БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
13. РАСТЕНИЯ В ЖИЗНЕННОМ ПРОСТРАНСТВЕ
13.2. Свет как сигнал
В этом разделе речь пойдет о качественном, а именно о сигнальном воздействии света (основы изложены в гл. 7, количественное воздействие света см. 13.7.1).
13.2.1. Фотопериодизм и сезонность
Растения сезонного климата получают через свою систему фитохромов (см. 7.7.2) очень точную календарную информацию, с помощью которой управляются основные этапы их развития (фотопериодизм). Экологические последствия этого многообразны. В тех областях, где есть опасность морозов, растения от поздних и ранних заморозков оберегает фотопериодическая чувствительность, смещая их распускание и отмирание независимо от температуры в данный момент времени в более «безопасные» периоды. В регионах с сильновыраженной сезонностью в отношении осадков (например, в муссонной области) фотопериодическая чувствительность гарантирует цветение и плодоношение, даже если из-за запоздавших или небольших осадков растения будут низкорослыми (индукция цветения, несмотря на субоптимальное вегетативное развитие), на чем основана известная стабильность урожая фотопериодически чувствительных индийских сортов риса в противоположность фотопериодически нечувствительным японским сортам.
Фотопериодизм сильно дифференцирован экотипически (генетически), что доказывают эксперименты по пересаживанию растений в области с другими широтами или высотами над уровнем моря. Арктические растения с большим трудом зацветают в умеренных широтах, растения же умеренных широт в приполярных районах зацветают раньше и сбрасывают листья позже, они «дожидаются» более коротких дней. Проростки деревьев, взятые на верхней границе леса и пересаженные в долины, несмотря на более благоприятные температурные условия, остаются в состоянии зимнего покоя до июня, напротив, пересадка лесных растений с низких высот на верхнюю границу леса также безуспешна, потому что их фотопериодизм не согласован с местными температурными условиями. Это важные барьеры к возможности использования более длинных вегетационных периодов в случае потепления климата.
На примере травянистых неофитов (пришельцев из других мест) наблюдалось, что должно смениться по меньшей мере 6 поколений, прежде чем сформируются новые, приспособленные к местному фотопериоду генотипы (экотипы); если речь идет о деревьях, то это составляет значительное время. Кроме уже известных влияний на ход цветения (см. 7.7.2.2) наблюдались также многочисленные воздействия на морфологию и физиологию обмена веществ. Так, например, Роаpratensis из Северной Скандинавии, по О. М. Хайде, при симулировании длинного дня путем облучения красным светом образует больше листьев, но при этом сами они тоньше (большее SLA, см. 13.7.3); это способствует увеличению их роста и продукции биомассы по сравнению с растениями с той же дозой дневного света, но без удлинения фотопериода красным светом, хотя интенсивность фотосинтеза на единицу листовой поверхности даже снижается.
Считается основным правило о том, что регулирование фотопериодизма при переходе осенью к старению и повышению резистентности протекает точнее и в мень
шей степени зависит от погоды, чем при выходе из зимнего покоя весной. Многие горные растения весной являются «оппортунистами», а осенью «упрямцами», что обеспечивает обратный перенос мобильных ресурсов из листьев до того, как они будут повреждены морозом.
13.2.2. Сигнал красного света в растительных сообществах
Излучение, которое проникает в зеленые листья или отражается от «зелени», богаче темно-красной областью спектра (700 — 800 нм) и беднее ярко-красной (620 — 680 нм), т. е. отношение более коротких красных волн к длинным (например, I666/I730) уменьшается (от англ. red/ farred ratio — R/FR; К/ИК). Тот факт, что растения с их чувствительностью к красному свету (фитохром, см. 7.7.2.4) могут как-то определять свое положение по отношению к соседям, имеет далеко идущие последствия для формирования растительных сообществ и конкуренции между растениями. На отдельных модельных растениях было показано, что их побеги избегают уже занятых «зеленых» мест и ресурсы оптимизируются в новых побегах.
А. Новопански с соавторами помещали перед стелющимися, растущими в нескольких направлениях молодыми побегами Portutaca olerácea зеленые и серые карточки, окружая таким образом растение. Верхушки побегов искали путь в направлении серых карточек в сторону от зеленых, абсорбирующих ярко-красный свет. Также и рост в высоту конкурирующих всходов, до тех пор, пока не начнет действовать обоюдное затенение, стимулируется очень слабым перемещением красных волн, идущих от соседних растений. С.Баллар с сотрудниками экранировали побеги Datura ferox и Sinapis alba фильтрами красного света, вследствие чего они становились «слепыми», их соседи больше не воспринимались, и по сравнению с контрольными растениями эти побеги не вытягивались в длину. Можно предположить, что подобные механизмы составляют сенсорику многих растений наряду с «нормальным» фототропизмом.
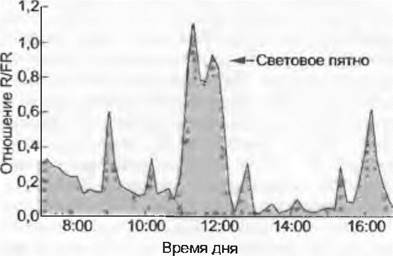
На семенное возобновление в густых растительных насаждениях смещение красных частей спектра от ярко-красной к более темной оказывает столь сильное влияние, что прорастание под сомкнутыми кронами, несмотря на отчасти уже имевшее место набухание семян, происходит лишь тогда, когда «окно» в кронах (сигнал яркокрасного цвета) укажет на возможность фотосинтетической деятельности, т. е. выживание. Таким образом, возобновление лесов семенным способом сильно зависит от динамики «окон» в насаждениях (англ. gap dynamics). Густые сомкнутые кроны снижают отношение R/FR независимо от интенсивности излучения от 1,2 при прямом солнечном свете до 0,2 под пологом (рис. 13.5). Итак, определение отношения R/FR является дополнительным непрямым методом для количественной оценки сомкнутости крон (LAI, см. 13.1.3).
Рис. 13.5. Дневной ход значений отношения R/FR в напочвенном ярусе субтропического леса в северо-восточной части Австралии (по С.Turnbull и D. J.Yates, из R.Chazdon и др.). R/FRна лесной почве минимально, когда остаточное освещение, проникающее сквозь кроны, еще и загораживается широкими листьями подлеска, и оно максимально, когда световое пятно (прямой солнечный луч), проникающее через просвет в кронах, достигает датчика. Отношение над насаждением в целом около 1,2
Смещение красных спектров света хлорофиллом проявляется и в отражаемом световом потоке от растительного покрова в целом, что может быть использовано для определения проективного покрытия с применением дистанционного зондирования (англ. remote sensing). Так называемый индекс листовой поверхности растительности NDVI (normalizeddifferential vegetation index) можно оценить поданным аэро- или космической съемки. NDVI основан на измерениях излучения в ярко-красном (IR, 660 нм) и инфракрасном (IFR, 730 нм) диапазонах спектра:
NDVI = IFR- IR /(IFR+ IR).
Чем плотнее почва покрыта фотосинтетически активными структурами, тем больше снижается отражение волн ярко-красной части спектра и увеличивается отражение в темнокрасной (инфракрасной) части. Имея такие данные, можно заблаговременно оценить потенциальный урожай на очень больших площадях (плохая перезимовка озимых хлебов весной заметна из космоса). Также с помощью данных этого «тематического сканирования» можно установить в общих чертах изменения в землепользовании или естественной растительности. На удивление надежна даже оценка продуктивности из космоса.