БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
13. РАСТЕНИЯ В ЖИЗНЕННОМ ПРОСТРАНСТВЕ
13.5. Водный режим
Наземное существование растений возможно только при эффективно действующих корнях, несжимающейся капиллярной проводящей системе (ксилема, см. 3.2.4.2), комбинации изменяющихся (устьица) и статичных (кутикула) приспособлений к защите от испарения и вакуолизированных в различной степени тургесцентных клеток (водный потенциал, см. 6.1.4.2. 6.3.2.1). При недостатке воды растение может усиливать как ее потребление, так и способы защиты от потери воды и ее последствий. Поскольку строение и функции компонентов, обеспечивающих регуляцию водного режима, были описаны выше, речь пойдет о реакциях на дефицит влаги и будет показано, как согласованно растения управляют водным потенциалом, транспортированием водных потоков и механизмами сопротивления транспортированию; при этом они сами влияют на водный режим экосистемы.
13.5.1. Водный потенциал и транспирация
Водный потенциал (см. 6.1.4.2) в определенной точке растения есть результат тока воды и сопротивления ее передвижению. С термодинамической точки зрения он означает пониженную по отношению к свободной воде доступность в точке измерения, что также может быть обозначено и как напряжение, отсасывание или отрицательное давление. Водный потенциал будет тем негативнее, чем в большей степени он падает. Пока он не перейдет критического значения (в листьях в зависимости от вида растения оно составляет от -1,5 до -2,0 МПа), говорят, что водный потенциал лишь немного превышает фактическую степень водоснабжения растения. При достаточной влажности почвы водный потенциал листьев тем ниже (негативнее), чем выше степень транспирации. Уменьшение потенциала с увеличением пропускной способности сравнимо с давлением в водопроводе, которое падает тем больше, чем шире открывают кран. Если в почве преобладает дефицит воды, даже очень незначительная транспирация (и даже полное закрытие устьиц) не может препятствовать падению водного потенциала до очень низких значений. В одинаковой степени низкий водный потенциал в листьях может появиться в зависимости от состояния снабжения водой при очень интенсивной транспирации или при почти полном ее прекращении. Без сведений о синхронном транспирационном потоке нельзя интерпретировать водный потенциал и ответить на вопрос, действительно ли растение испытывает дефицит воды (во всяком случае не при значениях более -2 МПа).
Ключ к оценке снабжения растения водой — диаграмма соотношений потенциала и транспирации (рис. 13.11). Она позволяет на графике, как правило, линейных связей, используя наклонные прямые, маркирующие возрастание гидравлического сопротивления, интерпретировать его величину и определить равновесный потенциал с почвой в ранние утренние часы (англ. predawn water potential) через регрессию потенциала у нулевой точки транспирации. Пронумеровав и соединив точки на диаграмме, отмеченные хронологически в течение одного дня, можно обнаружить, что линия соединения представляет собой поучительный гистерезис (в послеполуденные часы значения возвращаются к нулевой транспирации при более низком уровне водного потенциала по тому же пути, как эти значения «шли» утром при возрастающей транспирации). Это показывает ухудшение гидравлических условий в течение дня, что связано либо с кавитацией ксилемы (закупорка воздухом, газовая эмболия), либо с сопротивлением транспортирования в корневой системе.
Рис. 13.11. Связь между водным потенциалом листьев и степенью транспирации. Как правило, линейной связью описывают сумму всех гидравлических сопротивлений в континууме почва — лист (диагональные прямые а, b, с обозначают постепенно повышающееся сопротивление транслокации). Потенциал при транспирации, равной 0, означает равновесное состояние между растением и почвой в ранние утренние часы (PD, predawn water potential). А — нормальное снижение без гистерезиса; В — гистерезис в течение дня (время нанесено на график), указывающий на увеличивающееся затруднение водного транспорта (кавитация ксилемы или обезвоживание почвы вблизи тонких корней). При влажной почве большая крутизна линии регрессии также отражает патологические изменения в ксилеме или повреждения тонких корней (например, поражение грибами или корневая гниль)
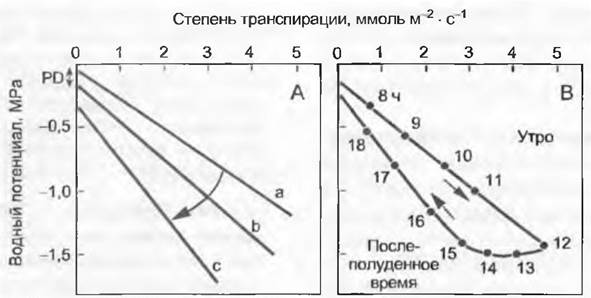
Водный потенциал листьев обычно ниже, чем потенциал в ксилеме осей побегов или ствола. Равновесное давление на срезе побега или черешка листа, полученное методом барокамеры (см. рис. 6.39), есть смешанная величина для всех дистальных тканей, в основном тканей листа, почему и уместно выражение «листовой» или «побеговый» водный потенциал (но не водный потенциал ксилемы). При интенсивной транспирации происходит резкий спад потенциала от оси побега через черешок листа в направлении пластинки.
Между обратным значением водного потенциала и дефицитом влагонасыщенности WSD (см. 6.3.6) существует нелинейная зависимость (кривая давления — объема), которая дает возможность выявить эластичность клеточных стенок, осмотическое давление при полном тургоре, а также водный потенциал в точке потери тургора, поэтому такие характеристические кривые применяют для многих растений открытого грунта. Водный дефицит вообще может возникнуть из-за эластичности клеточных стенок (в «бетонных» клетках не могло бы быть дефицита влаги, так как вода их не растягивает). Эластичность клеточных стенок есть также мера того, насколько прямо (и насколько сильно) всасывающая сила атмосферы при испарении сообщается с корневыми системами почвы. Поэтому в районах с периодическими засухами возрастает число растений с очень жесткими, малоэластичными стенками клеток. Это также одна из причин распространения склерофилии в регионах с сухими периодами.
Крайне важное с экологической точки зрения явление — кавитация ксилемы. Если капилляры при сильной транспирации приходят в состояние напряжения, может наступить прорыв воздуха и образование воздушных пузырьков (газовая эмболия, см. 6.3.5), что быстро приводит к разрыву гидравлического континуума. Так как в первую очередь у деревьев при хорошей погоде кавитация является правилом (акустически ее возникновение обнаруживается в ультразвуковой области), существует подозрение, что кавитация не всегда неблагоприятна для деревьев (возможно, она способствует предохранению от перегрузок). Водный потенциал в листьях вследствие этого опускается быстрее, чем в ксилеме (ограниченный континуум силы сцепления). Есть первые указания на то, что эмболия в позднее послеполуденное время может ликвидироваться точно так же быстро, как и возникнуть в полдень, причем активную роль в этом играют паренхима ксилемы и флоэма. Если удаляют флоэму, «ремонт» ксилемы явственно замедляется. Эти факты лежат в той развивающейся области исследований, в которой тонкий анализ физиологии ксилемы еще может преподнести некие сюрпризы. При этом в центре стоят не дебаты теории сцепления (которая в принципе едва ли оспорима), а изменение кавитации, ее влияние на реальное состояние напряжения в ксилеме, функция пор и древесинной паренхимы, а также то, каким образом ксилема способна быстро ликвидировать кавитацию.
Отчетливые различия в критических значениях водного потенциала, при которых проявляются первые эмболии (ультразвуковые измерения на засыхающих ветвях) у различных видов, могут коррелировать с диаметром проводящих путей (при большом диаметре эмболия более вероятна), что с другой стороны, возможно, находится в связи со специфической глубиной проникновения корней (до достигаемой почвенной влаги) и ритмами сезонного развития (влажный и сухой периоды). В средиземноморских смешанных насаждениях, состоящих из сезонно-зеленых видов с крупными порами и вечнозеленых с мелкими, последние, которые по сравнению с первыми обладают более глубокими корнями, менее (но более длительно) активны и в сухие периоды менее подвержены эмболии. У видов древесных растений, обитающих в сухих местах, эти признаки ксилемы более выражены.
13.5.2. Реакции на дефицит воды
Водный режим растений протекает в зависимости от степени дефицита воды в несколько этапов и включает «предохранительные» реакции. В природе можно различить 6 таких этапов, причем наличие первого и прежде всего второго этапов было «доказано» только для ограниченного числа видов.
Этап 1. Реакция устьиц на степень транспирации (даже при высокой влажности почвы), открытая впервые в 70-е годы XX в. О. Ланге и Р. Лёшем в Вюрцбурге и объяснявшаяся чувствительностью к влажности воздуха. Она изменила картину механики регуляции водного бюджета растения (рис. 13.12), когда обнаружилось, что устьица без всякого стресса (без существенно сниженного водного потенциала) реагируют на высокое напряжение транспирации, как бы профилактически уменьшая ширину щели. Между тем было доказано, что сигналом служит не сама влажность воздуха, а вызываемая ею степень транспирации. Предполагается, что в качестве
возбудителя действует зависящая от транспирации аккумуляция абсцизиновой кислоты (ABA) в апопластах замыкающих и побочных клеток.
Рис. 13.12. Реакции устьиц на влажность воздуха. Многие растения, в особенности многолетние, и при высокой влажности почвы сужают устьичные отверстия (листовая диффузионная проводимость), если воздух становится сухим. Таким образом они замедляют иссушение почвы. Пример такой «feed forward» -реакции основан на Eucalyptus paucifloraиз юго-восточной части Австралии (данные только для значений водного потенциала <1,5 МПа)
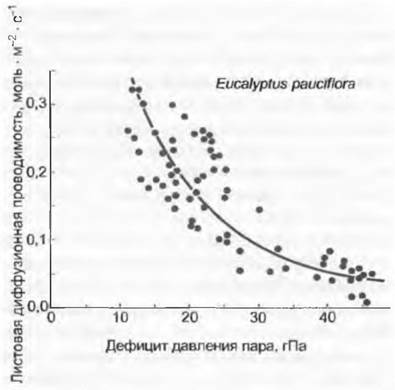
Следствием такого поведения на экосистемном уровне является то, что ко времени достижения хорошего снабжения водой ее резервы в почве не подвергаются чрезмерной нагрузке. Такое скромное потребление воды выгодно естественно только растениям с высокой продолжительностью жизни или растениям, «рассчитывающим» на засуху. Фактически это в первую очередь деревья и среди них особенно долго живущие, а также многолетние пустынные растения, у которых подобное поведение выражено ярче. Критические значения давления водяного пара в воздухе для возбуждения реакции находятся в пределах от 8 гПа (для адаптированных к гумидному климату) до 15 гПа (для адаптированных к аридным условиям). При температуре 20 °С это соответствует относительной влажности воздуха 65, или соответственно 35 %, при 30 °С — 80, или соответственно 65 %. При превышении этого порога способность к диффузионной проводимости устьиц падает вместе с повышением дефицита давления водяного пара (зависимость, близкая к линейной), что определяет поведение устьиц многих растений (прежде всего деревьев) в поздние предполуденные часы. Тем самым тормозится транспирация и стабилизируется водный потенциал. На пустынных растениях было показано, что в пасмурные дни с меньшей сухостью воздуха водный потенциал в листьях благодаря замедленной реакции устьиц падает даже сильнее, чем в ясные дни, когда дефицит давления пара достигает высоких значений и устьица сокращают транспирацию непропорционально резко. Возможно, что такое поведение устьиц, называющееся также реакцией прямого питания (англ. feed forward response), направлено на ограничение риска кавитации и поэтому у древесных растений выражено в большей степени (см. 13.5.1).
Этап 2. Реакция устьиц на идущий от корней сигнал, который дает знать о том, что в непосредственном окружении тонких корней наступает дефицит влаги. Если оставить горшки с почвами экспериментальных растений высыхать медленно, чтобы водный потенциал листьев в сколько-нибудь значимой степени не отдалялся от того, который имеется в почве (горшки заключены в барокамеру под избыточным давлением, рис. 13.13), то, следовательно, устраняется потеря тургора и при высокой влажности воздуха наступает устойчивая редукция ширины устьичных щелей, тесно коррелирующая с уменьшением содержания влаги в почве. Такое явление объясняется усилением продуцирования ABA в тонких корнях как реакцией на возрастающую сухость окружающей их среды. Данная реакция приводит также к ограничению транспирации, контролируемой устьицами, но критическое значение водного потенциала листьев таким образом не достигается. Из-за того, что протяженность «пути» между корнями и кроной дерева часто бывает длинной, эти реакции замедляются. Имеются доказательства того, что решающее значение имеет концентрация ABA в ксилемном соке, а не пассивная аккумуляция ее в апопласгах транспирирующих листьев.
Рис. 13.13. Реакция устьиц на сигнал, идущий от корней: А — корневой сигнал (абсцизиновая кислота) заставляет устьица реагировать на постепенное увеличение сухости почвы, если листья искусственно поддерживаются в тургесцентном состоянии (водный потенциал равен нулю). Это состояние моделируется с помощью повышенного давления (р) в корневой сфере, причем наполненный водой, вколотый в ось побега капилляр показывает уравновешенное нулевое давление. Если передвигать мениск к оси (всасывание), можно вызвать повышение давления в корневой камере; В — на примере пшеницы (Triticum aestivum) показано, что ниже критического значения содержания воды в опытном субстрате линейно снижается листовая диффузионная проводимость (раскрытие устьиц), несмотря на тургесцентное состояние побега
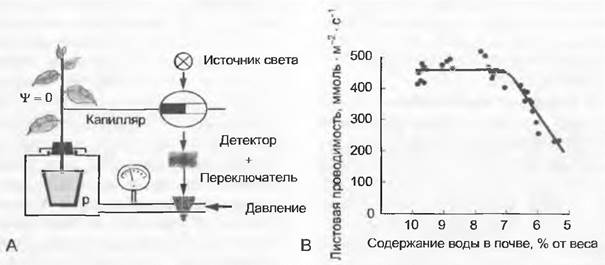
В природе этот механизм доказать очень трудно. Он имеет также «feed forward» характер, с одной стороны, направленный на снабжение питанием и, с другой стороны, предохраняющий, как и реакция на влажность воздуха, от внезапного возникновения настоящей стрессовой ситуации (потери тургора). Известно, что в ксилемном соке деревьев, растущих на сухих почвах, повышен уровень ABA. Тем не менее остается открытым вопрос о том, является ли это пассивным следствием общего замедления транспирационного потока или активным процессом.
Этап 3. Реакции устьиц на потерю тургора. Если водный потенциал листьев, несмотря на предохранительные меры этапов 1 и 2, приближается к осмотическому потенциалу клеточного сока, обратное давление клеточных стенок на протопласт становится нулевым. Мягкие листья увядают еще до достижения этой точки. Вблизи точки потери тургора лежит также критическое значение водного потенциала, при котором происходит замыкание устьиц (пассивно) вследствие обезвоживания (дегидрации). Устойчиво низкий тургор (низкий «рге dawn water potential») повышает чувствительность устьиц к сухому воздуху.
В природе потеря тургора происходит относительно редко. Имеются даже случаи (виды родов Piper, Helianthus), когда устьица, несмотря на заметное увядание, остаются открытыми, это подчеркивает, что устьица в общем начинают реагировать на сигналы водного потенциала очень поздно. Такое своеобразное поведение объясняется тем, что растения, увядая (опускание листовых пластинок), избегают полуденного солнечного облучения, и тем самым уклоняются от перегрева и в конечном счете от потери воды.
Из вышесказанного можно сделать вывод, что возможности растений сохранить благоприятный водный режим с помощью сиюминутных физиологических реакций исчерпываются. Дальнейшее зависит от толерантности к иссушению. Повторяющийся водный дефицит приводит к падению осмотического потенциала (аккумуляция осмотического давления, осмотическая адаптация, осморегуляция), что отодвигает время потери тургора и повышает устойчивость к обезвоживанию.
Этап 4. Распределение биомассы. Растения, которые в течение длительного времени и с повторами испытывают дефицит влаги, реагируют на него усилением роста корней, что идет за счет отложения биомассы в листьях. Благодаря этому баланс повышается в пользу усвоения воды (освоение более глубоких слоев почвы). Если механизмы, применяемые на этапах 1— 4, не достигают цели, наступает этап 5.
Этап 5. Сбрасывание листьев и вследствие этого редукция LAI. Если водный дефицит благодаря этому также не приостанавливается, в конечном счете наступает этап 6.
Этап 6. Изреживание насаждения и замена устоявшегося набора видов новыми засухоустойчивыми видами. Это могут быть виды с особой феноритмикой (беглецы —
от англ. escape), особенно глубокими корнями, развитой суккулентностью (стратегия избегания — от англ. avoidance) или подлинной засухоустойчивостью.
Наиболее эффективные методы «избегания» потери воды — облигатное сбрасывание листвы на время сухого периода и (или) переживание засухи в форме семян, луковиц или подземных побегов. Суккуленты и растения с глубокой корневой системой избегают физиологического стресса, причем последние более успешно переживают длительную засуху. Суккуленты нуждаются в незначительной, но регулярно поступающей влаге, так как они вынуждены довольствоваться только теми запасами, которые содержатся в их тканях. При сухой почве корни суккулентов теряют с ней контакт и изолируются от субстрата. Размеры потери воды тканями сильно варьируют, но большинство тканей может терять до 50 % воды. Подлинная засухоустойчивость, вплоть до полного высыхания, среди высших растений редкость (восстанавливающиеся растения, англ. resurrection plants), но довольно обычна среди лишайников, мхов и водорослей, обитающих в воздушной среде (так называемые пойкилогидрические растения в противоположность гомойогидрическим, стратегии которых к перенесению иссушения описаны ранее).
В природе наибольшее значение имеет реакция на дефицит влаги, соответствующая этапу 6. Хорошо приспособленный набор видов почти не нуждается в реакциях этапов 1 — 5, это значит, что данные виды ничего не вносят в механизм фотосинтеза, который на время должен замедлиться или даже прерваться. Это объясняет, почему вдоль естественного градиента влагообеспеченности на уровне листа обнаруживаются относительно слабые различия параметров водного режима. Различия проявляются умеренно, даже когда на 1 м2почвы в проекции транспирируют 0,01 м2 листьев (пустыня) или 5 м2 (буковый лес). Листья пустынных растений транспирируют (за исключением крайне суровой засухи) не столь уж иначе, чем мезофильные растения, но у них образуются хотя и очень немногочисленные, но полностью работоспособные листья. Эта экономика инвестиций в работоспособные листья тесно связана с их снабжением минеральными веществами и вместе с тем с бюджетом минеральных веществ экосистемы (см. 13.6.3).
Открытия, преимущественно сделанные в течение последних двух десятилетий, показали, что многое в области водного режима на уровне листа, оценивавшееся ранее как негативное явление (в смысле стресс-симптомов), в действительности представляет собой мгновенные противодействующие предупредительные реакции на системном уровне (реакции устьиц на этапах 1 и 2), или следствие высокой активности, а не выражение нанесенного ущерба (низкий водный потенциал); во всяком случае речь не идет о непосредственных воздействиях стресса на большой территории. Существенный вклад в проблему внесли также научные исследования районированных в сухих областях сельскохозяйственных растений. X. Пассиура (Канберра) смог, например, показать, что в очень сухих посевных районах Австралии сорта пшеницы с «плохой» корневой ксилемой дают лучший урожай. Этот парадокс основывается на наблюдении: регулярный дефицит воды в листьях (из-за плохо проводящей ксилемы) в течение вегетативного развития хотя и несколько сдерживает рост, но гарантирует, что ко времени цветения и поспевания зерен вода в почве еще будет. Очень сходно с этим можно оценить вышеописанные «feed forward» реакции, которые равным образом приводят в данный момент, казалось бы, к необоснованным ограничениям, но в течение длительного времени весьма способствуют процветанию и выживанию долго живущих растений.
Путем частичного удаления листьев F. Meinzer (Гавайи) и другие авторы смогли продемонстрировать, что устьица, кроме всего прочего, реагируют компенсаторным образом на потребности всех растений в сообществе и при этом не только приспосабливают к ним саму устьичную диффузию, но и обнаруживают регулировочное поведение, которое включает и аэродинамическую сопротивляемость пограничного слоя. В конечном счете и это сводится к способу измерения транспирации, причем, согласно упомянутым авторам, химические сигналы действовали бы слишком медленно, поэтому предполагаются гидравлические сигналы. Данные примеры также показывают, что употреблявшееся ранее разделение на аутэкологию и синэкологию мешает убедительной интерпретации процессов. Почти все обсуждавшиеся выше реакции объясняются только исходя из их действия на экосистемном уровне.
13.5.3. Поведение устьиц в природе
Наряду с уже описанными воздействиями водного режима ширина устьичных щелей определяется прежде всего световыми ресурсами и тем самым нормой фото
синтеза (и уровнем СО2 в тканях листа). Фотосинтез извлекает из воздуха и доставляет в лист СО2, который диффундирует снаружи через устьица (см. 6.5.7). Пока водный режим это допускает, устьица настроены на диффузионную проводимость, при которой в межклетниках листа содержание СО2 (Сi) поддерживается на уровне около 70 % от концентрации в окружающем воздухе (са). Чем выше степень фотосинтеза (А), тем больше должна быть диффузионная проводимость, чтобы поддерживать устойчивую межклеточную концентрацию (отношение Сi/са). С этим также связаны СO2-газообмен и диффузия водяного пара, что означает тесные связи между максимальной степенью фотосинтеза при нормальном уровне СO2 (Аmах) и максимальной устьичной диффузионной проводимостью (gmах). Чем выше интенсивность фотосинтеза, тем выше и максимальная устьичная проводимость водяного пара (рис. 13.14).
Рис. 13.14. Связь между максимальной интенсивностью фотосинтеза и состоянием транспирации у растений типа С3. При хорошем снабжении водой существует линейная корреляция между максимальной диффузионной проводимостью эпидермиса листа (устьица; gmax) и максимальной интенсивностью фотосинтеза Аmах на единицу листовой поверхности (Аcар) независимо от жизненной формы (точками обозначены хвойные и лиственные деревья, кустарники, разнотравье, злаки, суккуленты и др.). Это означает, что отношение концентрации С02 в межклетниках к его внешней концентрации (С1/Са) у всех растений типа С3 лежит в одной и той же области (0,7 — 0,8) и рассматривается как результат эволюционной оптимизации потери воды и поступления СO2 через систему пор
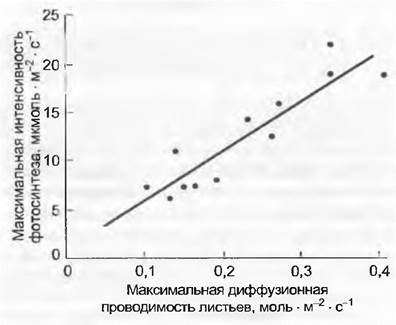
Сопоставление, приведенное на рис. 13.14, включает крайние случаи — быстро растущие травянистые растения и малоактивные, очень медленно растущие виды. Оно затушевывает тот факт, что средние значения для доминирующих видов древесных растений крупных биомов Земли в этих нормативных параметрах в естественных местообитаниях едва различаются, если в сравнение включено достаточно много видов. Глобальное значение gmax по отношению к простой проецированной листовой поверхности составляет для 151 вида древесных растений (218 ± 24) ммоль Н2O м2 • с-1. Травянистые растения и сельскохозяйственные культуры имеют в среднем почти вдвое большие значения, растения САМ-стратегии — значительно более меньшие. Минимальная диффузионная проводимость водяного пара (gmin) листьями в значительной степени определяется наличием отверстий в замкнутых устьицах, поэтому говорить о кутикулярной диффузионной проводимости некорректно. Варьирование gmin по отношению к gmax колеблется от 1/20 (травянистые теневые растения) до 1/300 или еще меньше у суккулентов (у деревьев и кустарников от 1/40 до 1/60, т. е. от 3 до 6 ммоль • м-2 • с-1).
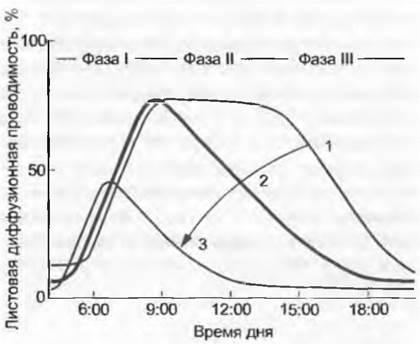
Рис. 13.15. Схематическое изображение дневного хода листовой диффузионной проводимости. При хорошей погоде и не слишком глубоком увлажнении почвы (водный потенциал листьев перед восходом солнца более 0,2 МПа) диффузионная проводимость листьев g (равна устьичной проводимости, так как кутикулярная транспирация очень мала) утром следует за увеличением светового довольствия (фаза I) до максимального значения g. Если достигается критическое значение дефицита давления водяного пара в воздухе (vpdпримерно 10 гПа), значение g снижается параллельно с дальнейшим поднятием vpd (фаза II). После полудня при том же состоянии климатических факторов, как и утром, значение gстановится все ниже и падает почти до нуля (независящие от времени факторы, аккумуляция конечных продуктов в листьях, пассивно увеличивающееся содержание ABA, активное повышение содержания ABA как сигнал от корней об обезвоживании непосредственно примыкающей к ним почвы, кавитация ксилемы и т. д. — фаза III). Так как в течение дня детерминанты g замещают друг друга, проведение статистического анализа для выяснения факторных зависимостей без разделения фаз невозможно. Кривая 1 — высокая влажность воздуха и почвы; кривая 2 — высокая влажность почвы, но пониженная влажность воздуха (vpd>10 гПа); кривая 3 — ситуация с дефицитом почвенной воды. Стрелкой показан увеличивающийся пресс сухости
Устьичная или листовая диффузионная проводимость — это два употребляемых синонимических понятия, так как можно пренебречь разницей между транспирацией воды через устьичные поры и транспирацией через устьица и лежащую между ними кутикулу. Диффузионная проводимость обратна диффузионной сопротивляемости г. Обе величины выводятся, по аналогии с законом Ома, из плоскостной транспирации Тг («транспирационного тока») и градиента водяного пара системы лист — воздух, ∆w («напряжение»; закон диффузии Фикка). При этом принимается, что внутри листа воздух насыщен водяным паром, причем давление пара в листе будет лишь функцией от температуры листа. Употребляются две размерности. Если для g = Тr/∆w используют характеристику (gН2O м-2 • с-1)/(gH2O • м-3), получают g (м • с-1). Если вводят для ∆w (кПа • Н2O, кПа-1 воздуха), то оказывается, что из-за безразмерного описания градиента влажности воздуха g характеризуется теми же размерностями, что и Тг, т. е. в г • м-2 • с-1, или в ммоль • м-2 • с-1. Важно, что в данном случае ∆wвыводится из соотношения: давление водяного пара внутри листа — давление водяного пара вне листа (общее давление воздуха). Отсюда значения g и исчисляемая из фактического значения степень транспирации будут в значительной степени независимы от давления воздуха, которое оказывает влияние на молекулярную диффузию (данные из различных высотных поясов). Это одна из причин, почему данная размерность находит применение все больше и больше.
Результатом взаимодействия условий влажности, солнечного излучения и температуры является характерный дневной график ширины устьичных отверстий (диффузионная проводимость; рис. 13.15). Вутренние часы раскрытие устьиц определяется в первую очередь светом и фотосинтезом. При определенной плотности потока фотонов РFD (у светолюбивых растений при 20 — 25 % полного солнечного излучения) достигается gmах. Если превышается критическое значение дефицита давления водяного пара (см. 13.5.2, по меньшей мере около 8 гПа, или около 65 % относительной влажности при 20 °С), это становится для большинства видов древесных пород определяющим фактором воздействия, и g снижается (при хорошей погоде с позднего дополуденного времени). При одинаковых климатических параметрах значения в послеполуденное время, как правило, ниже, чем в дополуденное. Такие реакции, зависящие от времени, объясняются аккумуляцией продуктов фотосинтеза и повышенным уровнем АБА. Дневной ход g у древесных растений выражается одновершинной кривой с максимумом перед наступлением полудня. Травянистые растения реагируют на погоду большей частью в послеполуденное время, это значит, что может быть выражен второй максимум g, когда напряжение транспирации опять понижается. При пасмурной, влажной погоде определяющим фактором будет один РFD. Диффузионная проводимость определяет также диффузию вредных газов (например, озона), поэтому при сухом воздухе и относительном дефиците воды будут незначительные нарушения.
При дальнейшем раскрытии пор изменения ширины отверстий при данной влажности воздуха почти пропорциональны изменениям транспирации, однако это сравнительно мало влияет на фотосинтез. Поэтому растения могут регулировать транспирацию в широких пределах, не оказывая пропорционального воздействия на фотосинтез. Только при незначительной ширине пор влияние на фотосинтез будет сильным. Эта асимметрия приводит к тому, что устьица листа представляют для транспирационного потока единственный значимый управляемый механизм, в то время как усвоение СO2 дополнительно к диффузии газов испытывает при полном раскрытии устьиц еще одно, примерно в 4 — 5 раз большее сопротивление в клетках мезофилла (у растений С3, у растений С4 сопротивление мезофилла значительно меньше). Устьичное сопротивление представляет собой таким образом (прежде всего у растений С3) только небольшую часть всех препятствий для усвоения СO2.
При широком раскрывании устьиц большое значение приобретает противодействие газовой диффузии в аэродинамическом пограничном слое листа. Величина листа и его опушение повышают сопротивление пограничного слоя, действующее аналогично устьичному сопротивлению, и редуцируют при этом эффективность устьичной регуляции при широком раскрытии отверстий. При незначительном раскрытии пор, а также при стрессовой ситуации, вызванной сухостью, эти аэродинамические факторы для диффузии воды относительно несущественны, но противодействуют тепловой конвекции и отражению тепла от листа, и тем самым снижению давления водяного пара, они могут косвенно содействовать поддержанию водного баланса. Впрочем, большое число растений засушливых областей довольно слабо опушены или вообще лишены опушения, а это противоречит широко распространенному мнению, что опушение представляет собой непосредственную защиту от испарения. Повышенное сопротивление пограничного слоя (которое стоит в одном ряду с устьичным) в любом случае снижает до минимума эффективность устьичной регуляции.
13.5.4. Водный режим экосистем
При высокой влажности почвы лес или луг в летний день при хорошей погоде в Центральной Европе отдают 4 — 5 мм испаряемой в атмосферу воды (1 мм = 1 л на 1 м2). На эту же величину снижаются запасы доступной для растений почвенной воды. Насколько часто это может повторяться (в течение периода хорошей погоды), зависит от содержания воды в корнеобитаемом почвенном профиле (см. табл. 13.3 в разделе 13.7.5.1). Потенциальный запас доступной влаги определяется глубиной профиля, возможными «мертвыми зонами» почвенного скелета (камни) и долей объема, приходящейся на поры средней величины. В качестве грубой прикидки для хорошо развитых, не песчаных почв можно принять общий объем пор 50 % от общего объема (больше в верхних слоях почвы, значительно меньше в глубоких горизонтах). Примерно половина этого объема, около 250 мм воды на I м профиля, свободного от скелетных включений, может считаться вполне доступной. Остальной объем почвенных пор приходится на быстро дренирующиеся крупные пустоты (например, ходы дождевых червей) или очень мелкие тонкие поры, вода которых недоступна растениям. Такой модельный профиль после основательного увлажнения мог бы быть при вышеназванных нормах дневного испарения обеспеченным влагой около полутора месяцев. При менее мощном профиле с большой долей камней или на песчаной почве с меньшей влагоемкостью период, в течение которого экосистема может обходиться без осадков, укорачивается. Ярко выраженная «feed forward» реакция устьиц удлиняет его (см. 13.5.2 — этапы 1 и 2). Атмосферная вода, которая стекает или остается на листовой кроне как смачивающая влага, для экосистемы потеряна.
Эти зависимости описываются уравнением водного режима:
N = Е + Tr + I + А + dB,
где N — сумма осадков, равна сумме эвапорации с поверхности почвы — Е, транспирации растений — Тг, потери перехвата (смачивающая влага) — I, оттока воды (поверхностный сток и просачивающаяся на глубину инфильтрационная влага) — А, изменений в запасах доступной влаги в почве — dB. За долгий промежуток времени (например, за весь год) изменение запасов почвенной влаги равно нулю. Е, Тг и I могут быть объединены в понятие эвапотранспирации, сокращенно ЕТ. Тогда N = ЕТ + А.
В сомкнутом растительном сообществе решающий фактор водного режима — перехват осадков. Очень много воды задерживается листьями, особенно при часто повторяющихся, но необильных дождях (на смачивание всего насаждения идет 1 — 2 мм воды). В бассейне Амазонки почти четверть общей суммы осадков (около 2000 мм в год) падает на потери перехвата, приближающиеся к объему стока самой Амазонки. На объем перехвата оказывают влияние характер расположения листьев и LAI. Густые хвойные леса задерживают вдвое больше воды, чем луг. При корчевке лесов или покосе перехват влаги ликвидируется, сток повышается/
В Центральной Европе ежегодная сумма испаряемой воды (ЕТ) составляет около 500 мм (максимум на небольшой высоте над уровнем моря в южной части — до 650 мм). Примерно 70% всей испаряемой воды, которую теряет Европа, уходит через листовые поры, что подчеркивает первостепенную роль растительного покрова для водного режима. Особенно способствуют испарению растения в сухой период, так как они с помощью своей корневой системы добираются до резервов воды, которые без растительного покрова не достигали бы состояния испарения (см. табл. 13.3 в разделе 13.7.5.1, а также 13.1.2). С другой стороны, растительность способствует образованию почв с высокой способностью к накоплению запасов воды и препятствует поверхностному стоку. Сомкнутые, хорошо обеспеченные водой растительные сообщества испаряют больше воды на единицу поверхности, чем открытые водоемы (например, озера), благодаря своей большой листовой поверхности, часто более высокой температуре и хорошим аэродинамическим связям с атмосферой.
Источники воды, питающие растительность, можно обнаружить с помощью дейтерия, стабильного тяжелого изотопа водорода (в тяжелой воде). Тяжелая вода испаряется несколько медленнее, чем нормальная вода, вследствие чего водяной пар (соответственно облака и дожди) во всем мире всегда содержит несколько меньше D2О, чем грунтовые воды. Растения со сравнительно малым количеством дейтерия в тканях (доказано на масс-спектрометре) получают влагу от поверхностных «свежих» почвенных вод в противоположность растениям, корни которых имеют контакт с грунтовыми водами. Это аналогично поведению тяжелого и относительно стабильного изотопа кислорода 18О по сравнению с «нормальным» 16О. Вода Н218О испаряется медленнее и аккумулируется поэтому в морях, в осадках которых находят 18О в количествах, пропорциональных интенсивности испарения и соответственно температуре, что используется в палеоэкологии для реконструкции климатов. При высокой транспирации тяжелый изотоп кислорода сильнее насыщает и ткани растений, что дает возможность вместе с дискриминацией 13С (см. 13.7.4) реконструировать закономерности изменений водного и углеродного режимов (например, пробы из старых годичных колец деревьев).
Рост растений и потребление воды теснейшим образом связаны друг с другом. Чтобы произвести 1 кг растительной массы, требуется от 500 до 1 000 л воды, для растений С4 — 250 — 400 л. В засушливых областях CAM-растениям необходимо особенно малое количество воды, «плата» за исключительно ночной газообмен — очень медленный прирост. Понятие эффективности использования воды (англ. water use efficiency— WUE) не имеет однозначного определения. Классическое определение пришло из сельского хозяйства. Там WUE означает количество полезного урожая (например, зерна, сена) на единицу использованной воды, попадающей на поверхность почвы (грамм сухого вещества на литр воды). Если, например, у зерновых культур около 50% продуцируемой растительной массы падает на зерно (harvest index), потребление воды на единицу урожая, как и на единицу биомассы, удваивается. Неизбежный расход воды на эвапорацию с поверхности почвы и потери перехвата засчитываются сюда же (это важно для поливного хозяйства). Много позже WUE была принята в качестве термина в физиологии газообмена и теперь определяет квоту фотосинтеза листа и одновременно транспирации (ммоль/моль). Параллельное существование двух определений вызывало немало путаницы и требует в каждом случае четкого определения применяемой концепции. К тому же слово «эффективность» амбивалентно, так как в экологическом смысле опять же требуется определить различия между продуцированием массы и устойчивостью (через «процветание», см. 12.1). Как предлагает В.Лархер, следовало бы применять нейтральное понятие «коэффициент».