К ним относятся пропионовокислые бактерии (Propionibacteriaceae), которые в качестве анаэробных организмов встречаются в рубце и кишечнике жвачных животных. У Mycobacteriaceae, относящихся уже к Actinomycetales, тенденция к ветвлению более выражена по сравнению с ранее рассмотренными формами. Если настоящее ветвление у Mycobacterium встречается только в молодых культурах, то у так называемых «лучистых грибков», или актиномицетов (Actinomycetaceae, Streptomycetaceae, Nocardiaceae), оно является правилом. Многочисленные виды почвенных актиномицетов развивают в искусственной культуре чаще всего колонии диаметром до нескольких сантиметров. Они образуются «мицелием», который часто состоит из одной-един- ственной клетки, лишенной поперечных перегородок, крайне нежной и, как правило, часто обильно разветвленной (диаметр нити 0,5 — 1 мкм; см. рис. 11.10, К). Ни хитин, ни целлюлоза в клеточной стенке не встречаются. Иногда нити становятся многоклеточными и могут легко распадаться на палочки, которые чрезвычайно похожи на некоторые палочковидные бактерии. Кроме того, они образуют (особенно на воздухе) различного рода экзоспоры, располагающиеся в цепочках.
Streptomyces scabies вызывает паршу картофеля и других овощных культур, внешне проявляющуюся в образовании коростинок на пораженной части растений. В корневых клубеньках ольхи и видов некоторых других родов обитает относящийся к актиномицетам симбионт (Frankia alni), который ассимилирует здесь свободный атмосферный азот (см. 9.2. If)- Виды Nocardia, как и представители рода Mycobacterium, могут окислять этан (ср. Flavobacterium). Виды Thermomono- spora и Thermoactinomyces растут при высоких температурах. Продукты выделения некоторых актиномицетов находят применение в медицине как антибиотики для борьбы с инфекциями, вызываемыми патогенными бактериями (актиномицин, стрептомицин и т. д.). В естественных условиях они, вероятно, важны для противостояния микроорганизмам-конкурентам. К группе коринеформных бактерий могут быть отнесены также Clavibacter (см. 9.3.2) и Rhodococcus (см. 7.6.2.1).
5. Микоплазмы (например, Mycoplasma), раньше обозначавшиеся также PPLO (от англ, pleuropneumonia-like organisms — организмы, сходные с возбудителями плевропневмонии), не имеют клеточной стенки и соответственно постоянной формы, поэтому их нельзя отнести к грамположительным бактериям на основании окрашивания по Граму. Однако данные по последовательностям 165-рРНК свидетельствуют о том, что они произошли от грамположительных бактерий. Сюда относится, очевидно, также род Меtallogenium, виды которого участвуют в окислении марганца в водоемах (марганцевые бактерии, см. 6.9.1).
Второй отдел: грамотрицательные бактерии (Negibacteriotа)
У грамотрицательных бактерий муре- иновая сеть тонкая, однослойная и составляет меньше 10% сухой массы клеточной стенки; вещества, поступающие при окраске по Граму, могут легко вымываться. Внешняя мембрана состоит из откладывающихся, однако в любом случае не связанных ковалентно липопротеинов, липополисахаридов и других липидов, которые составляют до 80 % сухой массы клеточной стенки. Ионы Са2+ повышают стабильность липопротеидного слоя. Тейхоевые кислоты (см. рис. 2.90, В) не обнаружены.
К этому отделу относятся кокки, палочки, вибрионы, спириллы, спирохеты и скользящие формы. По способу получения энергии это фототрофы или хемотрофы. В отличие от грамположительных бактерий некоторые представители отдела способны к фотосинтезу, причем здесь он происходит без выделения кислорода (в отличие от цианобактерий и эукариот). Родственные связи фототрофных и нефототрофных грамотрицательных бактерий чаще всего выражены значительно больше, чем связи с цианобактериями, которые при фотосинтезе используют воду в качестве донора электронов. Среди хемотрофных групп различают хемолитотрофные и хемооргано- трофные.
Классификация отдела находится в периоде становления и лишь частично использует возможные родственные связи. Наряду с порядками, в значительной мере являющимися естественными, имеются также различные семейства, объединенные в искусственные группы (1 — 12) по сходству облика и обмена веществ. Мы начнем с кокков и/или палочек и рассмотрим их по группам анаэробов (1), факультативных анаэробов (2) и аэробов (3). Далее будут следовать вытянутые и одновременно спирально закрученные формы, а именно жесткие спириллы (4) и гибкие спирохеты (5), а также бактерии, несущие придатки (6). Исходя из особенностей морфологии, в специальные группы выделяются хламидобактерии (7) и способные к скользящему движению виды из Cytophagales (8) и Мухоbacterales (9); последние образуют структуры типа плодовых тел. По обмену веществ можно различать хемолитоавтотрофные (11) и фотоавтотрофные (12), а по образу жизни — облигатно паразитические бактерии (10).
1. Анаэробные кокки и палочки объединяются в семейства Veillonellaceae и Bacteroidaceae. К последнему семейству примыкают организмы пока еще неясного систематического положения из рода Desulfovibrio. Это полярные моно- трихи или политрихи, представляющие собой по облику вибрионы (палочки, изогнутые в виде запятой) или спириллы (см. ниже, 4). Они относятся к небольшой группе восстановителей сульфатов до сульфитов, способных к «дыханию за счет сульфата» (см. 6.7), и живут как хемолитогетеротрофы. Виды Desulfovibrio— обитатели сапропеля («гнилого ила»).
2. К факультативно анаэробным палочкам относятся, в частности, организмы семейства Enterobacteriaceae (Enterobacter, Erwinia, Klebsiella и часто живущая в кишечнике теплокровных организмов и широко используемая в экспериментальных исследованиях кишечная палочка - Escherichia coli). Виды родов Photobacterium и Вепеккеа из семейства Vibrionaceae приспособились к жизни в морской воде и известны как светящиеся бактерии («биолюминесценция»). Рыбы и головоногие моллюски используют эти виды в своих органах свечения в качестве симбионтов для ориентации и отпугивания. Сюда примыкают роды Haemophilus, Yersinia (бокс 9.2) и Zymomonas. Flavobacterium, также примыкающий к этой группе, отличается способностью к окислению этана.
3. Аэробные кокки и палочки представлены среди грамотрицательных бактерий в большом разнообразии. К ним относятся, в частности, семейства Azotobacteraceae (Azotobacter) и Rhizobiaceae, виды которых часто способны к связыванию свободного атмосферного азота. Первые живут свободно и могут связывать до 20 мг азота на 1 г использованного сахара. Представители Rhizobiaceae (Rhizobium. Azo-, Brady-, Meso-, Sinorhizobium) заражают корни бобовых растений, которые реагируют на это образованием корневых клубеньков (см. 9.2.1). Phyllobacterium rubiacearum — вид, относящийся к тому же семейству, — является симбионтом в листьях различных Rubiaceae (Psychotria, Pavetta) и Ardisia из Myrsinaceae.
Род Agrobacterium не способен к связыванию молекулярного азота. Agrobacterium tumefaciens образует галлы на цветковых растениях (бокс 9.2, рис. А). Он использовался для получения трансгенных растений (см. хронологическую таблицу). Сюда же относятся семейство Pseudomonadaceae (с родами Pseudomonas и Xanthomonas), уксуснокислые бактерии (например, Acetobacter acetï), а также роды Bordetella, Citrobacter и Methylomonas.
4. Спириллы (Spirillales с родом Spirillum) — спирально извитые, жесткие палочки с разным количеством витков — от нескольких до множества. По типу жгутикового аппарата это биполярные политрихи. Чаще всего они являются аэробами, реже — факультативными анаэробами.
5. Спирохеты (Spirochaetales) — чрезвычайно длинные (до 500 мкм!) и тонкие (диаметр 0,1 — 0,6 мм) анаэробные или аэробные бактерии, спирально извитые, как и спириллы. В противоположность последним они гибкие: тонкие клеточные стенки позволяют им при отсутствии жгутиков активно двигаться за счет сокращения осевой нити, находящейся внутри клеток.
6. Бактерии с придатками — формы, имеющие различное эволюционное родство, которые при делении дают дочерние клетки неравной величины, а также образуют придатки в форме палочек и выростов. Палочки состоят из слизи, нитевидные выросты образуются в результате вырастания клетки. Gallionella feirugiпеа известна как железобактерия, которая весной в водоемах, содержащих железо, образует ржаво-коричневые массы. Pedomicrobium тап- ganicum окисляет марганец (марганцевые бактерии, см. 6.9.1).
7. Хламидобактерии обладают трубковидными чехлами (влагалищами), которые удерживают клетки вместе в цепочках. Широко известен так называемый «гриб сточных вод» Sphaerotilus natans (см. рис. 11.10, G — J). Эта бактерия (!) растет в сильно загрязненных водах, например, в отстойниках сахарных заводов. Образуя нити, хлопья и даже наросты, напоминающие густой мех. она может забивать трубы и сточные канавы. Сюда относится также Leptothrix.
8. Порядок Суtophagales. Представители порядка производят скользящие движения. Оба относящиеся сюда рода — Cytophaga и Sporocytophaga — вместе с рассматривающимися дальше миксобактериями являются аэробными почвенными бактериями, разлагающими целлюлозу. В порядке Суtophagales, в отличие от мик- собакгерий не образуется никаких плодовых тел. В качестве скользящих нитчатых форм сюда могут быть присоединены Тhiothrix (см 6.9.1) и Веggiatoaceae. О Сhloroflexus см. ниже (группа 12).
9. Порядок миксобактерии (Муxobacterales) составляет основную часть группы скользящих бактерий. Однако в эволюционном плане они продвинулись дальше предыдущей группы, поскольку обладают комплексной организацией, при которой одноклеточные формы могут образовывать плодовые тела. Красные или другого цвета скопления клеток миксобактерии, живущие на почве или в помете животных, представляют собой колонии («псевдоплазмодий») из мелких, лишенных клеточной стенки, активно изгибающихся палочек без жгутиков, которые за счет активных сокращений клеток могут успешно совершать скользящие движения. У некоторых видов палочки при совместном сползании собираются в определенных местах и образуют характерные, различающиеся в зависимости от рода по облику и окраске, иногда связанные слизью общие скопления — так называемые плодовые тельца, или цистофоры. Из их внутренней части снова могут образовываться подвижные клетки (например, у распространенных видов Мухососсus и у Сhondromyces, рис. 11.10, 1 — 19). В культуре некоторые миксобактерии могут питаться живыми микроорганизмами (например, бактериями). В жизненном цикле здесь наблюдается, таким образом, примечательная конвергенция с эукариотными Acrasiobionta.
10. Облигатно паразитические бактерии объединяются в порядок риккетсии (Rickettsiales). Будучи облигатными паразитами, риккетсии не могут культивироваться вне живых клеток. Они очень мелкие, от вирусов отличаются соотношением ДНК/РНК (1:3,5); кроме того, клеточная стенка здесь чувствительна к лизозиму и содержит мураминовую кислоту.
С риккетсиями, возможно, могут быть сближены долго считавшиеся вирусами возбудители пситтакозов (болезней попугаев). Это приспособившиеся к паразитическому образу жизни бактерии, имеющие ДНК и РНК, а также содержащие специфические вещества (например, мурамовую кислоту).
11. У хемолитотрофных бактерий — в противоположность гетеротрофным бактериям (см., например, группу I) — имеется обязательная взаимосвязь хемолитотрофии с автотрофной фиксацией СO2 (см. 6.9). Аэробные Nitrobacteгасеае окисляют аммиак до нитрита (Nitrosomonas) или нитрит до нитрата (Nitrobacter). Морфологически это могут быть кокки, палочки или спириллы; если у них имеются жгутики, то они прикрепляются субполярно или же клетки являются перитрихами. С этим семейством могут быть сближены бактерии, окисляющие восстановленные соединения серы (отчасти Тhiobacillus) или Fе2+ до Fе3+ (отчасти Thiobacillus и Siderocapsасеае). Наконец, стоит упомянуть водородные бактерии, или бактерии гремучего газа (например, Аlcaligenes eutrophus), которые только факультативно антотрофны. Они могут, с одной стороны, лучше развиваться на органических питательных средах, однако, с другой стороны, могут также окислять молекулярный водород с помощью гидрогеназ. Таким путем, эти бактерии получают энергию, осуществляют процессы восстановительного синтеза и синтезируют специфические углеводы, фиксируя CO2.
12. Фотоавтотрофные Rhodospirillales — в значительной мере анаэробы. Они отличаются наличием различных фотосинтетических пигментов (бактериохлорофиллы а — е) и каротинов. которые придают им характерную пурпурно-фиолетовую, красноватую, коричневую, оливковую или зеленую окраску. Кислород ингибирует синтез и функционирование бактериохлорофиллов, которые этим отличаются также от хлорофилла а цианобактерий (см. рис. 6.45) и эукариот. В качестве доноров электронов используются, в частности, органические соединения (Rhodospirillасеае). Эти приспособившиеся к использованию света как источника энергии фотоавтотрофные бактерии, филогенетически крайне неродственные, встречаются в виде кокков, палочек или спирилл. Если имеются жгутики, то они располагаются полярно или биполярно.
Семейство Rhodospirillасеае — лишенные серы пурпурные бактерии. Как и последующее семейство, отличается наличием преимущественно бактериохлорофилла а или b на цитоплазматической мембранной системе. Элементарная сера ими, как правило, не окисляется. Наиболее известные представители относятся к родам Rhodospirillum, Rhodobacter и Rhodomicrobium.
У представителей обоих последующих семейств в качестве донора электронов используется элементарная сера или сероводород. Сhromatiасеае накапливают в клетках или на их поверхности серу. Чаще всего они имеют пурпурную окраску, поэтому их называют серосодержащими пурпурными бактериями. Такими являются виды Chromatium и Thiospirillum,клетки которых достигают значительных размеров (20 — 40 х 3,5 — 4 мкм), а также Тhiocapsa. Зеленые серные бактерии, относящиеся к семейству Сhlorobiасеае (род Сhlorobium и некоторые другие), не могут ни накапливать, ни откладывать серу. Они содержат бактериохлорофиллы (преимущественно с или у некоторых также есть бактериохлорофилл а в незначительных количествах) в пузырьках, которые расположены вблизи цитоплазматической мембраны или прикрепляются к ней. Эта особенность отличает их от обоих предыдущих семейств. Особые формы известны под родовым названием Сhlorochromatium. Они выглядят как скопления из нескольких неподвижных зеленых серобактерий и расположенной в центре бесцветной бактерии с полярными жгутиками; такое образование движется в пространстве как единое целое. Сhloroflехасеае (лишенные серы зеленые бактерии, объединяемые в род Сhloroflexus), производят скользящие движения и поэтому представляют собой нечто среднее между группами 8 и 12.
Фотоавтотрофные бактерии обитают в анаэробных зонах в пресноводных лужах, прудах и озерах, в медленно текущих водотоках, а также в морских бухтах. Серосодержащие пурпурные бактерии образуют, например, розоватые или винно-красные налеты на разлагающихся частях растений у дна водоемов. Иногда наблюдается их массовое развитие («цветение воды») в более глубоких анаэробных зонах озер. Это происходит при определенных температурных условиях — при достаточно высоких концентрациях сероводорода, диоксида углерода и органических соединений. Благодаря высокому содержанию каротиноидов пурпурные бактерии могут улавливать проникающий в глубину коротковолновый свет и используют его для фотосинтеза. Соответственно на больших глубинах водоемов преобладают пурпурные бактерии, а из серобактерий — имеющие бурую окраску из-за большего содержания каротиноидов.
Встречаемость и образ жизни бактерий
Бактерии, представленные многими видами (около 3 000) и неизмеримым числом особей, встречаются по всему земному шару: в воде, почве, а с пылью — повсюду в атмосфере и на всех предметах. Их широкому распространению способствуют главным образом следующие факторы: во-первых, малый размер и связанная с ним очень большая поверхность по сравнению с массой тела, за счет чего становятся возможными очень высокая физиологическая активность и интенсивность обмена веществ (например, способность к очень быстрому размножению); во-вторых, устойчивость их вегетативных клеток и особенно спор к неблагоприятным внешним воздействиям, а также многообразие способов питания. При оптимальных условиях некоторые виды (например, Vibrio cholerae) могут делиться несколько раз в час, так что от одной бактериальной клетки за 24 часа могут возникнуть несколько миллиардов новых клеток.
Споры бактерий очень устойчивы к высыханию и экстремальным температурам; некоторые выдерживают пребывание в течение нескольких часов в кипящей воде (максимально 30 ч), а также при глубоком холоде. Также и вегетативные клетки многих видов особенно устойчивы к высыханию. Некоторые могут жить при высокой температуре (90 — 110°С), например, в горячих источниках, а некоторые сами активно выделяют значительное количество теплоты («самонагревание» более чем до 60 °С сена, навоза, табака, хлопка, например, такими видами, как Bacillus stearothermophilus или видами родов Thermomonospora и Thermoactinomyces).
Термофилия. Термофильные бактерии, например, виды родов Bacillus, Clostridium, различные микобактерии, а также рассматриваемые далее археи, не только выносят высокие температуры, но и нуждаются в них для оптимального роста. Настоящая термофилия в этом смысле встречается только у прокариот.
К умеренным термофилам относятся бактерии, которые высвобождают тепловую энергию при обмене веществ.
Термофильные бактерии используют термостабильные белки и ферменты, которые отличаются высокими температурными оптимумами. Стабильность белков повышается, в частности, ионами металлов или соединением с клеточными мембранами, а также особым аминокислотным составом; так, термостабильные белки содержат больше аргининовых остатков, чем термолабильные.
Обмен веществ. У прокариот, и именно уже только среди эубактерий, встречается большее число типов обмена веществ, чем среди эукариот. Большинство бактерий живет гетеротрофно — как сапротрофы или паразиты. Однако облигатный паразитизм (например, у риккетсий) редок; большинство патогенных видов может размножаться и вне растительного или животного организма. Поэтому их культивирование на соответствующих питательных растворах (например, мясном отваре с пептоном) не представляет никакой сложности. На твердых питательных средах (агаре, желатине) бактерии часто образуют слизистые скопления разного облика — «колонии» (см. 5.1), которые чаще всего бесцветны, но могут быть и окрашены за счет выделения пигментов. Пигменты в клетках (в тилакоидах, см. рис. 11.12, или же в цитоплазматической мембране) имеют только фотосинтезирующие зеленые бактерии или пурпурные бактерии — аналогично галобактериям из архей.
За счет выделяемых ферментов бактерии вызывают интенсивное разложение субстрата либо в анаэробных, либо в аэробных условиях. Как особенности физиологии обмена веществ разных бактерий можно, в частности, назвать: автотрофность — за счет либо фотосинтеза (красные и зеленые серобактерии), либо хемосинтеза (см. 6.9); гетеротрофность у сапротрофов, у паразитов или в симбиозе; кислородный или бескислородный энергетический обмен веществ; денитрификацию или десульфурификацию (см. 6.6; 6.7); связывание молекулярного азота (см. 9.2.2). Бактерии осуществляют многие процессы брожения: молочнокислое и масляно-кислое брожение, сбраживание целлюлозы, пектина и белка, аэробное уксуснокислое брожение (см. 6.10.2.2). Бактерии могут разлагать почти все природные вещества, даже нефть, парафины, асфальт. Углеводороды разлагаются тем хуже, чем короче их цепи; этан и метан используются только особыми специализированными видами (см. 6.9.1). К бактериальному разложению в значительной мере устойчивы лишь некоторые искусственные смолы и пластиковые материалы, а также особенно устойчивый спорополле- нин (см. 2.2.7.6).
Симбионты. Среди симбиотических бактерий особенно важны для многих сосудистых растений (Fabaceae, Ainus, Hippophae, Ardisia, Pavetta, Psychotria-, cm. 9.2.1) виды, фиксирующие атмосферный азот, в том числе из семейств Rhizobiaceae (Rhizobium, Phyllobacterium) и Actinomycetaceae (Frankia). Полагают, что митохондрии эукариотных клеток возникли путем эндосимбиоза (исходно эндосимбионтами были организмы типа риккетсий).
Возбудители болезней. Многочисленные виды бактерий вызывают болезни животных и человека. Профилактика таких болезней возможна путем иммунизации (вакцинации): при активной иммунизации человеку (или животному) вводится ослабленный возбудитель или его яды, чтобы стимулировать образование антител; при пассивной иммунизации вводят антитела из иммунизированных животных.
Примерами болезней человека, вызываемых грамположительными бактериями, могут быть: гнойные инфекции (Staphylococcus), сибирская язва (Bacillus anthracis), столбняк (Clostridium tetani), дифтерия (Corynebaclerium diphtheriae). туберкулез (Mycobacterium tuberculosis), угри (Propionibacterium acni), актиномикозы (Actinomyces bovi). Близка к грамположительным бактериям Mycoplasma pneumoniae, вызывающая заболевания легких. Грамотрицательные бактерии вызывают воспаление легких и инфекции дыхательных путей (Klebsiella pneumoniae-, Bordetella bronchiseptica; Haemophilus influenzae), тиф (Salmonella typhi), паратиф (Salmonella paratyphi), пищевые отравления (Salmonella typhimurium),чуму (Yersinia pestis), холеру (Vibrio cholerae), венерические заболевания (гонорея — Neisseria gono- rhoeae', сифилис — Treponema pallidum), воспаление головного мозга (Neisseriameningitides). пятнистая лихорадка (Rickettsia sp.).
Фитопатогенные виды внедряются в растение либо через устьица, гидатоды и т. п. (особенно виды Pseudomonas и Xanthomonas), либо инфицируют раны (морозобойные трещины, повреждения насекомыми и т. п., например, Erwinia carotovora). Патогенные бактерии отравляют организм хозяина токсинами. Наличие или отсутствие жгутиков не играет для патогенности никакой роли; странным образом для растений патогенны только палочковидные, неспорообразующие формы. Патогенные бактерии обитают чаще всего в межклетниках и оттуда растворяют срединные пластинки (см. 2.2.7.1), так что отделенные друг от друга клетки отмирают; иногда этот процесс ускоряется еще токсинами. Ткань хозяина превращается при этом в кашеобразную гнилую массу (мокрые гнили). В живые клетки внедряются относительно немногие бактерии (в том числе Pseudomonas tabaci).Изредка бактерии закупоривают сосуды и таким путем вызывают увядание и отмирание растения, чему чаше всего еще способствуют токсины увядания (например, Corynebacteriummichiganense). Известно свыше 200 бактериозов растений.
Биотехнология. Бактерии и другие микроорганизмы могут применяться в технических процессах и промышленном производстве. Это, например, такие области, как генная инженерия (трансформация), получение антибиотиков (также для отщепления боковых цепей синтетически полученных предшественников), ферментов и других белков; разложение отходов (например, метанное сбраживание ила сточных вод); обогащение металлов путем микробного выщелачивания (перевод трудно растворимых соединений меди и урана в растворимые в воде сульфаты видами Thiobacillus). Бактерии, растущие на содержащих нефть субстратах, могут служить индикаторами при поиске новых месторождений.
Третий отдел: синезеленые водоросли, или цианобактерии (Cyanobacteriota, Cyanoprokaryota, Cyanophyta)
Представители этого отдела напоминают водоросли (отсюда название — синезеленые водоросли; прокариотные водоросли) — это морфологически просто устро
енные орагнизмы, осуществляющие фотосинтез. При фотосинтезе высвобождается кислород (оксигенный фотосинтез). Однако в отличие от эукариотных водорослей строение клетки в основных чертах соответствует ранее рассмотренным бактериям, т. е. это прокариоты, и поэтому вместо старого названия «синезеленые водоросли» лучше употреблять название «цианобактерии».
Отличия от эубактерий. Среди прокариот цианобактерии представляют собой относительно гомогенную группу. Определение нуклеотидных последовательностей 16S рРНК показывает более высокий уровень гомогенности между родами цианобактерий, чем между ними и остальными бактериями. От фототрофных родов эубактерий цианобактерии отличаются другими фотосинтетическими пигментами (хлорофилл а вместо бактериохлорофилла) и высвобождением кислорода при фотосинтезе; однако наряду с оксигенным при определенных условиях может осуществляться и аноксигенный фотосинтез. Клетка цианобактерий в среднем в 5 — 10 раз крупнее бактериальной.
Отличия от эукариотных водорослей. Фототрофные цианобактерии (синезеленые водоросли), часто образующие простые или разветвленные нити, будучи прокариотами, отличаются от эукариотных водорослей (см. бокс 11.5) по следующим признакам. В клетках отсутствует оформленное клеточное ядро, митохондрии, лизосомы, эндоплазматическая сеть, ограниченные мембранами хлоропласта и окруженные тонопластом вакуоли с клеточным соком; правда, некоторые цианобактерии имеют, как и некоторые эубактерий, наполненные газом пузырьки — газовые вакуоли. Однако в противоположность всем эукариотам некоторые цианобактерии способны, как и некоторые эубактерий, связывать свободный атмосферный азот (N2). Эта способность обусловлена прежде всего наличием гетероиист (см. 9.2.1), которые отличаются от остальных клеток утратой пигментации, наличием целлюлозы, а также светопреломляющих полярных телец (рис. 11.15, F). Очевидно, образующиеся в гетероцистах соединения азота через тонкие каналы в полярных тельцах переходят в соседние клетки.
Строение клетки. В центральной, бесцветной части клеток (нуклео-, или центроплазме) располагаются элементы в виде гран, палочек, сети или нитей, которые содержат ДНК. Их совокупность, так называемый хроматиновый аппарат, представляет собой эквивалент ядра. При делении клетки весь этот комплекс делится надвое (рис. 11.15,1М). Центроплазма окружена нечетко отграниченной периферической окрашенной хроматоплазмой, в зависимости от формы клетки имеющей вид полого шара или цилиндра. Хроматоплазма очень вязкая и в отличие от протоплазмы эукариотных клеток не находится в движении. В диффузно рассеянных рибосомах она содержит рибонуклеиновую кислоту, а в тилакоидах находится ассимиляционный пигмент — хлорофиллы а и А, а в отдельных случаях также хлорофиллы с и d. В качестве дополнительных пигментов встречаются наряду с каротиноидами (особенно β -каротином, но также и зеаксантином, эхиненоном и миксоксантофиллом, но не лютеином) два водорастворимых хромопротеида (фикоби- липротеида), простетические группы которых (преобладающий здесь фикоцианин, а также фикоэритрин) носят название фикобилины. Фикобилины родственны желчным пигментам и в незначительно отличающейся форме встречаются также у отделов эукариотных водорослей — Сryptophyta и Rhodophyta. Фикобилипротеиды у цианобактерий, как и у красных водорослей, локализованы в тельцах — фикобилисомах (см. рис. 2.89), которые расположены на тилакоидах (рис. 11.14, А). Последние находятся на примерно равных расстояниях друг от друга и не сгруппированы по два, по три или в стопки.
Запасные вещества. Цианобактериальный (цианофициевый) крахмал запасается в виде не различимых при световой микроскопии частиц между тилакоидами. Это глюкан, близкий к гликогену и родственный багрянковому крахмалу красных водорослей. Кроме того, встречаются цианофициновые зернышки, видимые под микроскопом — немного угловатые мелкие тельца, которые состоят из полимеров аминокислот аргинина и аспарагина; очевидно, это азотсодержащие запасные вещества. Фосфор в виде высокополимерных фосфатов накапливается в волютиновых тельцах, построенных из нуклеопротеидов. Возможно, эти тельца также служат как накопители энергии (АТФ).
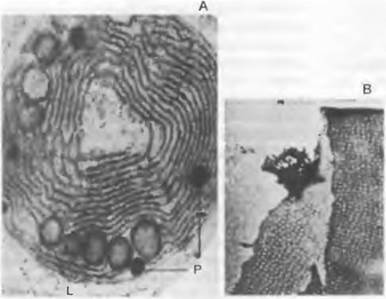
Рис. 11.14. Cyanobacteriota: А — концентрические тилакоиды (25 000>); В — Cylindrospermum, поровый пояс на поперечной перегородке (26 000х); L — липидное тельце; Р — фосфатное тельце
Жесткая клеточная стенка (= защитный слой) состоит из муреина: целлюлоза практически отсутствует (однако см. выше о гетероцистах). Снаружи у цианобактерий часто встречаются слизистые чехлы (влагалища), которые, как показывают электронно-микроскопические исследования, имеют волокнистую структуру и наряду с аминокислотами и жирными кислотами содер жат полисахариды. Клеточная стенка состоит из четырех слоев и разрушается лизоцимом. По ультраструктуре и химизму клеточные стенки цианобактерий занимают промежуточное положение между стенками грамотрицательных и грамположительных эубактерий.
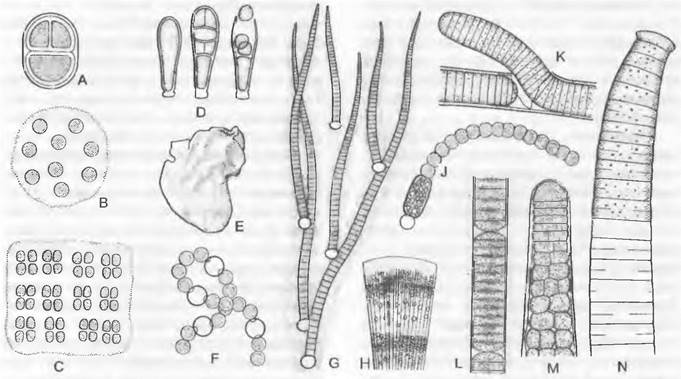
Морфология. Некоторые цианобактерии одноклеточные (например, Dermocагра). Остальные формы представляют собой колонии, содержащие от нескольких до многих клеток (Chroococcus, Мerismopedia), неразветвленные нити без (Oscillatoria) или с гетероцистами (Nostос, Аnabaenа) и нити с гетерополярной дифференциацией (Rivularia), нити с ложным ветвлением (Tolypothrix, Scytonemа) или настоящим ветвлением (Hapalosiphon). Ложное ветвление возникает в результате того, что материнская нить разрывается, а конец дочерней нити, прорывая слизистый чехол, продолжает расти вбок, образуя ветвь (рис. 11.15, К). В противоположность этому настоящее ветвление происходит за счет изменения плоскости деления. Ветвление начинается с клеток, которые за счет деления в другом направлении образуются параллельно продольной оси нити и далее сохраняют такую ориентацию деления. Некоторые нитчатые цианобактерии имеют на продольном и поперечном срезах от нескольких до многих клеток и при этом обладают настоящим ветвлением (Sigonema, некоторые виды с верхушечным ростом; Fischerellа). Здесь клетки делятся в основном более чем в одном направлении. У всех многоклеточных видов цианобактерий фактически мы имеем дело с колониями, в которых отдельные клетки рыхло расположены внутри выделяемой ими совместно слизи или в исходной клеточной стенке (см. 5.1).
Движение. Талломы многих видов, чаще всего нитчатых, совершают скользящие движения (2 — 11 мкм/с). Движение возможно только на твердом и к тому же влажном субстрате. Оно основано не только на простом выделении слизи (ср. движение десмидиевых водорослей, Zygnematophyceae), но и, по-видимому, на работе микрофибрилл. Эти структуры обвивают нить или клетку поверх муреинового слоя и за счет трения о субстрат сообщают вращательное движение. Для упора им служит собственная слизь, выделяемая через тончайшие поры диаметром в 10 нм (рис. 11.14, В) в клеточной стенке. К вращательному движению с перемещением вперед способны только нитчатые Oscillatoriaceae, тогда как представители других групп двигаются вперед без одновременного вращения.
Размножение цианобактерий осуществляется путем клеточного деления. Подвижные стадии со жгутиками отсутствуют. Нитчатые синезеленые водоросли растут интеркалярно путем деления любых клеток в нити. Поперечные перегородки образуются при этом центрипетально наподобие ирисовой диафрагмы (рис. 11.15, N). Они состоят только из материала опорного слоя Размножение происходит за счет неспецифической фрагментации нитей или за счет обособления гормогониев из небольшого числа клеток (рис. 11.15, L). Гормогонии — фрагменты нитей, состоящие из молодых, неспециализированных клеток, — отделяются от материнской нити, уползают и разрастаются в новые нити. У некоторых одноклеточных форм содержимое увеличивающейся в размере материнской клетки последовательно делится на большое число округлых эндоспор, каждая из которых после выхода из материнской клетки вырастает в новую особь. У некоторых видов с вытянутыми в длину клетками базальная часть остается стерильной, тогда как апикальная часть постоянно регенерирует для образования спор (рис. 11.15, D). Эндоспоры цианобактерий отличаются от эндоспор эубактерий своей структурой и развитием. Встречаются также и экзоспоры; они отшнуровываются от материнской клетки. Все эти виды спор не имеют жгутиков. Для переживания неблагоприятных периодов образуются (особенно у гормогониевых) покоящиеся клетки — акинеты. В них превращаются отдельные клетки, в которых откладываются запасные вещества, при этом размер их увеличивается, а стенки сильно утолщаются (рис. 11.15, J). Акинеты прорастают в гормогонии. Однако может быть и так, что боковые ответвления нитей целиком окружаются прочной стенкой и превращаются в покоящийся орган — гормоцисту. Таким образом, имеются различные возможности не только для вегетативного размножения, но и для образования покоящихся органов, которые могут встречаться в жизненном цикле при определенных (т. е. неблагоприятных) условиях.
Рис. 11.15. Cyanobacteriota: A — Chroococcus turgidus (400x); В — Aphanocapsa pulchra (500x); C — Merismopedia punctata (600x); D — Dermocarpa clavata, образование эндоспор; E, F— Nostoc commune: E — колония (1x), F — то же, нить с 4 гетероцистами (400х); G — Rivularia polyotis, часть колонии (200х); Н — Rivularia haematites, часть колонии на поперечном срезе, с отложениями извести и годичными кольцами (15х); J — Cylindrospermum stagnate, с удлиненной покоящейся клеткой и шаровидной гетероцистой у конца нити (500х); К — Ptectonema wollei с ложным ветвлением (200х); L — Lyngbya aestuarii, образование гормогониев (500х); М — Stigonema mamillosum, конец нити (250х); N — Oscillatoria princeps, конец нити (З00х)
Половое размножение неизвестно. Неясно, основан ли наблюдаемый иногда обмен генетическим материалом (в частности, могла происходить рекомбинация факторов устойчивости к разным антибиотикам двух штаммов в одном-единственном штамме) на парасексуальных процессах.
Систематика. Основанное на морфологических признаках подразделение совпадает с молекулярно-филогенетическими данными лишь частично. Сyanoprokaryota (т. е. цианобактерии) составляют единственный класс (Суаnорhусеае) и в зависимости от ступени организации подразделяются на несколько порядков.
1. Порядок: Сhroococcales. К нему относятся одноклеточные формы или простые колонии (шары; таблички; короткие неразветвленные нити). Неразветвленные короткие нити уже могут объединяться в своего рода «псевдопаренхиму». Размножение происходит путем клеточного деления, у некоторых образуются также эндоспоры (например, Dermaсагра; рис. 11.15, D) и экзоспоры. Если при этом дочерние клетки не вырастают до нормального размера материнских клеток, говорят о размножении посредством наноцитов. Synechococcus и Synechocystis — одноклеточные формы. У Сhroococcus и Gloeocapsa (см. рис. 5.2) клетки после деления остаются объединенными в колонии по 2, 4 или 8 внутри общих слизистых обверток, которые могут быть слоистыми (см. 5.1). У Сhroococcus молодые дочерние клетки полушаровидные (рис. 11.15, А), тогда как у Gloeocapsa они яйцевидно закругленные и располагаются в очень толстой слизистой обвертке. Виды обоих родов встречаются чаще всего в слизистых налетах на влажных скалах и стенах. У Aphamcapsa (рис. 11.15. B), Aphanotheca, Microcystis и Merismopedia (рис. 11.15,
C) число клеток, входящих в колонию, больше. Колонии в виде табличек у видов Merismopedia (рис. 11.15, С) возникают в результате клеточных делений строго в одной плоскости; эти виды обитают в пресных водах, некоторые — в морях.
2. Порядок: Oscillatoriales. У этих нитчатых форм еще отсутствуют гетероцисты и акине- ты, только конечные клетки отличаются по облику от остальных клеток нити. Деление клеток всегда происходит в одном направлении, поэтому ветвление отсутствует. Нити часто встречающихся в воде и иле видов Oscillatoria состоят из однородных, часто дисковидных клеток (рис. 11.15, М). Рост осуществляется интеркалярно, а размножение гормогониями. К этому же порядку относятся роды Phormidium, Schizothrix, Spindina, Trichndesmium. Plectonema и Lyngbya.
3. Порядок: Nostocales. Здесь клеточное деление происходит, как и у представителей предыдущего порядка, но перпендикулярно продольной оси нитей (см. рис. 5.1). Размножение происходит, как и у Oscillatoriales, гормогониями. В качестве особых, хорошо заметных форм клеток регулярно встречаются гетероцисты и иногда акинеты. Виды рода Nostoc, которые образуют в воде или на влажной почве округлые или аморфные лопастные колонии, представлены четковидными нитями, погруженными в общую полисахаридную слизь (рис. 11.15, F). Виды родов Cylindrospermum, Aphanizomenon и Anabaena, часть которых — планктонные формы, образуют покоящиеся клетки (рис. 11.15, J). У видов Rivularia (рис. 11.15, G, Н) в колониях, прикрепляющихся к водным растениям и камням, имеется четкое различие межу основанием и концом нити: на нижнем конце находится гетероциста, а кверху нить постепенно вытягивается в бесцветный волосок; таким образом, здесь имеется дифференциация, обусловленная планом строения. Виды Tolypothrix и Scytonema отличаются наличием ложного ветвления. К этому же порядку относятся роды Anabaenopsis, Calothrix и Aulosira (см. 9.2.1).
4. Порядок: Stigonematales. К порядку относятся наиболее дифференцированные формы. В результате поперечных и продольных клеточных делений для них возможны и характерны настоящее ветвление и наличие многорядных нитей. Размножение осуществляется гормогониями. Могут встречаться гетероцисты или акинеты. У Sligonema (рис. 11.15, М) таллом подразделяется на основание и вершину; верхушечная клетка отчленяет вниз сегменты, которые дальше делятся продольными и поперечными перегородками; многорядные нити могут образовывать также боковые ветви. К этому же порядку относятся роды Fischerelia, Hapalosiphon и Маstigocladus.
Встречаемость и образ жизни цианобактерий (синезеленых водорослей)
Цианобактерии, насчитывающие порядка 2 000 видов, распространены по всему земному шару. Часто их можно видеть невооруженным глазом как слизистую массу, тонко-нитчатые налеты, «цветение воды» и т. д. Они обитают прежде всего в пресной воде (даже в горячих источниках с температурой порядка 75 °С). но встречаются также на влажных и сухих почвах, на коре деревьев и скалах — до Арктики и Антарктики. Таким образом, некоторые виды приспособились к жизни вне воды.
Разрушение и образование горных пород. Синезеленые водоросли, обитающие на известковых породах, подвержены воздействию больших колебаний температуры и влажности. Они живут здесь отчасти на поверхности (эпилитно), отчасти в тончайших трещинках (эндолитно) и нередко образуют черные полосы (чернильные штрихи). Некоторые эндолитные виды могут растворять известковые породы, у других же (например, Rivularia, Schizothrix) известь откладывается в слизистых чехлах (рис. 11.15, Н), что в пресной воде приводит к образованию мела и известкового туфа, а в приливно-отливной зоне теплых морей — к отложению слоистых известковых корок (строматолитов). Ископаемые строматолиты обнаружены в докембрийских отложениях, и полагают, что образовавшие их синезеленые водоросли уже в ту геологическую эпоху покрывали большие площади и были широко распространены.
Виды, встречающиеся в больших массах у поверхности пресных и соленых вод, могут вызывать так называемое «цветение воды». Oscillatoria rubescens вызывает в эвтрофных водоемах красное «цветение» и известна как водоросль «бургундского цветения». Другие виды, такие, как Microcystis aeruginosa и Aphanizomenon flos-aquae, образуют ядовитые пептиды, что может вызывать гибель рыбы в пресных водах. Цветение воды, вызываемое Spirulina platensis в соленых озерах Южной Африки, — основной источник питания малого фламинго. При биологическом анализе воды обилие цианобактерий означает критическую степень эвтрофирования (бокс 11.7).
Азотфиксация. Во многих родах (Nostoс, Anabaena и др.) есть виды, которые связывают свободный атмосферный азот (см. 9.2.1). На заливаемых водой рисовых полях цианобактерии ежегодно связывают до 50 кг азота на гектар. В противоположность некоторым эубактериям (Rhizobiасеае) цианобактерии, фиксирующие атмосферный азот, тем самым полностью приспособлены к свободному образу жизни. Вклад синезеленых водорослей в функционировании экосистем при этом, по-видимому, больше, чем азотфиксируюших эубактерий. Среди цианобактерий также больше число азотфиксируюших родов и видов.
Представители нескольких родов вступают в симбиотические отношения с другими организмами. Водоросли некоторых лишайников (см. Mycobionta, приложение 2) — цианобактерии. Некоторые формы живут как эндофиты в полостях тканей других растений, например, Аnabaena — в листьях Аzolla (см. рис. 11.159, О), Nоstос — в талломе некоторых печеночников (Вlasiа, Anthoceros. см. рис. 11.127, D), в корнях Сусах и корневище Gunnera (покрытосеменного растения). В этих симбиотических ассоциациях цианобактерии снабжают своих партнеров азотом.
Четвертый отдел: прохлорофиты (Рrochlorobacteriota, Рrochlorophytа)
Прохлорофиты — одноклеточные, живущие в симбиозе с морскими асцидиями водоросли, у которых отсутствуют фикобилины и к тому же наряду с хлорофиллом а имеется хлорофилл b. По строению клетк это типичные прокариоты: ограниченное мембранами ядро отсутствует, клеточные стенки состоят из муреина. В культуре они размножаются самостоятельно. К этому отделу относятся также свободноживущие нитчатые пресноводные формы, по остальным признакам соответствующие диагнозу Рrochlorophyta.