БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
13. РАСТЕНИЯ В ЖИЗНЕННОМ ПРОСТРАНСТВЕ
13.3. Устойчивость к температурам
13.3.1. Морозоустойчивость
Среди климатических факторов, определяющих распределение растений по поверхности Земли, самые важные — это снабжение водой и устойчивость к низким экстремальным температурам. Мороз — первый «фильтр окружающей среды», через который должен пройти вид растения, прежде чем он может закрепиться в какой- либо области, подверженной морозам. Те растения, которые прошли этот отбор, устойчивы, т. е. им ничего существенного в общем не угрожает (естественная флора). Растения, побеги которых недостаточно устойчивы, могут переживать критическое время в виде семян (однолетники) или в том случае, когда их органы возобновления находятся в почве (например, геофиты). Эти способы «избегания» обозначаются часто как стратегия «уклонения» (англ. escape) от воздействия мороза (см. рис. 12.2). Морозоустойчивостью считается во всяком случае предотвращение образования смертельно опасных кристалликов льда в цитоплазме. Чтобы понять механизм морозоустойчивости, нужно вспомнить, что вода в растительной ткани находится в двух компартментах: за пределами мембраны протопласта в так называемом апопласте и в ксилеме, прежде всего внутри клеток, возможно в межклетниках (с очень слабо растворимыми субстанциями), симпласте (со значениями осмотического потенциала в тур1есцентном состоянии в интервате от -1,2 до -2,5 МПа).
Существуют два механизма морозоустойчивости.
Предотвращение замерзания, или устойчивость к переохлаждению (англ. super cooling), — это стойкое препятствование к образованию льда при минусовых температурах. В листьях многих растений эти значения достигают -5 °С, некоторых высокогорных растений — до -12 °С, а в ксилемной паренхиме некоторых древесных растений умеренной зоны — примерно до -40 °С. Решающими факторами для замедления замерзания являются отсутствие центров кристаллизации и переход воды в метастабильное состояние. Если критическая температура превышается, происходит моментальное замерзание ткани, что смертельно для клеток.
Толерантность к отрицательным температурам функционально представляет собой специальную форму устойчивости к засухе. В тканях начинается образование льда там, где вода имеет наиболее низкое осмотическое давление, именно в апопласте. При этом межклетники заполняются льдом (без повреждения тканей), причем симпласт постепенно отдает воду. Этот процесс предполагает высокую водопроницаемость, следовательно, подвижность плазмалеммы, не повреждающейся при очень низких температурах, генетический признак, который, однако, определяет хорошие акклиматизационные свойства (морозное закаливание; свойства мембранолипидов). В той, или иной степени обезвоживания клеток и толерантности протоплазмы к обезвоживанию играют роль осмотические параметры и прежде всего защитные субстанции, стабилизирующие клеточные мембраны (растворимые углеводы и так называемые стресс-протеины), которые создают единообразный эффект устойчивости как к морозу, так и к засухе.
Если для предотвращения замерзания важно отсутствие зародышей кристаллов, то для толерантности к замерзанию благоприятно некоторое замедление процесса образования льда. Понижение точки замерзания путем аккумуляции осмотического давления имеет значение только в связи с обезвоживанием клеток, индуцированным образованием льда. Сами по себе клетки в состоянии тургора дают очень небольшой эффект, так как, чтобы понизить температуру замерзания на 1,9 К, нужен 1 моль осмотического давления, что соответствует дополнительному осмотическому давлению 2,24 МПа (у большинства растений это соответствовало бы приблизительно удвоению давления).
Особое экологическое и практическое значение имеет то, что морозостойкость растений сильно связана с внешними и внутренними факторами, поэтому какой-либо конкретный вид нельзя характеризовать единой критической температурой. Насколько низкую температуру может фактически выдержать растение при данном потенциале устойчивости, определяют следующие пять факторов воздействия:
✵ обстоятельства акклиматизации (время года, предшествовавшие температурные условия; рис. 13.6);
✵ фаза развития (активная, растущая, несформировавшаяся или молодая ткань менее вынослива, чем сформированная, менее активная, старая);
✵ тип органа или ткани (корни выдерживают понижение температуры во много меньшей степени, чем листья; в зависимости от фазы развития и вида растения камбий может быть более или менее выносливым, чем древесинная паренхима, листовые почки — чем цветочные почки, и т. д.);
✵ обеспечение влагой (растения, требующие постоянного увлажнения, менее выносливы, чем засухоустойчивые);
✵ обеспечение питательными веществами (оптимально обеспеченные питательными веществами растения более выносливы, чем избыточно удобренные или те, которые испытывают недостаток в минеральных веществах).
Рис. 13.6. Годовой ход длины дня и суточных минимальных и максимальных температур (наверху — средняя трехдневная температура для г. Байрейта) и морозоустойчивость однолетней хвои Pinus sylvestris (внизу). Стрелками отмечено первое наступление ночных заморозков. Серые тона слева направо показывают фазы: чувствительности к морозу, морозоустойчивости, полной (максимальной) морозостойкости и фазу снижения морозостойкости весной
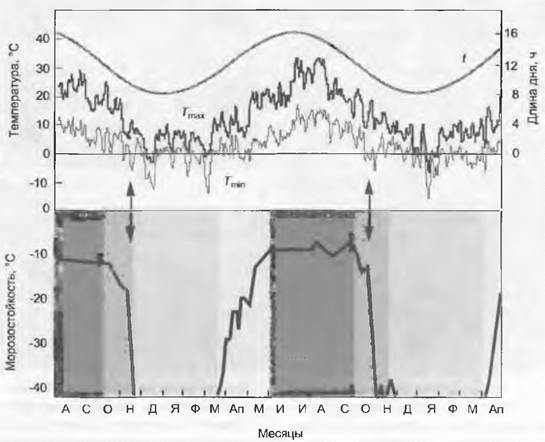
Критическая температура повреждения листовой ткани в умеренной зоне в течение вегетационного периода лежит между -2 и -8°С, причем разнотравье более чувствительно, чем С3-злаки и осоки, а листья летне-зеленых растений чувствительнее, чем вечнозеленых деревьев. Самые опасные ситуации — заморозки поздней весной после распускания почек и вообще экстремальные колебания температуры (резкое падение температуры после предшествующей мягкой погоды). У местных, типичных для данного местообитания растений в первую очередь повреждаются морозом цветы и листья, реже камбий, но это никогда не угрожает жизни всего растения. Особенно опасно радиационное выхолаживание в ясные ночи после прохождения фронта циклона. Максимальная толерантность к зимним морозам наземных органов растений умеренной зоны (при хорошей закалке) лежит между -25 и -40 °С, для взрослых древесных растений средиземноморской зоны — между -10 и -14°С (для растений, выращиваемых в кадках, критическая температура может быть -2 °С). Чувствительные виды переносят морозы под зашитой снега и благодаря подземному положению почек возобновления. Некоторые тропические растения повреждаются уже при температурах 0 — 7°С; тогда говорят о повреждениях от переохлаждения (англ. chilling).
13.3.2. Устойчивость к перегреву
Устойчивость к перегреву варьирует значительно меньше, у высших растений критические температуры лежат в пределах 50 — 55 °С (высокие значения температуры у розеткообразующих, стелющихся, склерофильных растений и суккулентов; максимальные значения, 60 °С, у некоторых С4-злаков, пальм в саваннах, кактусов). Повреждения от перегрева определяются в значительной степени морфологией, расстоянием от почвы, самозатенением основания побега и водоснабжением. В особенно опасном положении находятся всходы на сухих, темных, голых почвах, поверхность которых при сильном солнечном облучении может нагреваться более чем до 75 °С. В таких местообитаниях растениям необходимы быстрое развитие всходов в прохладное время года и последующее затенение почвы своими листьями или другими растениями. Многие виды могут переносить экстремально жаркие ситуации с помощью транспирационного охлаждения (до 10 К; см. 13.1.2). Если корни не достигают достаточных запасов почвенной воды (не редки корни, достигающие 30 м в глубину; см. 13.7.5.1), растения сбрасывают листья или переживают этот период лишь в виде семян.
13.3.3. Экология пожаров
Во многих частях света пожары представляют собой важный экологический фактор для развития экосистем и установления в них характерного набора видов. Крупные биомы приобрели свой характерный облик благодаря пожарам (саванны, семиаридный бушленд, средиземноморская растительность, прерии, а также и бореальные леса). Обычная естественная причина пожаров — молнии. Широкий спектр типичных приспособлений растений к существованию в этих условиях доказывает, что огонь задолго до человека, увеличившего частоту пожаров, был экологически действенным фактором (рис. 13.7). Пирофиты (растения, специализированные к пожарам) часто сохраняют длительное время жизнеспособные банки семян в почве или в кроне, они способны давать пнёвую поросль (расположенные под землей и способные к регенерации стволики, так называемые ксилоподии), снабжены защитной коркой (деревья) или в основном подземной апикальной меристемой (травы, геофиты). У дерновинных трав и верхушечно-розеточных растений (например, Xanthorrhoea, Yucca, Espeletia) защитой от огня часто служат «соломенная туника» или отмершие основания листьев. Феноритмика нередко тесно связана с наступлением пожаров (например, сбрасывание листьев в критическое время засухи). У многих видов Pinus, Eucalyptus, семейства протейных и других плоды раскрываются только после воздействия огня; лишь тогда семена достигают полной всхожести и рассеиваются. Этим обеспечивается возобновление в благоприятный момент, когда снижается конкуренция за свет и корневая конкуренция, а затрудняющий прорастание опад превращается в богатую питательными веществами золу.
Рис. 13.7. Реагирование растений на пожар. Защита от огня: В — толстой коркой (Pinus halepensis, Западное Средиземноморье); D — плотной «юбкой» из отмерших оснований листьев (Xanthorrhoea sp., Западная Австралия, на заднем плане справа); Е, G — погружением меристемы под землю, как у большинства злаков (G — сразу после пожара; Е — 10 дней спустя). Регенерация после пожара: D — за счет семенного банка в кроне и растрескивания одревесневших плодов только под действием высокой температуры (Hakea sp.); F— путем быстрого прорастания в обогащенном питательными веществами и незатененном слое (Eucalyptus sp., восточная часть Австралии); А, С — за счет подземной (С — Arbutusandrachne, Восточное Средиземноморье) или надземной (А — Pinus canariensis, Тенериф) поросли. Н — низовые пожары, которые только минерализуют слой подстилки, лишь слегка повреждают растительность и для обогащения экосистемы питательными веществами особенно благоприятны

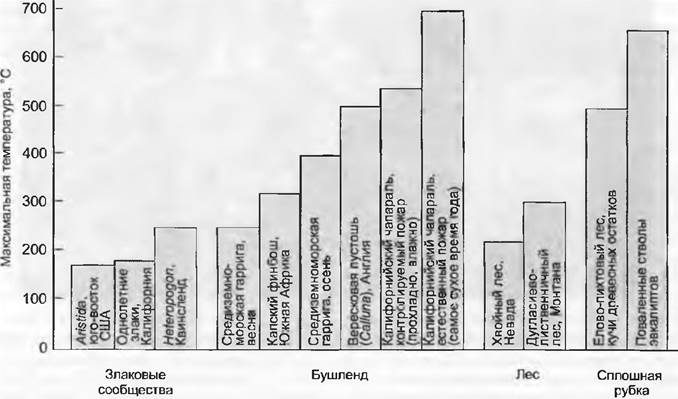
Обязательные циклы возгораний варьируют, они бывают ежегодными (саванна), малолетними (прочие травяные биомы), 30-, 40-летними (средиземноморский буш) и более чем столетними (до нескольких сот лет) в бореальных лесах. Возгоранию способствует легковоспламеняющийся мощный слой подстилки. В сухих редкостойных лесах и саваннах возникают, как правило, не разрушительные верховые пожары (с температурой до 1 000 °С и уничтожением всех древесных растений), а быстро проходящие низовые пожары (температура в слое подстилки и верхнем почвенном горизонте короткое время держится на 70 — 100°С, на высоте 0,5 — 1,0 м едва ли больше 500 °С; рис. 13.8). При этом органы возобновления древесных и травянистых растений повреждаются мало. То же самое можно сказать о пожарах в степях и тропических злаковых сообществах. Решающим фактором оказывается средняя продолжительность прохождения фронта огня по одному месту. При низовом пожаре она часто не достигает 2 мин, что недостаточно для того, чтобы летальные ожоги достигли чувствительной меристемы.
Рис. 13.8. Максимальные температуры поверхности почвы в различных типах растительности при прохождении фронта огня. Продолжительность пребывания пожара на одном месте и при этом наибольшие температуры повышаются с увеличением участия древесных растений. При сухости максимальное воздействие жара также возрастает
Для того чтобы предотвратить опасные пожары вблизи населенных пунктов, часто заблаговременно при прохладной и влажной погоде устраивают контролируемые пожары (например, «традиционное выжигание» в Калифорнии). Также и при управлении природными резерватами идут на то, что, зная экологическое значение пожаров, допускают их или даже устраивают вместо того, чтобы с ними бороться. Те экосистемы, в которых не бывает естественных пожаров или пожары бывают очень редко, страдают очень сильно от поджогов (см. 13.6.1). Поскольку у слагающих такие биомы видов нет устойчивости к пожарам, после последних остаются сильно деградировавшие вторичные сообщества. На месте влажных тропических лесов возникают высокотравные злаковые сообщества, чья восприимчивость к пожарам препятствует восстановлению лесов. Частота пожаров, превышающая определенную величину, также приводит к деградации даже приспособленной к пожарам экосистемы.
Значительный возраст крупных лесных деревьев во многих областях Земли представляет собой результат устойчивости к пожарам. Известный пример — Мамонтовы деревья (Sequoiadendron giganteum) в Калифорнии, которые доживают до 2000-летнего возраста исключительно благодаря своей толстой корке (рис. 13.9). Кроме регулирования набора видов и жизненных форм в экосистеме наибольшее значение пожаров состоит в поддержании круговорота минеральных веществ в таких жизненных пространствах, где незначительная влагообеспеченность задерживает процессы разложения или напочвенный ярус все более препятствует поступлению питательных веществ в виде опала для растительности верхних ярусов (например, мхи в бореальном лесу, см. 15.2.14).
Рис. 13.9. Толстая корка гарантирует долгую жизнь (см. на В остатки корки (Во) 15-сантиметровой толщины). Старый 2000-летний ствол Мамонтова дерева — Sequoiadendrongiganteum (А, С — Сьерра-Невада в Калифорнии) демонстрирует успешное существование с пожарами (D — пожар в эвкалиптовом лесу). Жар подавляется в бедной энергией пластинчатой структуре корки Sequoiadendron (Е), как в стопке газетной бумаги. Черные стрелки (В) указывают на обугленные и снова заросшие места, белые стрелки — на места, по времени совпадающие с соответствующими историческими событиями (1 — коронование Карла Великого, 2 — Колумб в Америке, 3 — конец Первой мировой войны)
