БОТАНИКА ТОМ 3 - ЭВОЛЮЦИЯ И СИСТЕМАТИКА - 2007
11. СИСТЕМАТИКА И ФИЛОГЕНИЯ
Цель систематического исследования — упорядочить бесконечное многообразие организмов самых разных форм и образа жизни. Для этого необходимо умение распознавать виды и объединять их в систематические группы более высокого ранга (роды, семейства и т. д.). В задачу систематики входит также описание отдельных видов и более высоких систематических групп, присвоение им особых названий и создание определительных ключей для их идентификации. Опираясь на учение о биологической эволюции, созданное Чарлзом Дарвином (см. гл. 10), систематика добивается такого объединения видов в роды, родов в семейства и т. д., которое отражает естественные родственные связи между организмами и группами организмов. Единственное объективное основание для группирования, представляемое самой природой, — история происхождения организмов. В последние годы цель выявления родства (родственных связей) групп стала значительно ближе, так как систематический анализ все в большей мере опирается на информацию, содержащуюся в ДНК организмов. Эту информацию в сочетании с данными о «традиционных» признаках, например, морфологических, в наше время успешно интерпретируют систематически, используя сильно математизированные и потому результативные методы, а также статистически проверяя качество предлагаемых систематических гипотез. Благодаря этому систематика стала методически вполне современной наукой.
Систематика — первый и важнейший шаг в исследовании биологического разнообразия. Она предоставляет справочную основу для всех остальных биологических дисциплин и всех тех частей (отраслей) нашего хозяйства, которые имеют дело с организмами или их продуктами, обеспечивая точную идентификацию и наименование каждого организма и тем самым — ясные связи между организмами. Систематика является также незаменимой основой для понимания эволюционных взаимосвязей биологических явлений. Если, к примеру, цитолог исследует эволюционное происхождение ядерной мембраны, генетик задается вопросом о возникновении интрона, морфолог пытается реконструировать структуру самых первых цветков, физиолог занимается эволюцией различных путей фотосинтеза, а эколог анализирует происхождение разнообразных потребностей растений в составе почвы, то реконструированный систематиком филогенез, в котором определены исходные и производные организмы, представляет собой важнейшее обобщенное знание объекта.
В этой главе сначала представлены методы систематического исследования, а затем дан обзор строения и систематики бактерий, грибов и растений.
11.1. Методы систематики
11.1.1. Распознавание видов
Вид (species) — основная единица систематики. Это не означает, что вид должен быть и основной единицей эволюционных изменений (см. 10.3). Существует множество различных концепций вида (см. 10.3.1), однако большинство видов на практике распознают на основе прерывистых фенотипических вариаций. Таким образом, если можно отчетливо разделить, какую- то труппу индивидуумов, например, по длине листа, характеру опушения цветоножки, окраске цветков и по размеру плодов (см. рис. 10.30), на две подгруппы, причем вариации рассматриваемых признаков внутри этих подгрупп оказываются непрерывными, то данные подгруппы можно рассматривать как различные виды. Этот метод настолько объективен, насколько надежно может быть вычислена и повторно подтверждена наблюдаемая прерывистость мерных и счетных признаков. Во всех этих случаях необходимо прежде всего показать, что мы работаем с однородным или сопоставимым репрезентативным материалом. Впрочем, требуемую для распознавания видов меру фенотипической прерывистости устанавливают субъективно. Фенотипическая прерывистость — это документируемый факт, тогда как отнесение обнаруженных подгрупп исследуемой группы к вариациям, подвидам или видам оказывается лишь формальной проблемой номенклатуры. Для видов общепринято, что относящиеся к ним индивидуумы происходят от одного общего предка и потому не полифилетичны.
Используемые для распознавания видов фенотипические признаки в принципе те же самые (см. 11.1.3.1), что применяются для объединения видов в систематические группы более высокого ранга. На практике большинство признаков анатомо-морфологические, так как первоначальное определение видов чаще всего ведется при работе с музейным материалом — гербарием, и четкая выраженность таких признаков как в природе, так и в коллекциях музеев упрощает отнесение исследуемого индивидуума к тому или иному виду. Огромное значение ботанических музеев (гербариев) состоит прежде всего в том, что только в этих, порой сотни лет собираемых коллекциях исследователь имеет в распоряжении достаточно репрезентативный материал исследуемой группы растений. Самые крупные гербарии мира содержат примерно 6 млн экземпляров растений, и благодаря принятому межгербарному обмену материалом исследователь может получить на время необходимый материал из различных коллекций. Вместе с тем крайне важно исследовать и живые растения, так как определенные признаки, например, число хромосом, можно изучить только на таком материале.
11.1.2. Монографии, флоры и определительные ключи
Важным продуктом систематического исследования на видовом уровне являются монографии как систематические обработки определенного круга родственных форм (например, монография рода Primula) и флоры как систематические обработки растений, населяющих какую- либо географическую область (к примеру, флора Германии). Монографии и флоры, с одной стороны, содержат описания изученных растений, а с другой стороны, делают доступной идентификацию растения благодаря определительным ключам. В монографиях дополнительно содержатся все формальные аспекты систематической обработки (ботаник, первым описавший таксон, — автор вида, рода и т.п., время описания и название публикации, синонимы и т.п.). Описания могут быть весьма различной детализации, но должны ясно показать, как изменчивость внутри вида или группы, так и границы с другими видами и группами.
Определительный ключ может быть построен по дихотомическому или политомическому принципу. Возьмем, к примеру, три признака с разными состояниями (см. 11.1.3.2):
1 - 0 Лепестки красные
1 - 1 Лепестки желтые
2 - 0 Плод ягода
2 - 1 Плод коробочка
3 - 0 Растение однолетнее
3 - 1 Растение многолетнее и распределение признаков по видам:
Вид |
Окраска лепестков |
Плод |
Жизненная форма |
А |
Красная |
Ягода |
Многолетник |
В |
Красная |
Коробочка |
Однолетник |
С |
Желтая |
Ягода |
Однолетник |
D |
Желтая |
Коробочка |
Многолетник |
В этом случае дихотомический ключ — наиболее часто используемая форма определительных ключей — выглядит следующим образом:
1. Лепестки красные 2.
Лепестки желтые 3.
2. Плод ягода вид А
Плод коробочка вид В
3. Растение однолетнее вид С
Растение многолетнее вид D
В политомическом ключе все признаки для одного вида собраны вместе:
Вид Комбинация признаков
А 1-0; 2-0; 3-1
В 1-0; 2-1; 3-0
С 1-1; 2-0; 3-0
D 1-1; 2-1; 3-1
Преимущество политомического ключа в том, что он позволяет определить вид и тогда, когда, к примеру, окраска цветков неизвестна, так как уже одна комбинация типа плода и жизненной формы дает возможность различить все четыре вида. В дихотомическом ключе, напротив, без знания окраски цветков нельзя различить виды А и С или В и D. Правда, данный недостаток можно уменьшить, если включить в тезы и антитезы ключа вопрос не об одном, а о двух или большем числе признаков. Это всегда и делается в хороших ключах, но и здесь может встретиться случай, когда невозможно наблюдать запрашиваемый признак. Политомические ключи все чаще предлагают в электронной форме. Например, предлагаемые в компьютерной программе ’Intkey’ (M.J. Dallwitz) так называемые интерактивные ключи имеют многочисленные преимущества по сравнению с дихотомическими ключами. Прежде всего они состоят в том, что работа по определению может быть начата с любого признака или с любой комбинации признаков. Содержащаяся в монографиях или флорах информация также частично уже доступна в форме компьютерной базы данных.
11.1.3. Исследование родства
Каждую систематическую группу независимо от ее ранга обычно называют таксоном. Степень родства между таксонами определяют по относительному возрасту последнего общего предка. Так, таксон В имеет более тесное родство с таксоном С, чем с таксоном А, если последний общий предок В и С моложе общего предка А и В (рис. 11.1).
Рис. 11.1. Родство. Степень родства определяется относительным возрастом последнего общего предка. Родство таксона В с таксоном С ближе, чем с таксоном А, поскольку последний общий предок (2) В и С моложе, чем последний общий предок (1) А и В
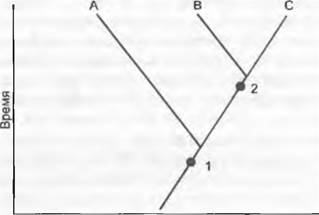
Знание родственных взаимоотношений между таксонами важно с трех точек зрения. Во-первых, родство (о котором недавно можно было сформулировать лишь более или менее хорошо обоснованную гипотезу) — это единственная объективная основа классификации и потому имеет для систематика решающее значение. Во- вторых, классификация, базирующаяся на родстве, имеет известную прогностическую силу. Это означает, что определенное свойство вида может быть с большей вероятностью обнаружено у близких, нежели у дальних родственников этого вида. Так, отдельные алкалоиды группы морфинов мака снотворного (Papaver somniferum) известны только для очень близкородственного мака изящного (Papaver gracile) и для чуть менее близкого мака прицветникового (Papaver bracteatum), но не характерны для более далеких видов мака. Однако родство не всегда имеет прогностическую силу, так как признаки могут быть утрачены. В-третьих, знание родства — единственная возможность реконструировать эволюцию признаков любого вида. Если, например, физиологию растений интересует вопрос, возникли ли среди амарантовых (Аmaranthасеае) С4-расгения из С3-растений только один раз или несколько раз, то ответить на него возможно, рассматривая не сам признак, а родственные отношения таксонов с таким свойством.
11.1.3.1. Признаки
Группы признаков, используемые для определения видов и, более того, для исследования родства, т. е. для объединения видов в таксон более высокого ранга, крайне разнообразны.
Историческое развитие систематики показывает, что расширение области наблюдаемых признаков тесно связано с развитием новых методов наблюдения и возрастанием легкости их использования, а также с общим состоянием теоретических знаний. С античных времен важнейшую базу сравнений составляли признаки габитуса растения, затем с XVI — XVII вв. до К. Линнея и далее вплоть до XIX в. — макроскопические признаки цветков и плодов. С широкого использования микроскопа в XIX в. началось исследование слоевищных растений и их органов размножения (Е. М. Fries, Н. A. de Вагу, А. Pascher, W. Hofmeister, J.-B. Payer и др.) и, кроме того, анатомических признаков побеговых растений (к примеру, В. Н. Solereder, С. R. Metcalfe). После утверждения эволюционной теории все более важным источником исследования родства становится палеоботаника (Н.Solms-Laubach, R. Kidston, W. Zimmermann и др.). Еще во второй половине XIX в. выяснили и значение особенностей ареалов для исследования родства (А. Kerner V. Marilaun, R.v. Wettstein и др.). В 20-е годы XX столетия в систематике начали использовать сведения о числе хромосом и их структуре (например, Е. М. East, Е. В. Babcock), а в начале второй половины XX в большее значение приобрети данные фитохимии (R.E. Alston, R. Hegnauer и др.) и электронной микроскопии (I. Mantón, к примеру, в отношении водорослей). В два последние десятилетия прошлого века все более важными становились данные о строении белков (например, L. D. Gottlieb — изоферменты) и нуклеиновых кислот (например, C.R.Woese, J. D. Palmer, М. W. Chase, D. Е. & Р. S. Soltis), доминирующие в современной систематике.
Большое значение для систематики имеет морфология (см. гл. 4), которая описывает внешнее строение растений. Примеры разнообразия морфологических при
знаков: клеточная организация зеленых водорослей (монадная, коккоидная, нитчатая, тканевая, талломная и т. п.; см. гл. 5; 11.2, Сhlorobiontа), супротивное или очередное расположение листьев, число и расположение тычинок в цветке (см. 11.2, расположение частей цветка). Для оценки структурных гомологий морфологических признаков часто необходимо изучить их развитие (онтогенез). Так, например, многочленные андроцеи могут возникать совершенно разным способом (см. рис. 11.173). Признаки внутреннего строения исследует анатомия. При этом гистология (см. гл. 3) занимается структурой тканей, как, например, коллатеральным или биколлатеральным расположением ксилемы и флоэмы в проводящих пучках покрытосеменных растений или наличием настоящих тканей или плектенхимы у зеленых и красных водорослей (см. 11.2, Сhlorobiontа, Rhodophytа). Цитология (см. гл. 2), еще одна часть анатомии, имеет дело с тонким строением клеток. Так, большое значение для систематики имеют структура базального тельца жгутиков (см. рис. 11.88) и детальный ход клеточного деления у зеленых водорослей или электронномикроскопическая структура пластид ситовидных трубок у покрытосеменных (см. рис. 11.167). Кариологиякак часть цитологии исследует хромосомы клеточного ядра. При этом выявленная с помощью различных методов окрашивания структура хромосом в интерфазе и преимущественно в метафазе, а также весь хромосомный набор клетки оказываются вариабельны при сравнении различных таксонов. Разделами анатомии можно также считать палинологию и эмбриологию. Палинология занимается структурой спор и пыльцевых зерен (см. рис. 11.174,11.175, 11.176). Здесь выявлены различия в характере слоев стенки спор, видимой при электронной микроскопии скульптуры поверхности спор, в числе, форме и расположении апертур. Развитие спорангиев, гаметофитов, гаметангиев, эндосперма и зародыша — область эмбриологии. Различия в возникновении выстилающего слоя (тапетума) спорангиев лежат, к примеру, в основе подразделения папоротников на группы, а развитие зародышевого мешка у покрытосеменных варьирует по числу участвующих в его образовании мегаспор и конечному числу слагающих клеток (см. рис. 11.187). Структуру содержащихся в растениях веществ изучает фитохимия. При этом речь может идти о пигментах фотосинтеза (различные хлорофиллы и дополнительные пигменты; см. 6.4.2), запасных углеводах (крахмал, инулин) или о таких общих для большого круга покрытосеменных растений классов веществ, как алкалоиды, терпеноиды или флавоноиды (см. рис. 11.168). Систематик может получить, кроме того, важные признаки также из области физиологии (например, С3-, С4-фотосинтез; см. 6.5), экологии (например, требования к почве), географического распространения таксона, выявляемого географией растений, или из фитопатологии (например, в отношении специфичности патогенных грибов или травоядных животных; см. 9.3,9.4); особенно важно при изучении родства принимать во внимание исследованные палеоботаниками ископаемые формы. В настоящее время утрачивает свое значение экспериментальная систематика, занимающаяся гибридизацией и интерпретирующая успех скрещивания как критерий родства. Напротив, сегодня для исследования родства особенное значение приобрел анализ протеинов и нуклеиновых кислот. В то время как серологический анализ протеинов (сравнительное осаждение протеинов специфическими антителами) и анализ аминокислотных последовательностей в них сейчас также теряют свое значение, анализ изоферментов (выявление числа генетических локусов и числа и качества аллелей ферментов путем электрофореза; см. бокс 10.1), как и прежде, остается обычным приемом для характеристики генетической конституции популяций или близкородственных видов. Анализ РНК и главным образом ДНК начался с практически утратившего сейчас значение метода гибридизации ДНК, когда заключение о степени родства основывали на фотометрически регистрируемой скорости образования двойных цепочек ДНК из отдельных участков ДНК, выделенных из различных видов. Анализ ДНК методом обработки ферментами-рестриктазами, разрезающими ее на специфические участки (например, из 6 или 4 пар оснований) (RFLP), также отходит на задний план. При RFLP-анализе ДНК разных видов нарезают с помощью одинаковых рестриктаз. Различия в получаемых фрагментах обусловлены мутациями в области нарезанных участков. Эти мутации используют как признаки. При преобладающем теперь секвенировании РНК/ДНК определяют последовательность нуклеотидов в нуклеиновой кислоте и каждую позицию нуклеотида рассматривают как признак. Сегодня в систематике растений наиболее часто используют, пожалуй, нуклеотидные последовательности генов или участков генов пластидной rbcL (большая субъединица рибулозо-1,5 - бифосфаткарбоксилазы; другие все чаще используемые гены пластид — это, к примеру, matK и ndh F или же определенные пластидные нитроны и спейсеры) и рибосомальной ДНК клеточного ядра. К ри- босомальным ДНК клеточного ядра принадлежат как ITS -последовательности (внутренний транскрибируемый спейсер), так и 16/18S последовательности РНК, которые кодируют малую субъединицу рибосомы. Начали также использовать и митохондриальные гены. Предпочтительное применение пластидной ДНК в сравнении с ядерной обусловлено тем, что большинство ядерных генов представлено целыми семействами, вследствие чего при их гомологизации возникают проблемы, в то время как пластидные гены чаще всего имеются в единственном числе. Правда, пластидная ДНК чаще всего наследуется только по материнской линии (см. 10.1.2.5). Особенно информативными могут быть выяв- [яемые при секвенировании или RFLP- анализе макромутации ДНК типа инсерций или инверсий, поскольку они редко возникают параллельно у разных видов. Для анализа ДНК на низком таксономическом уровне сегодня еще используют множество методов так называемых отпечатков (см. бокс 10.1), с помощью которых, чаще всего с применением полимеразной цепной реакции (PCR), иногда удается даже различить отдельные индивидуумы.
Строение ДНК во многих отношениях имеет преимущество перед другими признаками растений. Это определяется тем, что данные о ДНК возможно однозначно кодировать. В последовательности ДНК можно точно определить положение нуклеотидов. Четко отнести морфологический признак к одному из альтернативных состояний, напротив, часто очень непросто или даже невозможно из-за существования переходных форм. Это особенно проявляется в работе с близкородственными видами, где различия часто только количественные. Другое преимущество данных о ДНК в том, что они позволяют сравнивать друг с другом организмы, имеющие мало общего в фенотипических признаках (например, одноклеточные водоросли и семенные растения), но с одинаковыми генами (например, rbcI). Из строения ДНК можно получить, наконец, очень много признаков, но при этом следует учитывать, что при секвенировании одного гена анализируют множество нуклеотидных последовательностей, хотя рассматривают только один ген. В противоположность этому анализ фенотипов включает обширную область различных признаков и тем самым — также очень большое и совершенно неизвестное число генов.
11.1.3.2. «Конфликты» признаков
В зависимости от исследуемого таксона и его систематического ранга используют разные группы признаков. Для дальнейшего обсуждения ценности признаков имеет смысл разграничить признак и его состояние (англ. character, character state). Так, например, окраска цветка — признак, а красная, белая, синяя и т. д. — состояния этого признака. Необходимость развития методов оценки признаков следует из того, что чаще всего не все признаки подтверждают одинаковое группирование объектов. При рассмотрении нескольких признаков в сущности всегда встречается «конфликт» признаков.
При рассмотрении, например, трех таксонов — А, В и С и двух признаков, имеющих состояния 0 и 1, может оказаться, что признак 1 противопоставляет группу А + В таксону С, а признак 2 — таксон А группе В + С (рис. 11.2).
Рис. 11.2. «Конфликт» признаков. Признаки 1 и 2 со своими состояниями 1-0, 1-1, 2-0 и 2-1 по-разному указывают на родство между таксонами А, В и С: признак 1 показывает группирование А с В, признак 2 — В с С
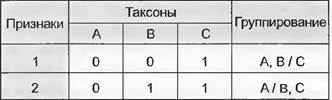
«Конфликты» признаков имеют место, во-первых, из-за того, что при сравнении двух (и более) таксонов идентичные проявления признака могут быть результатом конвергенции или параллелизма (см. 10.4), т. е. возникали в процессе эволюции неоднократно и независимо друг от друга, что вовсе не очевидно из структуры самого признака. В то время как для обоснования соответствия у двух таксонов какого-либо структурного признака, например, морфологического, дополнительно к его особому качеству можно привлечь его относительное положение и онтогенез, для оценки идентичного нуклеотида, находящегося в одинаковой позиции в ДНК двух таксонов, таких дополнительных критериев нет. «Конфликты» признаков возникают, во- вторых, потому что значимость признаков для выявления родства различается в зависимости от времени их возникновения относительно времени возникновения исследуемой группы (см. 11.1.3.4).
Для разрешения «конфликтов» признаков используют разные пути. Первая возможность состоит в том, чтобы приписать различным признакам разный «вес» и таким образом решить проблему. Насколько несомненно то, что разные признаки имеют разные «веса» вследствие своей разной сложности, настолько же трудно объективно оценить их различные «веса». В результате возникают варианты классификаций, имеющие сильный субъективный компонент. Соответственно они могут сильно различаться, несмотря на одинаковый набор используемых признаков. Объективизацией представлений о классификации организмов занимаются, с одной стороны, нумерическая систематика, а с другой стороны — филогенетическая систематика.
11.1.3.3. Нумерическая систематика
Нумерическая систематика (фенетика) занимается парными сравнениями всех таксонов для установления сходства их друг с другом и расчета структуры сходств в исследуемой группе. В нумерической систематике анализируемые таксоны обычно называют операциональными таксономическими единицами, «operational taxonomic unit» (ОТU). Конкретный способ действий состоит в том, чтобы с самого начала (как и во всех других методах обработки данных) выяснить выраженность признаков у всех таксонов исследуемой группы. Затем можно закодировать их в так называемой бинарной матрице данных, в которой таксоны помещают напротив признаков (рис. 11.3, А). Так, например, для признака «окраска цветков» проявления признака красный цвет или белый цвет кодируют произвольно как 0 и 1. На следующем этапе попарного сравнения всех таксонов рассчитывают сходство (или дистанцию, принимая 1 за сходство). Простой способ его вычисления состоит в том, что число признаков, общих двум таксонам, делят на общее число наблюдаемых признаков (рис. 11.3, В). Рассчитанный таким образом коэффициент сходства известен и как коэффициент простого совпадения («simple matching coefficient»). Наконец, на третьем этапе, используя все коэффициенты сходства, таксоны объединяют в группы со все уменьшающимся сходством. Представленная на рис. 11.3, С фенограмма является результатом так называемого кластерного анализа, в котором таксоны располагаются в иерархическом порядке. Различные группы, или «ветви», расходящиеся от места разветвления, называют также кластерами.
Рис. 11.3. Нумерическая систематика: А — матрица данных для таксонов от А до F и 10 признаков, имеющих состояния 0 и 1 каждый; В — парные сходства между таксонами (отношение числа общих признаков к общему числу признаков); С — рассчитанная по коэффициентам фенограмма
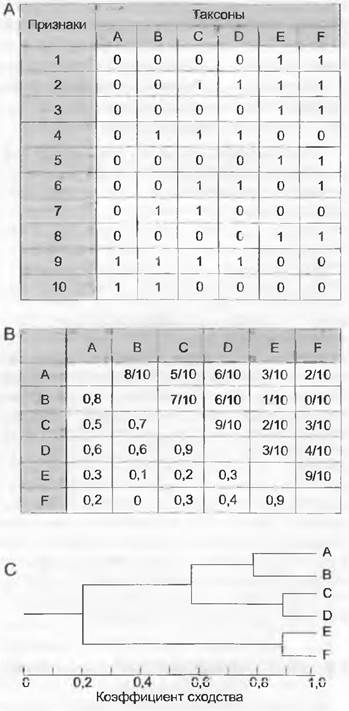
Только что описанный метод можно видоизменять во многих местах или использовать различные опции. Так, некий признак (например, окраска цветков) может иметь не два, а большее число состояний (например, красный, белый, желтый (англ. multistate character). Этот вариант можно рассчитать, как и признаки, только с двумя состояниями. При кодировке количественных признаков, таких, как длина листа, следует создать классы значений, которые обозначают, например, все листья короче 10 см как 0, а все листья длиннее 10 см — как 1. Такие классы обычно задают произвольно. Но в принципе существует возможность отказаться от кодирования таких количественных признаков и рассматривать простую разность между выраженностью признака у двух таксонов как меру сходства (дистанции) по этому признаку. Кроме очень простого коэффициента простого совпадения можно использовать большое число других коэффициентов сходства или дистанции. С их помощью можно, например, учитывать, обусловлена ли общность между двумя таксонами отсутствием или наличием проявления признака. Коэффициент Жаккара не учитывает сходство по отсутствующим признакам. При рассмотрении, например, признаков последовательности ДНК не учитывают увеличение вероятности многократных и конвергентных изменений одного нуклеотида с увеличением дистанции между двумя таксонами (например, дистанция Кантора, дистанция Кимуры). Наконец, есть много разных приемов кластерного анализа и еще так называемый ординационный анализ. Например, метод главных компонентов («principalcomponent analysis», РСА) в отличие от кластерного анализа дает неиерархическое представление отношений сходства. Обычно применяют - в особенности для молекулярных данных — метод UPGMA (метод невзвешенного парно-группового среднего, «unweighted pair group method using arithmetic averages») и соседнего связывания («neighbour joining»,NJ). В то время как UPGMA поставляет «правильные» результаты только когда темп эволюции во всех эволюционных линиях одинаков, NJ может справиться и с различными темпами эволюции благодаря постоянному пересчету исходной матрицы дистанций.
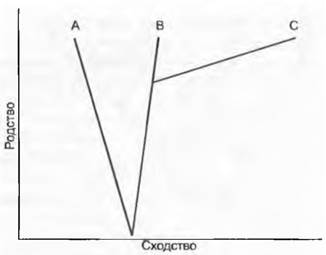
Претензия фенетики на объективность во многих отношениях ограничена. Как и все другие методы, она начинает с отбора признаков и их оценки, для чего нет никаких объективных критериев. Специфическая проблема фенетических методов состоит в том, что как для вычисления коэффициентов сходства, так и для дальнейших расчетов парного сходства есть множество различных способов, которые могут привести к разным результатам, но нет объективного критерия выбора этих способов. Исходя из этого, представляется принципиальным вопрос, можно ли отождествлять сходство и родство, так как именно родство является единственным объективным критерием группировании. Сходство и родство тождественны только в том случае, когда темп эволюции во всех эволюционных линиях одинаков и нет параллелизмов и конвергенций. Если ускоренная эволюция в одной линии развития, например, при завоевании совершенно нового жизненного пространства ведет к сильной фенотипической дивергенции, то сходство будет неверно отражать родственные отношения (рис. 11.4). Первоначально, впрочем, фенетика и не претендовала на реконструкцию родства.
Рис. 11.4. Сходство и родство. При различной скорости эволюции сходство не отражает родство. Хотя А и В более сходны, чем В и С, более близкородственными оказываются В и С
11.1.3.4. Филогенетическая систематика
Метод филогенетической систематики (кладистики) разработан энтомологом В.Хеннигом (W.Hennig). На первом этапе отбирают все признаки. Второй этап анализа заключается в оценке признаков по относительному времени их появления. Те признаки, которые встречаются только внутри исследуемой группы (англ.: ingroup) (рис. 11.5), называют относительно продвинутыми, или апоморфными (апоморфия), а те, которые имелись уже вне исследуемой группы, считают исходными, или плезио- морфными (плезиоморфия). В зависимости от того, имеется ли апоморфия только у одного или у нескольких таксонов, говорят об аут- или синапоморфии, а при плезио- морфии используют соответственно понятие симплезиоморфия. Оценка состояния признака как апоморфного или плезиоморфного относительна, потому что она меняется с объемом исследуемой группы. Так, обладание обоеполыми цветками для покрытосеменных в целом — плезиоморфный признак (в сравнении с однополыми цветками), так как обоеполые цветки — это исходное состояние у этих растений (см. 11.2. Цветок). Для всех семенных растений, однако, обоеполые цветки — это апоморфия, так как это состояние признака встречается только у покрытосеменных и может быть использовано для обоснования монофилии данной группы. Важнейшее достижение Хеннига состояло в обосновании того, что для установления родства годятся только апоморфные, но не плезиоморфные признаки. Так, если бы, например, при сравнении автотрофной астры (Аstегасеае/Аsteralis) с автотрофным коровяком (Scrophulariасеае/Lamiales) и паразитической заразихой (Orobanchaceae/Lamiales) пришли к выводу о более близком родстве астры и коровяка на основании наличия у обоих хлорофилла, то этот вывод был бы ошибочным. Причина в том, что наличие хлорофилла — исходное свойство зеленых растений и как плезиоморфия не может служить для установления родства в одной маленькой их подгруппе. У астры и коровяка хлорофилл сохранился, а у заразихи был утрачен.
Рис. 11.5. Сравнение с сестринской группой. Если в исследуемой группе таксонов А, В, С проявления признака для А будут 1 -0, для В — 1 -0 и для С — 1 -1, а проявление признака 1 -0 у сестринской группы Б считать плезиоморфным состоянием, то придется постулировать только одну трансформацию признака: 1-0 —> 1-1 в линии, ведущей к С. Если же принять за исходное состояние (1-1), то потребуются две трансформации признака: или трансформация признака (1-1 —> 1-0) имела место как при возникновении А, так и при возникновении В, или эта трансформация ((1-1 —> 1-0)) произошла у общего предка А, В, С уже после возникновения Б, а при возникновении С случилась обратная трансформация: ((1-0 -» 1-1)). Сравнение с сестринской группой служит основой принципа экономии при распознавании апоморфных признаков
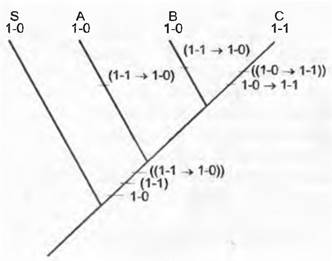
Для решения вопроса, является ли состояние признака апоморфным или плезиоморфным, важнейшим методом служит сравнение с сестринской группой (англ. sister groupcomparison). При этом принято считать, что состояние признака, встречающееся у ближайшего родственника исследуемой группы (сестринской группы), плезиоморфно. Причина такого допущения в том, что оно экономнее (англ. more parsimonious), чем альтернативное допущение о возможности апоморфного проявления признака в несестринской группе1. Если в исследуемой группе из трех таксонов А, В и С состояния признака следующие: А 1-0, В: 1-0 и С 1-1, а состояние признака 1-0 у сестринской группы S считать плезиоморфным, то требуется только одна трансформация признака (англ, character transformation) от 1-0 к 1-1 в линии, ведущей к С (рис. 11.5). Если же принять, что исходным состоянием было 1-1, потребовались бы две трансформации признака: либо трансформация от 1-1 к 1-0 имела место дважды при возникновении А и В, или же эта трансформация имела место у общего предка А, В и С после возникновения S, а при возникновении С произошла обратная трансформации от 1-0 к 1-1 (рис. 11.5). Так как лишь апоморфный признак дает возможность судить о родстве, одинаковое состояние признака 1-0 у А и В не свидетельствует о тесном родстве этих таксонов. Скорее у А и В сохранилось состояние признака общего предка.
1 Авторы нечетко представляют кладистический принцип парсимонии. Речь должна идти о независимом появлении одинаковых состояний признака в исследуемой и сестринской ей группах. — Примеч. ред.
Под экономностью (англ. parsimony) как причиной выбора информативного признака не следует подразумевать, что и сама эволюция должна была бы быть экономной и всегда идти кратчайшим путем. Экономность — скорее общий научный принцип, который требует минимального числа гипотез, необходимых для объяснения.
Даже если известен самый близкий родственник исследуемой группы, необходимо кроме сестринской группы дополнительно провести сравнение ее с другими родственными группами. Сестринская группа, и эти другие родичи, вместе взятые образуют внешнюю группу (англ. outgroup). Понятие «внешняя группа» более общее, чем понятие «сестринская группа». Проблематичность сравнения с сестринской и внешней группами в том, что принимается неизменность раз возникших признаков в этих группах. Так как сестринская группа геологически того же возраста, что и исследуемая группа, а остальные таксоны внешней группы геологически старше, они имели столько же (сестринская группа) или даже больше времени (другие таксоны внешней группы) для эволюционного преобразования, чем сама исследуемая группа. Поэтому просто невероятно, чтобы с момента возникновения исследуемой группы ее сестринская или внешняя группа не изменились.
После того как на втором этапе анализа признак оценен как апоморфный или плезиоморфный, на следующем этапе для установления родства оставляют только апоморфные признаки. Если при первичном отборе признаков правильно распознаны все случаи параллельного развития — параллелизмы (англ. parallelism), а также случаи обратного развития признаков — реверсии (англ. reversal), то апоморфные состояния признаков должны быть непротиворечивыми. Однако так почти никогда не бывает. Противоречивость признаков, вызванную параллелизмами или реверсиями, называют гомоплазией (англ. homoplasy), состояние признака называют соответственно гомопластичным, если он в исследуемой группе возникал более одного раза или после возникновения вернулся к исходному состоянию. Существующие «конфликты» признаков разрешают таким образом, чтобы, используя подходящие методы вычисления и следуя принципу экономии, минимизировать число необходимых в целом трансформаций признаков.
Этот последний шаг — важнейший принцип используемого сегодня чаще всего максимально экономного кладизма, который практически заменил филогенетическую систематику в ее первоначальной только что описанной форме и который использован в компьютерных программах, например, PAUP (’phylogenetic analysis using parsimony’). Принцип максимальной экономии состоит в отказе от предварительной оценки состояния признака как апоморфного или плезиоморфного и тем самым от сравнения с сестринской внешней группой; все признаки рассчитывают так, чтобы число необходимых трансформаций признака в полученных кладограммах было минимальным. Несмотря на это, сестринскую/внешнюю группу включают в анализ. Предпочтение отдают самой экономной (англ. most parsimonious) кладограмме, которая первоначально не имеет исходной точки («неукорененная» кладограмма — англ. unrooted). «Корень» как исходную точку такой кладограммы располагают соответственно результатам сравнения исследуемой и внешней групп. Оценка признаков как апо- или плезиоморфных при использовании метода максимальной экономии получается лишь после того, как кладограмма рассчитана и «укоренена». Различные группы, расходящиеся от точки разветвления, или «ветви» кладограммы, называют также «кладами» (’clades’).
Так называемая эволюционная систематика пытается, хотя и без формализованных методик, oтразить в классификации как сходство, так и родство. Еще и сегодня часто используемые системы покрытосеменных растений, например, системы А. Сгот- quist, R. M.T.Dahlgren, А.Л.Тахтаджана и R. F. Thorne, представляют, как раз эту эволюционную систематику.
11.1.3.5. Метод максимального подобия
В интерпретации признаков последовательности нуклеотидов ДНК все чаще применяют метод, названный методом максимального подобия (maximum likelihood).
Исходным пунктом этого метода является формулировка модели эволюции последовательности нуклеотидов, специфичной для исследуемой группы. С одной стороны, такая модель учитывает вариации последовательности в исследуемой группе, к примеру, оценивая частоту субституций на основе наблюдаемой изменчивости в каждой позиции или из вариаций последовательности, делая вывод об относительной частоте транзиций и трансверсий. С другой стороны, в модели эколюции последовательности дополнительно принимают общие допущения, например, о независимости друг от друга позиций последовательности. Затем рассчитывают родословное древо, которое объясняет с максимальной вероятностью (maximum likelihood) наблюдаемые в исследуемой группе вариации последовательности на основе принятой модели эволюции последовательности, специфической для исследуемой группы.
11.1.3.6. Статистический анализ гипотез о родстве
Гипотезы о родстве, рассчитанные с помощью фенетики, кладистики или метода максимальной экономии, в известной мере доступны статистической проверке их стабильности. Один из таких методов — бут- стреп-анализ. Путем случайного выбора из оригинальных данных и многократных повторений составляют новую матрицу данных, а затем анализируют ее. Эта новая матрица имеет такой же размер, как и оригинальная матрица, но может отличаться от нее тем, что один признак, например, использован трижды, зато два другие вообще не включены в матрицу. Величина бутст- реп-индекса для одной клады 90 означает, что в 90 % повторных анализов различных матриц данных положение таксонов на этой кладе идентично с положением в анализе матрицы оригинальных данных. В кладистике обычно используют Decay-индекс (а также Bremer-Index). Здесь определяют число дополнительных шагов (трансформаций признаков), необходимых для того, чтобы разрешенная клада (т. е. показывающая родственные отношения между А, В и С; см. рис. 11.1) преобразовалась в политомию (когда невозможно различить родственные связи А, В и С друг с другом). Значения Dесау-индекса I, к примеру, означало бы, что одна из разрешенных ветвей (клад) в максимально экономном родословном древе становится неразрешенной в древе, которое только на один шаг длиннее максимально экономного. Увеличение Dесау-индекса интерпретируют как увеличение обоснованности гипотезы о родстве.
11.1.4. Филогения и классификация
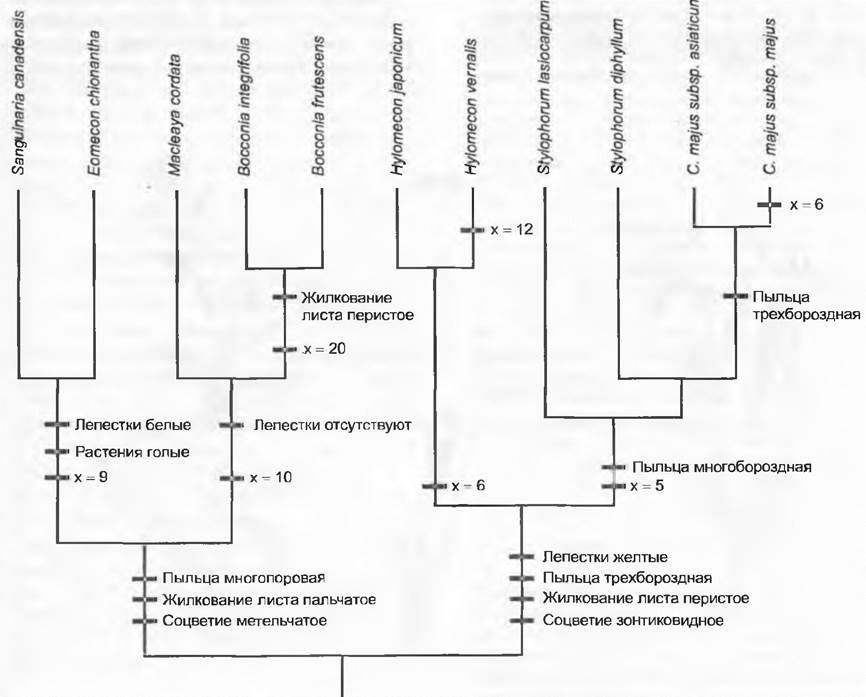
Выше описанные возможности систематического анализа ведут к созданию гипотез о родственных отношениях между таксонами, т. е. филогении таксонов. Особая проблема, равно присущая фенетическим и кладистическим методам, состоит в том, что они с трудом разрешают сетчатую эволюцию (т. е. возникновение нового таксона путем гибридизации дивергировавших линий развития), столь обычную у растений (см. 10.3.3). Как фенограмма, так и кладограмма не являются собственно родословным древом, потому что не дают сведений о предках или потомках. Филогению таксона можно использовать для того, чтобы выявить эволюцию признака. Простое нанесение признаков на фенограмму или кладограмму показывает, как изменялся признак в процессе филогенетического развития таксона. Это представлено на примере форм, родственных чистотелу большому (Сhelidonium majus: Рарауегасеае- Сhelidonioidеае) (рис. 11.6). Фенограмма или кладограмма должны быть основаны на таких признаках, которые независимы от тех, чью филогению собираются реконструировать.
Рис. 11.6. Эволюция признаков. Нанесение признаков на кладограмму показывает, как мог бы меняться признак в филогенезе таксона. В круге родства чистотела (Chelidonium majus)многопоровые пыльцевые зерна возникли у общего предка Sanguinaria, Еотесоn, Macleaya и Bocconia, а имеющиеся у общего предка Hylomecon, Stylophorum и Chelidoniumтрехбороздные пыльцевые зерна стали многобороздными у предка Stylophorum и Chelidonium. При возникновении Chelidonium вновь появились трехбороздные пыльцевые зерна
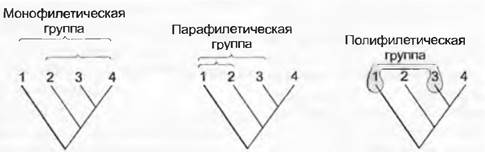
При переводе фенограмм и кладограмм в формальную классификацию следуют многократно принятому принципу филогенетической систематики, согласно которому надо признавать только монофилетические группы. Монофилия (рис. 11.7), по определению, — это тот случай, когда таксон содержит всех прямых потомков одного общего предка. Парафилия — это тот случай, когда все включенные в таксон группы происходят от одного общего предка, но не все потомки этого предка включены в данный таксон. При полифилии в один таксон включают прямых потомков разных предков.
Рис. 11.7. Моно-, пара- и полифилия. Монофилетический таксон (3-4, 2-4, 1-4) содержит всех потомков одного непосредственного общего предка. Парафилия наблюдается тогда, когда все включенные в таксон группы (1-2, 1-3) имеют одного общего предка, но не все потомки (3-4, 4) этого предка включены в данный таксон. Полифилетический таксон содержит подгруппы (1 и 3), которые произошли от разных предков
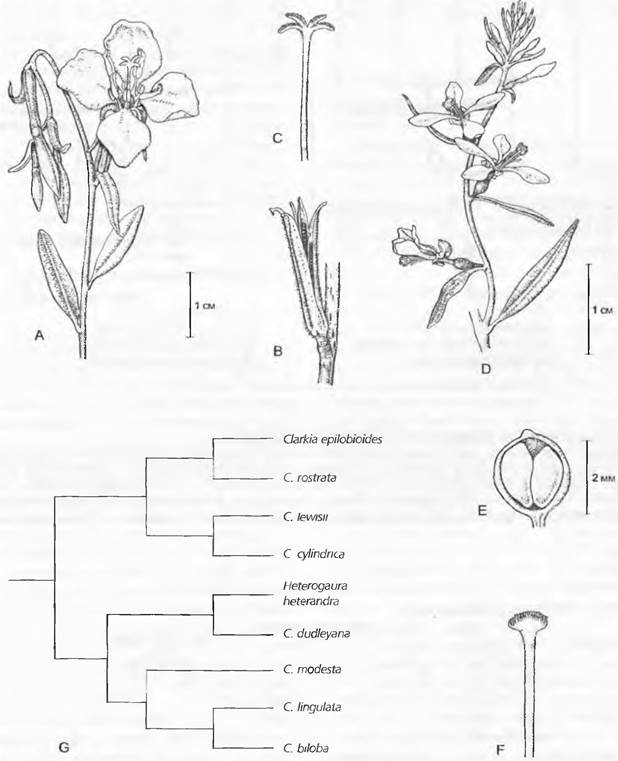
Один из многих известных примеров парафилии — это два североамериканских рода: Сlarklа и Нeterogaura (Оnagrace). В то время как Clarkia обладает двумя кругами
фертильных тычинок, раздельным рыльцем и плодом коробочкой, Неterogаига имеет только один круг фертильных тычинок, головчатое рыльце и плод орех, благодаря чему этот таксон и был описан как особый род. Молекулярные анализы ясно показали, что Heterogaura близкородственна некоторым видам Сlarkiа и, таким образом, возникла из рода Сlarkiа(рис. 11.8). Если сохранить Heterogaura как род, тогда Сlarkiапо отношению к Heterogauraпарафилетична, так как, хотя все представители Сlarkiа и сводятся к одному прямому общему предку, но вследствие исключения Heterogaura не все потомки этого предка включены в один таксон. Отказ от парафилетических таксонов прежде всего предполагает, что критерием для классификации должно быть не сходство, а родство. Поэтому Heterogaura, явно непохожую на виды родаСlarkiа, следует классифицировать как представителя рода Сlarkiа, потому что ее самые близкие родичи относятся к Сlarkiа1. Молекулярные анализы показали также, что двудольные растения парафилетичны относительно однодольных (см. рис. 11.213). Однако многие оспаривают требование филогенетической систематики не учитывать в классификации плезиоморфные признаки.
1 Этот таксон целесообразно включать в род Сlагкiа потому, что некоторые виды последнего более близкородственны Heterogaura, чем другим видам Сlarkiа. — Примеч. ред.
Рис. 11.8. Парафилия (А — F). Сlarkiaа (А — С) и Неterogaura (D — F) различаются числом фертильных тычинок, формой рыльца и типом плода. Анализ родства показывает, что выделение Неterogauга heterandra в самостоятельный род ведет к признанию парафилии рода Сlarkiа
11.1.5. Номенклатура
Для наименования и классификации растений существует много формальных правил, которые содержатся в регулярно перерабатываемом «Международном кодексе ботанической номенклатуры». Формальные аспекты систематического исследования называют также таксономией. Из-за крайне неоднозначного определения этого понятия в литературе мы здесь от него полностью отказались. Правда, понятия «систематика» и «таксономия» обычно рассматривают как синонимы.
В системе растений используют ряд обязательных таксономических рангов, или категорий. При этом речь идет об абстрактных понятиях упорядочивания, которые в иерархии занимают совершенно определенные положения. Название вида представляет так называемый бином (двойное название), состоящий из названия рода и видового эпитета(например, Achillea millefolium). Полное видовое название включает также фамилию автора, который описал этот вид. У Achillea millefolium L. буква L означает Линней. Названия надвидовых таксонов состоят из одного слова (униноминальны) (например, Achillea). В табл. 11.1 представлены важнейшие таксономические категории, их общепринятые окончания, а также соответствующие названия конкретных таксонов на примере тысячелистника обыкновенного (Achillea millefolium). Новый таксон в ботанике тогда считают действительно обнародованным (англ. validly published), когда его описание удовлетворяет определенным критериям. Это выбор названия в соответствии с твердо установленными правилами, наличие диагноза, публикация в одном из общедоступных литературных источников, а также указание так называемого типа (см. ниже), который должен быть сохранен и общедоступен. Условия доступности для всех исследователей литературного источника и типа определены в «Кодексе». Все научные названия растений используют в латинизированной форме. Для родовых названий, а также названий таксонов более высокого ранга используют имена существительные, а для видовых эпитетов (и других названий внутривидовых категорий) чаще всего употребляют прилагательные. Первоописание в ботанике должно быть составлено на латинском языке. Благодаря типификации описанное название типа однозначно связано с конкретным растительным индивидуумом. Тип названия вида или внутривидового таксона чаще всего представлен растением, сохраняемым в гербарии. Только в исключительных случаях соответственно исторической традиции типом можно считать изображение растения или его описание. При этом тип вовсе не всегда типичен для таксона. Род типизирован видом, а семейство — родом. Если для таксона существует несколько законно опубликованных названий, то вступает в силу правило приоритета. Используют то название, которое было законно опубликовано первым. Это правило не относится к таксонам выше ранга семейства, а иногда могут быть признаны законными («законсервированы») и не приоритетные названия. Позднее опубликованные названия получают статус синонимов законного названия.
В недавно широко обсуждавшейся «филогенетической номенклатуре», представленной коротко в «Филокодексе» (’PhyloCode’), при наименовании организмов ориентируются исключительно на филогенетическое родство.
Таблица 11.1. Иерархический перечень основных таксономических категорий, их нормативные окончания, а также таксономическое положение в системе тысячелистника обыкновенного (Achillea millefolium)
Таксономический ранг (русское и латинское название) |
Окончания |
Таксономическое положение (примеры, синонимы) |
Царство (regnum) |
Eucarya |
|
Подцарство (subregnum) |
-bionta |
Chlorobionta |
Отдел, или тип (divisio/ phylum) |
-phyta, -mycota |
Streptophyta |
Подтип (subphylum) |
-phytina, mycotina |
Spermatophytina |
Класс (classis) |
-phyceae, -mycetes или -opsida (или -atae) |
Magnoliopsida |
Подкласс (subclassis) |
-idae |
Rosidae |
Надпорядок (superordo) |
-anae |
— |
Порядок (ordo) |
-aies |
Asterales |
Семейство (familia) |
-aceae |
Asteraceae (= Compositae) |
Подсемейство (subfamilia) |
-oideae |
Asteroideae |
Триба (tribus) |
-eae |
Anthemideae |
Род (genus) |
Achillea |
|
Секция (sectio) |
Achillea sect. Achillea |
|
Серия (series) |
— |
|
Группа видов (aggregatum) |
Achillea millefolium agg. |
|
Вид (species) |
Achillea millefolium |
|
Подвид (subspecies) |
A. m. subsp. sudetica |
|
Вариация (varietas) |
— |
|
Форма (forma) |
A. m. subsp. s. f. rosea |