БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
13. РАСТЕНИЯ В ЖИЗНЕННОМ ПРОСТРАНСТВЕ
Обеспечение солнечной радиацией, снабжение водой и минеральными веществами, а также синтез углерода, необходимый для роста и продуцирования биомассы, — важнейшие связующие звенья между растениями и физико-химическими свойствами окружающей их среды. Биохимические и физиологические основы этой связи были обсуждены в разделе 6 (частично в разделах 7 и 9). В данном разделе рассматриваются реакции отдельных растений, растительных сообществ и экосистем на естественные колебания солнечного излучения, запасов влаги и питательных веществ, а также баланс углерода в его экологическом преломлении. В заключение обсуждаются биологические взаимодействия и влияние человека на растительность и использование им растений.
13.1. Радиация и бюджет энергии
Режим излучения и бюджет энергии определяются климатом как напрямую, так и опосредованно через режим испарения, образования облаков и осадки, а также местные или глобальные влияния на температуры. При полном полуденном солнце летом в Центральной Европе на земную поверхность попадает энергия в количестве до 900 Вт на 1 м2. Происходящее с этим огромным количеством энергии в экосистеме, да и на каждом отдельном листе имеет решающее значение для понимания жизни растений.
13.1.1. Количество радиации и радиационный баланс
Общее количество излучения солнечной энергии на земную поверхность называют суммарной (глобальной) радиацией. Она подразделяется на прямую и рассеянную и благодаря отражению и поглощению атмосферой в средних широтах составляет только около половины интенсивности излучения, измеренного в направлении Солнца за пределами атмосферы («солярная константа» = около 1400 Вт м2). Около половины суммарной радиации приходится на область видимого излучения (длина световой волны 380 — 780 нм), что в значительной степени соответствует спектральной области фотосинтетически активного излучения в области длин волн от 380 до 710 нм (от англ. photosynthetic active radiation — PAR, PhAR — в основном характерна для области с длинами волн 400 — 700 нм; Вт м2). Так как фотосинтетически активное излучение (см. 6.4.1, бокс 6.2) состоит из фотонов (частиц света), имеющих различную энергию (нагрузку квантов: синие — энергетически богатые, красные — энергетически бедные), нейтральными нефильтрующими сенсорами излучения в первую очередь будут всегда улавливаться коротковолновые, богатые энергией спектральные области. Так называемые фотонные датчики фильтруют свет, и все фотоны в области между 400 и 700 нм регистрируются примерно равноценно и таким образом становятся фотонными счетчиками. Благодаря прямым стехиометрическим связям между абсорбированными фотонами в области спектра 400 — 700 нм и фотосинтетической фиксации СО2, в биологии за стандарт принята плотность фотонного потока (от англ. photosynthetically active photon flux density — PPFD, или сокращенно PFD, обычно измеряемая в мкмоль фотонов на 1 м 2 в 1 с-1 иногда используется единица «Эйнштейн», 1 Е = 1 моль фотонов, что не согласуется с СИ). Широко распространено, но некорректно данные датчиков фотонного потока (неправильно называемых квантовыми датчиками) обозначать в мкмоль • м 2 • с-1 как PAR, PhAR (см. выше). Наоборот, для расчета энергетического баланса энергии непригодна PFD, так как плотность фотонного потока не является единицей энергии.
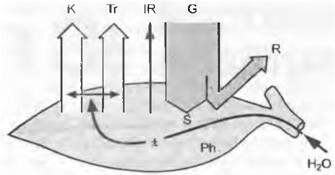
Диффузные компоненты суммарной радиации проникают в растительные сообщества существенно лучше (глубже), чем прямое излучение, создающее резкие тени. Растения в зависимости от формы и размера листьев могут повышать долю диффузной радиации в сообществе (например, хвоя или тонкие перистые листья акаций увеличивают долю рассеянного света). Часть суммарной радиации будет отражаться от поверхности, на которую она падает, и на это отражение растительный покров оказывает существенное влияние. Отражение от пустынных кустарников со светлыми листьями составляет около 20 % радиации, от елового леса — лишь 10 %, от голой почвы — до 30%, а от свежевыпавшего снега — до 80 % радиации. Оставшаяся часть, радиационный баланс, представляет собой то количество энергии, которое абсорбируется листьями и, соответственно, сообществом (рис. 13.1). Днем это количество всегда имеет положительное значение, ночью — нулевое или отрицательное. Отрицательный ночной радиационный баланс возникает благодаря собственному термическому излучению.
Рис. 13.1. Энергетический баланс листа. За вычетом отраженной (R), излученной (IR) и использованной для фотохимических процессов (Рh) энергии большая часть падающей суммарной радиации (G) энергия радиационного баланса (S), должна быть отдана листом, так как его масса слишком мала, чтобы аккумулировать существенную энергию. В зависимости от водоснабжения она может либо расходоваться на теплоту испарения (транспирацию, Тг, «неощутимый поток тепла»), либо уходить в воздух через конвективную отдачу (К, «ощутимый поток тепла»)
Все тела (даже газы) излучают термическую энергию пропорционально четвертой степени их абсолютной температуры. Баланс этого теплового излучения зависит от температуры находящихся напротив или окружающих тел. Теплое тело в холодном окружении излучает больше, чем воспринимает. Для растений при ясном небе в светлую ночь потеря теплоты путем излучения в холодное окружающее пространство значительна, и температура листьев охлаждается на 3 — 5 К по сравнению с воздухом, что может привести к их переохлаждению (К, Кельвин, для разницы температур независимо от «градусной» шкалы). Облачность или туман препятствуют этому эффекту.
13.1.2. Баланс энергии и микроклимат
Лист может «избавиться от забот» о полученной через радиационный баланс и адсорбированной самим листом энергии с помощью четырех способов: (1) термическое излучение; (2) фотохимическое связывание энергии (максимум 1 — 2 % от общей PAR); (3) транспирация воды Тг; (4) отдача энергии через тепловую конвекцию К (отток ее через нагретый окружающий воздух). Днем только последние два компонента имеют значение для энергетического баланса листа Q (аккумуляция теплоты не имеет значения из-за малой массы листьев):
Q = Тг + К; Тг = g∆wv; К = h∆Tq,
где g— способность к диффузной проводимости листового эпидермиса для газообразной воды (по сути, способность к проводимости устьичных отверстий); ∆w — градиент молярных соотношений водяного пара и воздуха между внутренними тканями листа и окружающим воздухом (на уровне моря при давлении воздуха 0,1 МПа численно соответствует градиенту давления пара); v — теплота испарения волы (при 20 °С равна 2,45 кДж/г); h — способность поверхностных слоев листа отдавать теплоту в воздух (функция ширины листа и скорости ветра); ∆Т — разница между температурами листа и воздуха; q — теплоемкость воздуха. Проводимости — обратные величины соответствующих сопротивлений (сопротивление диффузии водяного пара и сопротивление отдаче теплоты).
Посредством g и h растения с помощью своих листьев оказывают физиологическое и морфологическое воздействие на свой собственный климат, а также на климат окружающей среды, при этом они сами зависят от водоснабжения. Охлаждение может отнимать энергию только при более высокой влажности почвы («латентный» тепловой поток; температура самого листа остается близкой к температуре окружающего воздуха или ниже ее на 1 — 2 К). При недостатке влаги и закрытых устьицах поток энергии идет в принудительном порядке к тепловой конвекции («ощутимый поток теплоты»), что может привести к их гибели от перегревания, если отсутствуют морфологические адаптации, облегчающие теплоотдачу Растениям более жарких и сухих местообитаний часто свойственны листья, перпендикулярно направленные к солнечным лучам, мелкие и хорошо отражающие свет. Этим достигаются снижение абсорбции излучения и хорошая термическая связь с воздухом (низкое аэродинамическое сопротивление пограничных слоев) и избегается перегрев. Зная величину, Q, влажность и температуру воздуха, скорость ветра (метеорологические показатели), а также величину g и ширину листа, можно рассчитать его температуру.
В растительных сообществах для газового и теплового обмена добавляются еще и аэродинамические препятствия. Чем гуще и ниже сообщество, тем сильнее его независимость от атмосферных условий и поэтому тем больше оно удерживает теплоту и влагу. В наибольшей степени это проявляется у прижатых к земле горных растений (особенно подушек), в листовом ярусе которых при ярком солнце могут создаваться условия, близкие к влажным тропикам, что не имеет ничего общего с данными какой-либо метеостанции. При таких эффектах сомкнутости прямое влияние устьиц на транспирацию уменьшается. Структура сообщества сама становится определяющей воздействующей величиной.
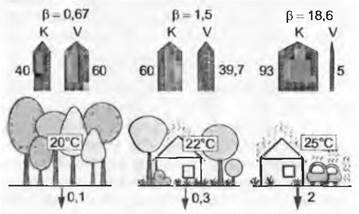
В целом аналогичные связи имеют значение и для энергетического баланса всей экосистемы. Роль транспирации листьев и всего сообщества выполняет эвапотранспирация ЕТ (или, иначе, общее испарение V, включая эвапорацию почвы и ее увлажненных поверхностей; причем в сомкнутых сообществах, а также при увлажненной поверхности почвы вклад транспирации листьев составляет более 80 %). При повышенном испарении экосистема остается сравнительно холодной, при пониженном нагревается. Эквивалент энергии, выраженный отношением K : V, называют отношением Боуэна, β (англ. Bowen ratio). Считается, что, если β меньше 1, растительность не испытывает недостатка во влаге. В условиях сухости или при наличии почвенной корки β увеличивается в направлении бесконечности, т. е., когда вся вода используется, энергия должна почти целиком уходить на нагревание воздуха (небольшая ее часть временно аккумулируется в почве в виде теплоты; рис. 13.2). Значение β можно бесконтактно определить метеорологическими методами (измерение радиационною баланса и вертикальных климатических градиентов над растительным сообществом).
Рис. 13.2. Влияние транспирирующих растений на окружающую температуру. В качестве примера представлено три ситуации, в которых из-за редуцирования зеленого покрова все большая часть лучистой солнечной энергии должна «отторгаться» в виде тепловой конвекции (К). При сомкнутом растительном покрове и влажной почве более половины энергии через теплопотребление затрачивается на испарение воды (V) в «холодном» состоянии, воздух остается прохладным, так называемое отношение Боуэна: β = К/V1<1, и тепловым потоком от почвы можно пренебречь. С возрастанием уплотнения почвы вследствие преобладания нетранспирирующих поверхностей значение К увеличивается, β становится >1, воздух и почва нагреваются сильнее. Рисунок наглядно показывает, почему воздух в городских зеленых насаждениях холоднее, чем в застроенных, «опечатанных» районах (численные значения даны в % от излучаемой энергии)
Поверхность почвы после дождя высыхает за несколько дней, причем испарение с самой почвы очень небольшое. После освоения глубоких слоев почвы корнями (см. табл. 13.3 в разделе 13.7.5.1) испаряющаяся вода попадает в атмосферу, что невозможно без участия растений. Растения связывают таким образом глубоко залегающие запасы воды с атмосферой и посредством устьиц сохраняют контроль над данным процессом (см. 13.5.2). Эти закономерности объясняют тот факт, почему скверы и парки в городах представляют собой прохладные островки и почему после сведения лесов температура повышается так, что может даже изменить климат на больших площадях (теплые восходящие потоки воздуха снижают количество осадков, рис. 13.3). Зеленый растительный покров влияет как на водный (см. 13.5), так и на энергетический баланс в ландшафте. Благодаря своей морфологии и регуляции транспирации растения влияют на собственный климат и на климат экосистемы.
Рис. 13.3. Региональные климатические последствия сведения лесов. Между левым и правым изображениями лежит примерно 400 летняя история. Первооткрыватели 16-го столетия описывали эти места как зеленые джунгли (А — близ Валенсии); в наше время в этой части Венесуэлы в результате сведения лесов, перевыпаса скота, деградации почв и повторяющихся пожаров господствуют колючие кустарники (B — близ Баркисимето). Прогрессирующая ликвидация транспирационного охлаждения (см. рис. 13.2) привела к региональному потеплению и семиаридному климату. Отношение Боуэна (β), равное примерно 1, держит экосистему слева сравнительно прохладной (<30 °С), а равное значительно больше 1, допускает повышение температуры в системе справа свыше 40 °С, что само по себе вызывает сильный восходящий поток тепла и приводит к снижению осадков

Пример действия транспирации растений, регулирующего климат, может дать изучение А. Х. Розенфельдом и Дж. Дж. Роммом городского климата Лос-Анджслеса. Под воздействием непрерывного роста городской агломерации и связанного с этим уплотнения почв температура воздуха в черте города повышается в среднем на 1 К все 15 лет. Если бы между домами было посажено больше тенистых деревьев (трапспирационное охлаждение) и крыши были окрашены в более светлый цвет (отражение), можно было бы сэкономить в гол 0.5 млрд. долларов, расходуемых на охлаждение и очистку воздуха от смога. Во всех городах южной части США эффект составлял бы по приблизительным подсчетам 5 — 10 млрд. долларов в год, даже не считая улучшения качества жизни в столь зеленом городе. Благодаря расходу энергии на транспирацию растения, не считая затенения, действуют еще и как охлаждающее климатическое устройство.
13.1.3 Свет в растительном сообществе
При проникновении потока фотонов через крону растения или растительного сообщества так же, как и через толщу воды (см. рис. 12.10), его плотность последовательно ослабляется. Масштабы этого ослабления определяют индекс листовой поверхности (LAI) насаждения, поскольку листья, которые получают количество света, недостаточное для положительного углеродного баланса, отторгаются от растения (или в таких местах совсем не образуются). Аналогичным образом в водных толщах фотосинтетически активный планктон распределяется по глубинам. Для высших растений граница чистой прибыли от фотосинтеза у адаптированного к слабой освещенности листа находится на уровне примерно 0,2% от максимальной полуденной плотности фотонного потока (PFD около 3 — 5 мкмоль • м-2 • с-1). Если учесть потери углерода листом в ночное время и потребности в углероде нефотосинтезирующих органов, минимальная потребность PFD для положительного углеродного баланса повышается до 0,5 — 1 % от интенсивности, измеренной над сообществом в середине дня.
Для гомогенного насаждения в фотометрии по аналогии с законом поглощения Ламберта — Бэра действительно экспоненциальное отношение (рис. 13.4) в формуле ![]() где I и Iо — PFD под рассматриваемым ярусом сообщества и над ним; к — коэффициент поглощения.
где I и Iо — PFD под рассматриваемым ярусом сообщества и над ним; к — коэффициент поглощения.
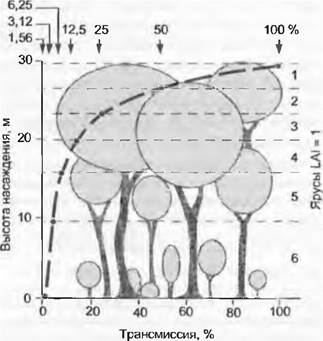
Рис. 13.4. Типичная картина поглощения света в лесу. Для упрощения принято, что в каждом из 6 ярусов насаждения листовая поверхность равна 1 м2 на 1 м2 поверхности почвы (индекс листовой поверхности LAI = 6). Листовая поверхность распределена гомогенно, и в каждом ярусе хальбирует трансмиссионный поток фотонов (коэффициент поглощения к = 0,69). Экспоненциальное ослабление PFD, согласно закону Ламберта- Бэра, определяется тем, с какого количества таких листовых ярусов (с какого суммарного значения LAI) достигается критическая граница для положительного углеродного баланса в самом нижнем ярусе насаждения (минимальный PFD, необходимый для положительного углеродного баланса одного листа). Максимально возможное значение LAI определяется преимущественно коэффициентом поглощения, который, как правило, находится в пределах от 0,4 до 0,8
Величина варьирующего коэффициента поглощения зависит от средней величины листьев и от угла их отхождения, а также немного от листовой трансмиссии и значительно от положения Солнца и участия диффузного излучения. Типичные значения к составляют 0,4 — 0,5 для насаждений с листьями, расположенными в вертикальной плоскости или с очень мелкими листьями (например, граминоиды, хвойные; мелкие листья способствуют сильному рассеянию света), и 0,7 — 0,8 для расположенных в горизонтальной плоскости крупных листьев (например, некоторые представители высокотравен и очень широколистные деревья). Для луговых сообществ к равен примерно 0,5, для лиственных лесов умеренного пояса — 0,65. Можно определить значения к и среднего угла отхождения листьев исходя из разницы ослабления света под различными азимутальными углами (компьютерный анализ распределения световых пятен на фотоизображениях, полученных с помощью направленного вверх объектива типа «рыбий глаз»).
Зная величину к и I0 (последнее — по данным метеостанции), можно спрогнозировать величину I для определенного значения LAI и таким образом дать приблизительную для этого местоположения величину плотности потока фотонов на лист. Зная величину I, I0 и к, получают значение и для LAI. В настоящее время техника, которая с помощью концентрически сегментированных «рыбий глаз»-сенсоров и сложных алгоритмов сама определяет значения к, требует для определения LAI только двух операций, по возможности синхронных: замеров излучения под пологом насаждения и над ним, причем может очень быстро и не нарушая структуры выяснить один из важнейших биологических параметров растительного сообщества, если выполнены «крайние условия», прежде всего распределение листьев — гомогенное и случайное.
Типичные значения LAI для сосновых лесов — 3 — 4; для смешанных лиственных лесов умеренного пояса — около 5,5; для высокопродуктивных сенокосных лугов — 7 — 8; для всех ярусов равнинных влажно-тропических лесов — 8; для густых еловых посадок — 10. В сомкнутых посевах сельскохозяйственных культур к моменту максимального развития вегетативных побегов величина LAI в зависимости от сорта равна примерно 4, в естественных лужайках высокогорий — около 2. Значения LAI свыше 10, иногда упоминающиеся в литературе, нереальны. В лесах при таких расчетах к LAI добавляется также ослабление светового потока стволами и ветвями деревьев, поэтому нередко используется понятие PAI(англ. plant area index), причем вклад таких нелистовых структур в общее значение обычно не достигает I. Калибровка этих измерений требует прямого (деструктивного) определения листовой поверхности (урожай). Простой и очень поучительный (даже если и ретроспективный) метод приблизительного определения LAI — это «накалывание» листьев в лиственных лесах вскоре после осеннего листопада, лучше всего при влажной погоде. Среднее число проколов листьев иглой сквозь свежий слой опавших листьев непосредственно показывает величину LA1 перед листопадом, если до этого не было сильного переноса их ветром. Как правило, результаты этого метода на удивление хорошо совпадают с расчетными методами поглощения.
Путем определения значений LAI в разных ярусах сообщества для многих из них можно выявить очень характерное вертикальное распределение листовых поверхностей и тем самым типичную кривую абсорбции света в насаждении. В лесах максимум абсорбции часто выражен в верхней части крон деревьев (до половины PFD абсорбируется в самом верхнем м2 листовой поверхности на I м2 поверхности почвы), в то время как в луговых травостоях абсорбция большей частью происходит в нижних ярусах.
Этому способствует и распределение углов отхождения листьев по ярусам: вертикально ориентированные листья в верхних ярусах травостоя, горизонтально — в припочвенных ярусах (например, розеткообразующие травы). Фактическое световое довольствие листа падает вместе с косинусом угла паления излучения к листовой нормали (закон косинуса). Между углом отхождения листа и его анатомией имеется тесная связь. Чем вертикатльныe ориентирован лист, тем в большей степени его мезофилл расположен симметрично по отношению к сторонам листа, т. е. в узких, почти вертикально ориентированных листьях часто исчезает дифференциация между палисадной и губчатой паренхимой. В сельском хозяйстве и садоводстве световое довольствие оптимизируется путем создания смешанных культур (например, огурцы или тыква с кукурузой). Угол отхождения лист и, соответственно, геометрический компонент абсорбции излучения ярусом насаждения играют большую роль в оптимизации урожая, и выбор техники возделывания и ухода связан с обеспечением высокого значения LAI. Вертикальная ориентация листа (например, у японских сортов риса) обеспечивает высокие значения LAI в тропиках, что благоприятно влияет на увеличение урожая при азотном удобрении. Более горизонтально ориентированные, а у злаков повисающие листья (например, у индийских сортов риса) быстро приводят к самозатенению, повышение значений LAI посредством удобрений не обеспечивает увеличение урожая. Даже в ярко выраженных «солнечных» листьях для фотосинтеза, как правило, достаточно меньше половины того полного излучения, которое дает полуденное солнце. Листья наружного слоя насаждения часто подвержены сверхоптимальному излучению. Неоднократно было показано, что вертикальный профиль распределения излучения в растительном сообществе коррелирует с распределением азота и максимальной степенью фотосинтеза Аmах (больше азота и более высокое значение Аmах в верхних ярусах насаждений, см. 13.6.3). Внесение азотных удобрений может повысить Аmах настолько. что весьма тенелюбивые виды, например какао, больше не будут нуждаться в затенении и самые верхние их листья смогут вынести полное освещение тропическим солнцем.