БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
13. РАСТЕНИЯ В ЖИЗНЕННОМ ПРОСТРАНСТВЕ
13.6. Бюджет питательных веществ
Питательные вещества растений, за исключением содержащихся в воде и СО2, делятся на две группы: так называемые минеральные вещества, или элементы, непосредственно происходящие из исходного неорганического материала почвы (или из атмосферной пыли), и азот, происходящий из атмосферы, но в ходе круговорота питательных веществ в конце концов оказывающийся в том же почвенном растворе, что и собственно минеральные вещества. Часто в понятие минеральных веществ включают и азот в его растворимой неорганической (минеральной) форме (NО-x и NН+4). При недостатке каждый питательный элемент может быть фактором, лимитирующим рост растений. В основном это все же азот и фосфор, доступность которых в природе критическая. Поэтому данный раздел прежде всего посвящен роли этих двух элементов в экосистеме, особенно азота. Значение остальных питательных элементов растений и лежащих в основе круговорота азота химических процессов было рассмотрено в разделах 6.2, 6.6 и 9.2.
13.6.1. Доступность питательных веществ почвы
Вопрос о том, насколько много питательных веществ фактически имеется в распоряжении одного растения или целого сообщества, относится не к содержанию их в почве, а к их доступности. Часто только очень малая часть этих веществ присутствует в доступной для растений форме. На искусственных субстратах (или в гидрокультуре) доступность часто соответствует концентрации веществ в растворе субстрата, но не так обстоит дело в системах с естественным круговоротом минеральных веществ и естественным ростом. В этих условиях водная вытяжка из почвы свидетельствует очень мало или вообще не свидетельствует о фактической доступности веществ. Шведский ученый Т. Ингестад доказал опытами с питательными растворами, что корни в состоянии извлекать питательные соли из растворов таких малых концентраций, которые лежат близ границы доказательств традиционными методами. Тем не менее, непрерывно варьируя в питательном растворе потребляемые растением количества питательных солей, можно добиться почти экспоненциального роста растения, несмотря на такие пониженные концентрации.
Тем самым было приведено чрезвычайно важное с точки зрения экологии доказательство, что степень роста определяется нормой поступающих питательных веществ (англ. nutrient addition rate), а не концентрацией их в субстрате свыше относительно низких нижних граничных величин. Ингестад продемонстрировал, что с помощью нормы поступления питания на хемостате можно «устанавливать» степень роста, и, если давать питательных веществ больше, чем растение в состоянии усвоить, концентрация в растворе повышается. Эти связи не для всех минеральных веществ значимы в одинаковой степени, но сам принцип представляет собой исходный момент для понимания круговорота питательных веществ в природе и в то же время источник недоразумений, возникающих при сопоставлении агроценозов и естественных систем. В первом случае обеспечение потребностей (из практических соображений) делится на отдельные фазы, поэтому избытки удобрений поступают в почвенный раствор и грунтовые воды. Во втором случае предложение и усвоение питательных веществ (по существу биологическое производство посредством деятельности микроорганизмов) тесно сопряжены, поэтому обильный рост может происходить даже тогда, когда почвенный раствор очень беден питательными веществами, и аграрный метод анализа почв обнаружил бы дефицит последних. В опытах для каждой почвы при постоянной влажности и температуре можно учесть регулируемое микробами высвобождение неорганических азотных соединений. При этом в качестве продукта минерализации в слабокислых или нейтральных, хорошо аэрируемых почвах образуются в основном ионы NO-3, в кислых грубогумусовых или модеровых — особенно ионы NH-4.
Наилучший пример тесной связи между высвобождением питательных веществ и их потреблением — первичные влажно-тропические леса в Амазонской области на древних сильно выщелоченных почвах, с которых стекающая вода после сильных ливней по качеству близка к дистиллированной. Система тесно «связанная». Круговорот питательных веществ таким образом полностью замкнут, что может быть компенсировано возможными «брешами» в системе, когда, например, минеральные вещества почвы обогащаются за счет частиц пыли, принесенных из Сахары, что лишь недавно было доказано. Тесная связь между высвобождением и потреблением минеральных веществ обеспечивается свободно живущими и симбиотическими микроорганизмами (микориза). Они представляют собой в известной степени «склеивающие вещества» системы. В явно сезонном климате эта связь время от времени разрывается, поскольку предложение и спрос не синхронизированы. В этих случаях большую роль играют нейтральные «пулы» питательных веществ в почве (обмен ионами, образование комплексов, микробная биомасса).
Снабжение экосистемы всеми элементами, кроме азота, производится в конце концов за счет сохранившихся резервов в пронизанной корнями подпочве и биомассе, если не считать внесения летучей пыли. Если их большая часть находится в биомассе, как в тропических лесах, существует очень большой риск, что после пожара может быть потерян весь капитал минеральных веществ системы, который аккумулировался тысячелетиями. Снабжение азотом, напротив, может осуществляться путем микробной фиксации азота воздуха, запасы которого теоретически неисчерпаемы (см. 9.2.1). Но азотфиксаторы требуют больших количеств фосфатов, поэтому фосфорный и азотный режимы связаны между собой уже в этой сфере.
В водоемахколичество, состав и сезонная ритмика развития бентосных и планктонных растительных организмов решающим образом зависят от содержания в воде питательных веществ, особенно азота и фосфора.
В качестве примеров богатых питательными веществами эвтрофных водоемов с высокой продуктивностью можно назвать: в области морей — так называемые «зеленые океаны» (особенно у западных берегов материков, например Перу, Западной Африки, где ветер оттесняет бедные питательными веществами поверхностные слои воды и устремляет на их место богатые глубинные слои, или в антарктических и арктических морях с обусловленными температурами сезонными движениями воды), а также коралловые рифы, прибрежные мангры, ватты и устья рек (аллювиальные поймы) с хорошим снабжением питательными веществами, приносимыми с суши; в области пресных водоемов — озера на низменности с изменяющейся температурой воды, с перемешиванием водных слоев весной и осенью или реки, богатые взвешенными частицами. Им можно противопоставить бедные питательными веществами мезо- и олиготрофные водоемы со средней или малой продуктивностью, например, «синие океаны», без просачивающихся кверху глубинных слоев воды (например, части Средиземного моря или центральная часть Южной Атлантики), холодные горные озера, дистрофные болотные воды (с высоким содержанием гумусовых веществ и pH от 3,5 до 5,0), холодные горные ручьи.
В водоемах с изменяющейся температурой воды (например, в морях и озерах умеренной зоны) фитопланктон достигает пиковых значений после весенней циркуляции воды вследствие хорошего снабжения питательными веществами (а также благоприятного светового и температурного режима), затем летом эти значения снижаются из-за расхода питательных веществ и еще раз несколько поднимаются во время осеннего движения воды перед установлением зимнего низкого уровня. Как результат усвоения СO2 часть автотрофных водных растений (макрофиты в щелочных пресных водоемах) образует отложения извести (например, известковый туф, озерный мел), причем вместо хорошо растворимого бикарбоната кальция образуется почти нерастворимый карбонат кальция:
Са(НСO3)2 —> СаСO3 + Н2O + СO2.
В водоемах с обилием организмов и недостаточным перемешиванием воды часто образуются как следствие деятельности гетеротрофных деструентов бедные кислородом или вообще лишенные его глубинные слои или отложения сапропеля, в которых могут существовать только немногие специализированные анаэробные виды (особенно бактерии).
13.6.2. Источники и накопители азота
Бюджет азота занимает особое место в бюджете минеральных веществ. Растения содержат азота примерно в 10 раз больше, чем фосфора. Запасы азота в тканях пропорциональны содержанию протеина. Количество последнего приблизительно рассчитывают, умножая процентное содержание азота на 6,25. Концентрация протеина в высушенных при высокой температуре тканях колеблется между 1 % в древесинных пробах и 25 % в листьях быстрорастущих травянистых растений. В листьях летнезеленых деревьев содержание его колеблется в пределах 13 — 15 % (2 — 2,5% азота), в вечнозеленых листьях (хвоя) — примерно вдвое меньше (табл. 13.1). В эндосперме зерен хлебных злаков концентрация протеина составляет примерно 13 % (т. е. около 2 % азота).
Из-за того, что азот имеет основное значение в обмене веществ, анализы на азот в экологических исследованиях проводятся наиболее часто. До конца 80-х годов XX в. преобладал так называемый метод Кьельдаля, который основывался на растворении в серной кислоте при высокой температуре (320 °С) и последующей нейтрализации NaOH с одновременной отгонкой высвободившегося аммиака и последующим титрованием в слабощелочной среде (общий азот = азот, по Кьельдалю). В настоящее время используют преимущественно физические методы (элемент-анализаторы). Часто применяется метод растворения через сжигание при более 500 °С в чистом кислороде, после чего проводится газовый анализ с использованием гелия в качестве газа-носителя (например, СМН- анализаторы, которые дают количество элементов, измеряемое в процентах от высушенной при высокой температуре навески). Норма расходов при этих физических методах соответствует нескольким миллиграммам растительного порошка на один анализ.
Таблица 13.1. Концентрация азота (N) в листьях и специфическая для важнейших биомов поверхность листьев (SLA)
Растения / тип растительности |
N, % |
SLA, м2/кг |
Травянистые растения: |
||
двудольные полевые культуры |
3,8 |
24 |
хлебные злаки |
3,4 |
25 |
умеренные злаковые сообщества |
2,6 |
17 |
тропические злаковые сообщества Древесные растения: |
1,1 |
|
тропический лес сезонного климата |
2,7 |
14 |
лиственный лес умеренной зоны |
2,0 |
12 |
тропический вечнозеленый лес |
1,7 |
10 |
вечнозеленый лес умеренной зоны |
1,3 |
6 |
субтропический лавролистный лес |
1.1 |
4 |
склерофильный буш |
1.1 |
7 |
вечнозеленый хвойный лес |
1,1 |
4 |
Примечание. Средние значения N для 5 — 40 (большей частью около 10) характерных видов (стандартные ошибки около +8 % для N и ±15% для SLA) даны в % от сухого веса, SLA— в м2 листовой поверхности на 1 кг сухого веса листьев.
За исключением самых молодых примитивных почв, большая часть азота, необходимого для ежегодного роста растений, происходит от рециклизации, т. е. от разложения микроорганизмами мертвой растительной субстанции. При этом восстановленные формы азота в хорошо аэрированных умеренно кислых и нейтральных почвах постепенно окисляются (см. Nitrosomоnas, Nitrobacter) и опять перерабатываются в нитраты или в очень небольших количествах в качестве веселящего газа N2O, или N2 выбывают из системы (рис. 13.16).
Рис. 13.16. Азот в экосистеме: А — система без антропогенного внесения азота с естественным связыванием N2 путем свободной и симбиотической азотфиксации, а также с получением растворимых в дождевой воде окисидов NO, от молний, естественных пожаров и вулканических явлений. Поступление и расход азота почти уравновешены; В — экосистема с сильно повышенным поступлением азота из антропогенных источников. Здесь принято, что система уже в значительной степени насыщена азотом и только малая часть дополнительного N интегрируется в круговороте азота (прирост биомассы, повышенный N — пул в гумусе). Большая часть азота покидает систему (грунтовые воды, повышенная азотная газовая эмиссия, создающая парниковый эффект). Внесение азота на рис. В (дано приблизительно, по современной оценке) для индустриальных районов. Цифры приведены в килограммах N на 1 га в год
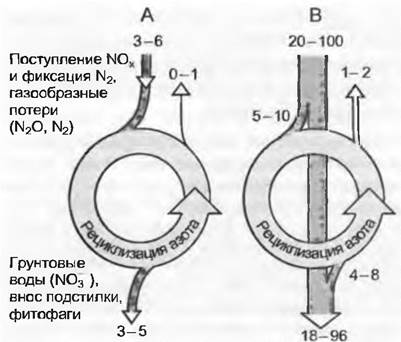
В очень кислых и сырых почвах растворимый азот находится преимущественно в аммонийной форме. Новые результаты показывают, что свободные органические соединения азота даже в таких условиях доступны для растений (например, в тундре). Атмосферные источники растворимых соединений азота — это процессы окисления в атмосфере (молния, огонь) и в последнее время антропогенные NОx-соединения из процессов сгорания. Вторым из важных атмосферных источников можно считать и аммиак из фекалий. Третий источник представляют свободно живущие цианобактерии и четвертый — симбиотические системы (см. 9.2, клубеньковые бактерии на корнях бобовых, симбиоз со специализированными грибами). В зрелых системах (поздние стадии сукцессий) фиксация азота из воздуха для покрытия годичной потребности играет очень небольшую роль, даже если имеются в наличии бобовые, но она важна для создания долговременного фонда азота в почве. По сравнению с рециклизацией в слабоантропогенных экосистемах все остальные источники азота имеют довольно невысокое значение и должны только компенсировать возможные «бреши» в системе (вышеупомянутое высвобождение газообразного азота, потери с инфильтрационной водой и потреблением фитофагами, фиксация в почвенном гумусе). В умеренных лесах годичная потребность в поступлении нового азота составляет в условиях скомпенсированного за многие годы азотного баланса примерно 5 — 6 кг • га-1. В Центральной Европе антропогенно обусловленное поступление азота из воздуха составляет 20 — 30 кг • га-1 (в экстремальных случаях до 100), что является одной из причин, почему в настоящее время в грунтовых водах обнаруживается так много нитратов.
Существенный источник минеральных веществ (и азота) — горизонтальный трансфер питательных веществ в ландшафте. Он может быть направленным и диффузным. К направленному переносу относятся, например, снос растительной ветоши с вершин в западины регулярно дующими в определенных направлениях ветрами или систематическое перемещение питательных веществ животными и человеком (например, зверями с прогалин в лес или в историческое время перенос подстилки из леса в конюшни и на поля). Диффузное перемещение следует из большей доли вероятности того, что богатые питательными веществами частицы из хорошо снабжаемой системы перемешаются скорее в плохо снабжаемую («бедную»), чем наоборот. Через много лет такие потоки вещества из «питательных» территорий в «истощенные» суммируются и образуют мозаику доступности питательных веществ в ландшафте.
Многие многолетние растения растут неравномерно и получают большую часть питательных веществ и азота из собственных запасов, как, например, при весеннем распускании побегов в районах с холодной зимой. В этих случаях процессы инвестиции и потребления питательных веществ далеко отстоят друг от друга по времени.
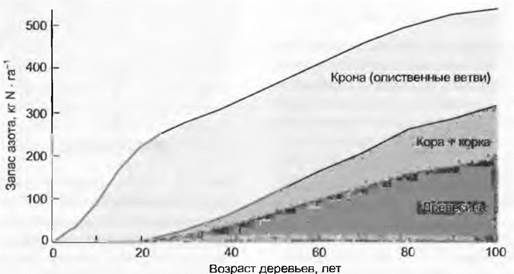
Важнейшие запасы азота в экосистеме содержатся в биомассе и почвенном гумусе. Значение гумуса как накопителя азота увеличивается по направлению к полюсам и достигает минимума во влажных тропических лесах из-за крайне малых запасов гумуса на 1 м2. Концентрация азота в гумусе примерно в три раза больше, чем в листьях, и в 10 — 20 раз больше, чем в древесине. Этот удивительный факт говорит о том, что значительная часть почвенного азота встроена в ароматические углеродные соединения и почвенные пептиды, в которых отношение С/N составляет около 15. Химически связанный таким образом азот недоступен для растений и едва доступен для микроорганизмов. Если азот попадает в гумусовые фракции, он на долгое время исключается из круговорота. Гумификация, т. е. закрепление углерода в почве в форме комплекса гуминовых кислот и пептидов, лежит в основе конкуренции за азот между растениями. Эти резервы могут быть активированы в результате мощного механического нарушения почвы или известкования. При корчевке лесов активизируются и процессы разложения, которые увеличивают поступление азота в водосборы (рис. 13.17), а также удовлетворяют потребность восстанавливающегося леса в азоте (рис. 13.18).
Рис. 13.17. Высвобождение азота в поверхностные воды после сплошной рубки в Нью-Брунсвике (США). Пиковые значения концентрации нитратного азота на поверхности осушенного ручья соответствуют ежемесячным потерям азота примерно 5 кг • га-1. Сумма за первый год после вырубки составила около 70 кг • га-1
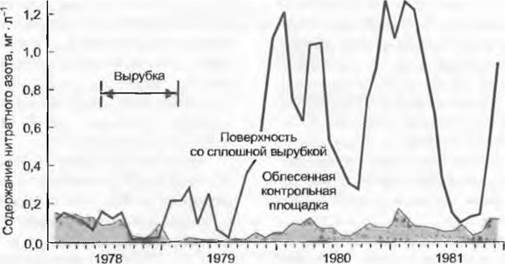
Рис. 13.18. Восстановление запасов азота в биомассе елового леса в Австрии после сплошной рубки. С момента завершения формирования крон (около 25 лет) запас азота в них остается постоянным, запас в стволах продолжает увеличиваться. График наглядно показывает, что в 100-летнем лесу почти половина общего азота сосредоточена в олиственных ветвях
13.6.3. Стратегии инвестиций азота
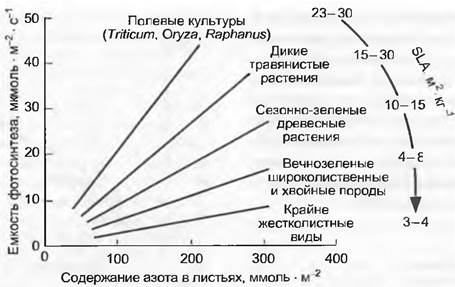
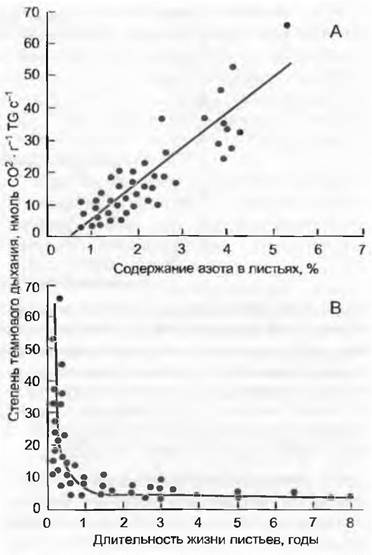
Экономика инвестиций азота — одно из краеугольных положений функциональной экологии растений. Успех вида в изменяющихся внешних условиях определяется тем, сколько азота, где и на какое время было введено в действие. Ткани с высоким содержанием азота (протеина) отличаются высокой метаболической активностью (фотосинтез, дыхание, образование новых тканей; рис. 13.19), но одновременно по тем же самым причинам они очень привлекательны для фитофагов. Преждевременные потери таких растительных тканей из-за нарушений, довольно тяжелы. Связи между максимальной степенью фотосинтеза Аmaх и содержанием азота на единицу листовой поверхности в пределах, определенных морфотипов листа настолько тесные (и линейные), что по данным содержания азота с относительно малой вероятностью ошибки можно предсказать Amax (рис. 13.20). Благодаря тесному и также линейному соотношению между Аmах и gmax (максимальная устьичная диффузионная проводимость для водяного пара; см. рис 13.14) выявляется двойная зависимость.
Рис. 13.19. Распределение азота в листьях кроны Eucalyptus grandis. Зонирование имеет следующие основания: вверху (соответственно, снаружи) доступно больше света, «солнечные» листья имеют большую емкость фотосинтеза и поэтому больше протеина. Листья внутренней части кроны получают меньше света, они старше, поэтому часто и более склероморфные (азот «разбавляется» за счет большего количества углерода), у них меньше площадь (SLA) и меньше потребность в RubisCo (ключевой фермент фотосинтеза)
Рис. 13.20. Зависимости между емкостью фотосинтеза, содержанием азота в листьях и характерной листовой поверхностью. Между содержанием азота на единицу листовой поверхности и наиболее высокой относительно листовой поверхности степенью фотосинтеза при нормальном содержании СO2 в воздухе существует тесная линейная связь. Угол наклона прямой, выражающей зависимость, снижается с уменьшением листовой поверхности на грамм сухого веса (SLA, м-2 • кг-1). Листья с маленьким SLA, как правило, толстые и (или) более жесткие и долголетние. В них связано относительно большее количество углерода в нефотосинтетических структурах
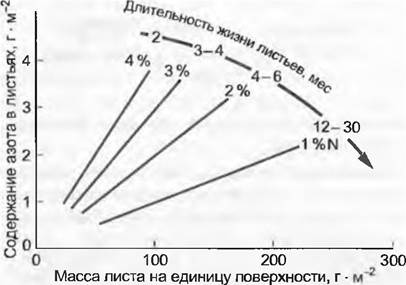
Значения обеспечения азотом на единицу листовой поверхности также тесно коррелируют с SLA или обратной величиной LMA (англ. leaf mass per area — масса листьев на единицу площади, г • м-2) (рис. 13.21). Листья, 1 г сухого вещества которых приходится на небольшую поверхность, содержат меньше N и больше С, поэтому для травоядных они менее привлекательны, но они также меньше ассимилируют и относительно инвестиций углерода на единицу поверхности довольно «дорогие».
Рис. 13.21. Зависимости между инвестициями азота и углерода в листья. В пределах, определенных морфотипов листа существует линейная связь между количеством азота на единицу листовой поверхности и сухим веществом на единицу листовой поверхности (leaf mass per area, LMA= 1 /SLA), независимо от вида растения. Такое группирование вдоль дискретных регрессий соответствует примерно одинаковой длительности жизни этих типов листьев (сходные концентрации азота в % от сухого веса и длительность амортизации, т. е. времени, за которое лист оправдывает «издержки своей конструкции»). Снижение процентного значения азота с увеличением длительности жизни листа может иметь две причины: редукция содержания N в протопласте или увеличение массы клеточных стенок при таком же снабжении протопласта азотом. Как правило, обе эти причины имеют место
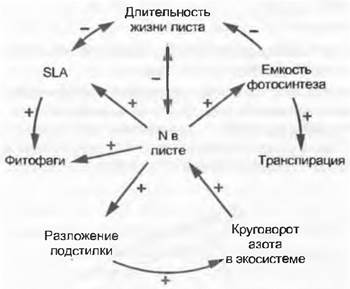
Очевидно, что такие различия в инвестициях могут быть компенсированы только длительностью функционирования. Листья с малым процентом азота и большим углерода должны долго оставаться активними, чтобы при низкой степени фотосинтеза возместить собственные затраты углерода и иметь возможность доставлять ассимиляты по всему растению. Такие листья обычно существуют долго (склерофильные листья, вечнозеленые иглы хвойных). Напротив, листья с большим SLA и высокой концентрацией N в % (см. табл. 13.1 в разделе 13.6.2) живут недолго, уже за несколько дней «амортизируются» (травянистые растения). Итак, по-разному снабжаемые листья после естественного старения разлагаются как подстилка с очень различной скоростью. Тип листа определяет таким образом и скорость рециклизации азота в системе. Отсюда вытекает многомерная взаимозависимость фотосинтеза, водного режима, длительности функционирования, риска поедания животными, разложения подстилки и круговорота азота в экосистеме (рис. 13.22).
Рис. 13.22. Центральная роль азота в особенностях листьев. Концентрация азота в листьях сама по себе препятствует разнообразным изменениям листьев или обусловливает их как в самом листе и отдельном растении, так и в экосистеме. Здесь даны некоторые особенно важные связи. Стрелки показывают направление воздействия, знаки «+» или «-» обозначают направление реакции в случае, если концентрация азота повышается (!) и интенсивность роста растения возрастает. Знаки «+» или «-» меняются на обратные, если концентрация N и связанная с ним интенсивность роста падают, причем отношение к последней проявляется только в пределах, определенных морфотипов, что подтверждается на примере одинаково быстрорастущих летне-зеленых и вечнозеленых древесных растений. Часто принимаемое влияние степени транспирации на питание листьев не доказано и невозможно, поэтому в данном случае стрелка отсутствует. Высокая влажность почвы может способствовать как доступности азота, так и транспирации, но это — совпадение (т. е. первое никак не причина второго). Растения при постоянно насыщенном водяными парами воздухе прекрасно процветают без транспирации (движение воды и транспорт питательных веществ в ксилеме, тесно связанные с флоэмными функциями)
Некоторые из этих зависимостей настолько прочны, что они значимы во всех биомах и для всех жизненных форм, как показал П. Райх на многих примерах (рис. 13.23). Как гласит основное правило, с возрастанием зрелости сообщества в сукцессионном ряду чаще попадаются многолетние, богатые углеродом и более слабые в отношении ассимиляции типы листьев, если не существует сезонных ограничений (листопадные виды в области с холодной зимой). Чем дольше питательные вещества связываются листьями, тем меньше риск потери их системой. Каждый листопад означает риск выноса из системы этих питательных веществ. Ограничения также устанавливаются и самозатенением растений. Длительное функционирование листьев часто в целом связано с медленным ростом или низким LAT.
Рис. 13.23. Митохондриальное дыхание и концентрация азота в листьях: А — с увеличением концентрации азота в листьях увеличивается их темновое дыхание (ср. рис. 13.20); В — с увеличением темнового дыхания падает продолжительность жизни листьев. Эти сравнения охватывают листья растений всех климатических зон. Каждая точка обозначает отдельный вид
Молодые (рудеральные) сообщества растений состоят из быстрорастущих малолетних растений с богатыми азотом, тонкими, быстро амортизирующимися листьями, которые после отмирания очень быстро разлагаются. Некоторые растения с долго живущими листьями (например, хвойные деревья) сохраняют свои старые листья еще какое-то время в кроне, когда они из-за затенения более молодыми по сути дела больше не поставляют сколько-нибудь значимой нетто-продукции углерода. На этой стадии такие листья представляют собой живые консервы питательных веществ, которые при необходимости могут быть извлечены (например, при весеннем распускании, потерях листьев из-за фитофагов, дефиците питательных веществ из-за высыхания верхнего слоя почвы). У ели (Picea abies) хвоинки старше 4 — 5 лет относятся к этой категории.
До некоторой степени описанные явления дополняются другими защитными механизмами. Растения несомненно могут сдерживать определенных фитофагов путем синтеза алкалоидов, гликозидов, фенолов и терпенов, образования масел, млечного сока или смолы. Правда, эти защитные механизмы требуют затрат, и фитофаги часто к ним устойчивы. Смертельно ядовитый для человека и крупного рогатого скота тисс (Taxus baccata) для некоторых диких животных — излюбленное лакомство. Эвкалипты, несмотря на очень высокое содержание эфирных масел, ежегодно теряют до половины своих молодых побегов из-за фитофагов. Насколько известно, ни одна трава не является слишком ядовитой для улиток. Бедность протеинами и жестколистность представляются наиболее действенным средством для увеличения длительности жизни листьев. Чистая продуктивность недолговечных листьев с высокой активностью и долгоживущих с низкой может быть почти одинаковой, что объясняет, почему растения с комплементарными стратегиями листьев могут весьма успешно сосуществовать без принципиальных различий в степени роста и процветания. Примеры — кедровая сосна и лиственница (Pinus cembra и Larix decidua) в Альпах, черника и брусника (Vaccinium myrtillus и Vaccinium vitis-idaea) в березовой лесотундре Северной Европы или сезонно-зеленые и вечнозеленые виды в средиземноморском мелколесье.
Близко к понятию эффективности использования воды (water use efficiency, см. 13.5.4) широко употребляемое и столь же двусмысленные понятие эффективности использования азота (nitrogen use efficiency — NUE). NUE трактуется очень неоднозначно, часто для представления о том, насколько растение «эффективно», в качестве синонима NLJEиспользуется отношение N/C или просто % N, если растение обходится малым количеством азота. При этом NUE может приводиться только для листьев или для содержания азота во всем растении. NUE определяется фотосинтезом относительно Аmах, чаще всего без учета длительности жизни листьев и тем самым без учета «фотосинтетического урожая» за весь период их функционирования. Так как остается неясным, что же подразумевается в действительности под эффективностью (эффективны для чего?), этого понятия следует избегать и в крайнем случае заменять его «коэффициентом использования азота». Как правило, достаточно содержательно просто понятие концентрации азота (без какой-либо надуманной целесообразности).
Как было объяснено в разделе 12.1, в естественных растительных сообществах, приспособленных к определенным местообитаниям и развивавшихся в них длительное время, на уровне сообщества отсутствует дефицит минеральных веществ, даже если интенсивность роста каждого индивидуума сама по себе почти всегда лимитируется питательными веществами, что имеет значение и для продукции биомассы на единицу площади. Упомянутая в разделе 6.2.2.2 специфичная для каждого элемента симптоматика, представляющая собой кратковременную реакцию на дефицит элемента питания, весьма существенна для сельскохозяйственных растений. При конкуренции видов и генотипов в местообитании остаются, однако, на долгое время, как правило, лишь те таксоны, которые могут справиться с ситуацией дефицита так, что подобных симптомов у них не проявляется. Удивительно, что с помощью анализа на элементы у дикорастущих растений можно лишь в редких случаях выяснить, есть ли дефицит минеральных веществ и, если есть, какого вещества именно. Эти растения растут таким образом, что дело не доходит до дефицита существенных питательных веществ, поскольку при известных условиях их органы, образующиеся в меньшем количестве и меньших размеров, полностью функциональны. В таких случаях рост соответствует наличию ресурсов. Рост, превышающий ресурсные возможности, быстро бы элиминировал такой вид или генотип благодаря снижению жизненности. Растения очень холодных биомов (высокогорья, полярные области), у которых поступление азота затруднено, имеют даже более высокие концентрации азота в листьях, чем сравнимые таксоны из теплых стран, что Т. Чепин назвал «растительным потреблением» (от англ. luxuriousconsumption). Условия этих жизненных пространств не дают возможности выживать с плохо снабжаемыми, малопродуктивными листьями. У таких приспособившихся видов оптимальное (экономное) потребление ограниченных ресурсов обеспечивается контролируемым ростом.
13.6.4. Гетерогенность почв, конкуренция и симбиоз в корневой сфере
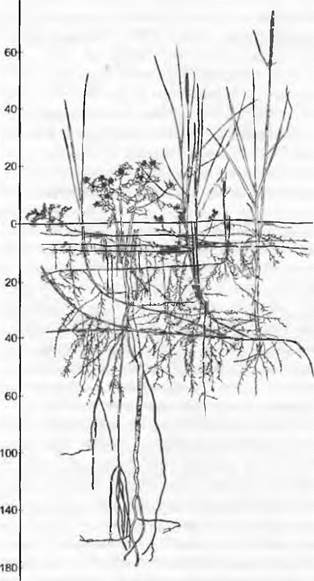
Питательные вещества распределены в почве неравномерно, и различные виды растений охватывают своими корнями разные почвенные горизонты (рис. 13.24). У этой гетерогенности тем большее значение, чем менее подвижны питательные элементы, особенно фосфаты. Гетерогенность имеет при этом 4 компонента: 1) фактически неравномерное распределение веществ в почвенном пространстве; 2) неравномерное (помехи в почве) и очень видоспецифичное распределение корней в почве; 3) различные у разных видов выгодные процессы (например, типы образования микориз и «открытые» симбиозы); 4) неравномерное распределение влаги в почве.
Рис. 13.24. Видоспецифичное освоение корнями почвенного профиля на примере пионерной растительности одной дюны Балтийского моря (с Ammophila arenaria, Eryngium maritimum, Elymus furcatus, Leymus arenarius и др.)
Питательные вещества почвы доступны для растений только тогда, когда почва достаточно увлажнена. В сухих почвах не только блокируются микробиологические процессы минерализации, но и парализованы транспорт (диффузия) и усвоение веществ. Многие нарушения развития и даже нападения вредителей (например, вредителей лесов), которые интерпретируются как следствие сухости почвы, в действительности происходят от недостатка питания. У многих из так называемых мелкоукореняющихся растений незначительная часть корней все-таки располагается на большой глубине (см. табл. 13.3 в разделе 13.7.5.1), этим они могут при большой сухости покрывать потребности кутикулярной транспирации (при закрытых устьицах), избегая настоящего иссушения. Недостаток воды относится к одной из проблем снабжения питанием, поскольку приводит к блокированию питательных веществ в наиболее биологически активном верхнем слое почвы.
Это может с новой точки зрения объяснить открытый М. Колдуэллом в штате Юта «гидравлический лифт»: производя корнями ночной перенос воды из бедных питательными веществами, но влажных глубоких слоев почвы в богатые питательными веществами, но бедные влагой верхние слои, растение осуществляет доступ к питательным веществам. Так как влага от верхних тонких корней отдается в освоенную ими ризосферу, совсем незначительные перемещения воды дают большой эффект.
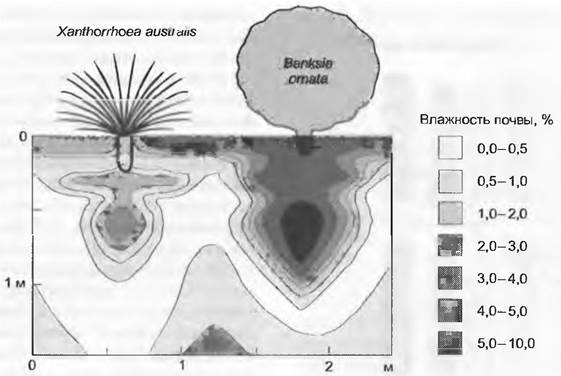
Путем стока выпадающих осадков по наземным структурам и пространственнодифференцированного использования влаги, содержащейся в почве, растения оказывают влияние даже на гетерогенность распределения питательных веществ, особенно во время сухих периодов (рис. 13.25). Эта пространственная дифференциация создает матрицу для сосуществования растений с различным распределением корней по профилю. X. Муни с сотрудниками доказали, что видовое разнообразие в калифорнийском чапарале теснейшим образом связано с такими пространственными моделями освоения почвы.
Рис. 13.25. Воздействие растений на распределение воды в почве и связанной с этим доступности питательных веществ. Благодаря стоку дождевых вод вдоль побегов и специфическому для каждого вида распределению сосущих корней в почвенном профиле (см. рис. 13.24), а также «гидравлическому лифту» возникает, в дополнение к гетерогенному распределению питательных веществ в почвенной структуре, некая модель их доступности. Рисунок иллюстрирует влажность почвы на сухую навеску и вместе с тем доступность питательных веществ в сухом австралийском буше после выпадения 24 мм осадков на долговременно сухую почву (Найнти Майлс Плейн, Южная Австралия)
Использование различных источников азота в почве проявляется в специфичном для каждого вида изотопном составе азота в растениях. В процессах разложения и перестройки в почве стабильный 15М-изотоп (по сравнению с соединениями 14N) метаболизируется несколько медленнее, поэтому, с одной стороны, 15N запасется в почве, а с другой — доступный для растений азот большей частью несколько беднее изотопом 15N, чем азот воздуха, причем это сильно зависит от того, из каких фракций (на какой глубине почвы) этот азот выработан микробами. Наряду с этими двумя различающимися по изотопам компартиментами азота, бобовыми с помощью симбионтов создается третий, в котором почти без изменения сохраняется такое же соотношение 15N/14N, как в атмосфере (15N не различается клубеньковыми бактериями). Исследования в тундре и высокогорьях, проведенные недавно, показали, что вересковые, осоки, бобовые и все остальные виды растений как четвертая группа используют совершенно разные азотные фонды почвы. Вересковые используют азотные фонды крайне бедные 15N, Сурегасеае имеют доступ к фонду, особенно богатому 15N. Маркированное 15N удобрение позволяет проследить пути азота в экосистеме (другие стабильные изотопы см. 13.5 4, 13.7.4).
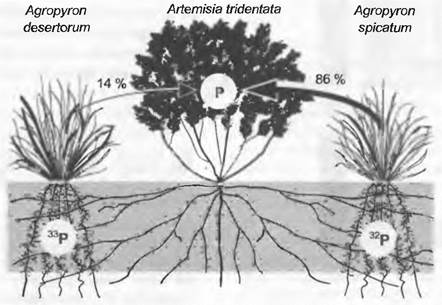
Гетерогенность, связанную с конкуренцией, можно продемонстрировать на примере потребления фосфора. К сожалению, фосфор не имеет стабильных изотопов, однако можно использовать радиоактивный изотоп, с помощью которого при высокой степени разведения можно проследить пути фосфора от определенных источников Р в почве к растениям. Водном классическом эксперименте (рис. 13.26) М. Колдуэлл с сотрудниками выясняли, откуда доминантный в Большом Бассейне (США) кустарничек Artemisia tridentata получает фосфор, когда он делит почвенное пространство с двумя видами злаков рода Agropyron. Agropyron desertorum — агрессивный неофит, угрожающий вытеснить A. tridentata, a A. spicatum — аборигенный вид, издавна свойственный флоре Большого Бассейна. Маркированные 32Р и 33Р фосфорные удобрения были введены в корневое пространство обоих видов Agropyron, и спустя некоторое время определено соотношение обоих изотопов у Artemisia. Результат: Anemisia имеет мало шансов на то, чтобы приблизиться к фосфору в сфере влияния A. desertorum, фосфор в тканях Л. tridentata происходит почти целиком из корневого пространства A. spicatum. Неофит в этом жизненном пространстве — «расхититель» фосфора.
Рис. 13.26. Корневая конкуренция за питательные вещества почвы. Типичный кустарничек Большого Бассейна Artemisia tridentata (вид полыни) произрастает в состоянии конкуренции с Адгоругоп desertorum (занесен из Евразии) и A. spicatum (аборигенный вид). Источники фосфора для полыни познаются из количественного соотношения абсорбированных радиоактивных изотопов 32Р/33Р. Два различных изотопа фосфора случайным образом инъецировались в сферы корневых систем обоих злаков (график показывает один такой случай). A. tridentata доступен в основном только фосфор от аборигенного злака; фосфор из корневой сферы злака-неофита доступен полыни только в малой степени, хотя все растения микоризообразующие
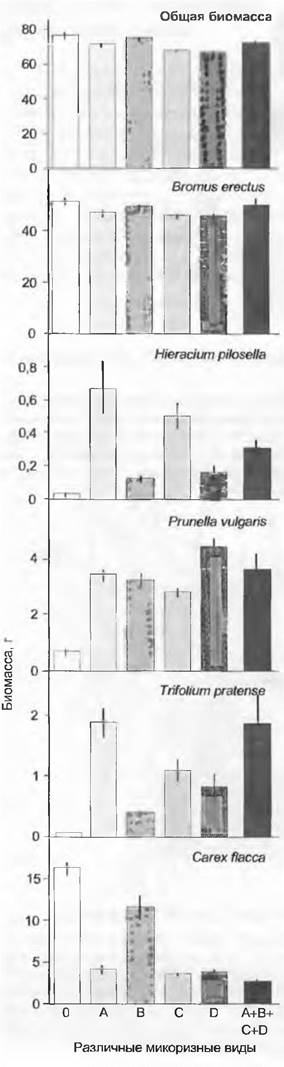
Аномальное неравенство возможностей отдельных видов для доступа к питательным веществам почвы, обусловленное симбиозом, было продемонстрировано недавно М. ван дер Хайденом и его коллегами на примере сообществ лужаек (рис. 13.27). Они добавили в богатую видами модельную экосистему микоризный посевной материал на стерильном субстрате, выращенный из отдельных спор разных генотипов Glomus. В зависимости от генотипа микоризы другие виды растений доминировали или подавлялись. Некоторые виды растений исчезали, если не получали «своего» генотипа. Наличие определенных микоризных грибов, следовательно, определяло, может ли прокормиться данный вид или нет, и таким образом обусловливало растительное биоразнообразие этих лужаек. Могут ли эти результаты отразиться на общеупотребительной практике выращивать экспериментальные растения на определенных участках, вызывает сомнение.
Рис. 13.27. Влияние микоризы на рост растений (биомасса ± стандартная ошибка). В маленькую модельную экосистему с типичными видами бедных фосфором центрально-европейских малопродуктивных лужаек на известняках были инокулированы на стерильном естественном субстрате разные генотипы (изоляты) образующего эндомикоризу рода Glomus (А, В, С, D). В зависимости от изолята выиграли разные виды растений. Без микоризы (0) выигрывает единственный безмикоризный вид, Сагех flacca. Доминантный Bromuserectus не реагирует на тип изолята. Видоспецифичное микоризообразование, соответственно повышающее доступность питательных веществ, — решающий фактор биоразнообразия
13.6.5. Азот и фосфор в глобальном аспекте
В крупных пространственных масштабах и на протяжении длительного времени продуктивность Земли лимитировалась в основном тремя факторами: температурой, водой и фосфором, если солнечную радиацию и концентрацию СО2 в атмосфере рассматривать, как заданные величины. Хотя большее количество водяных паров (больше облаков) и может задерживать солнечное излучение, современное антропогенно обусловленное повышение СО2 может потенциально увеличивать связывание углерода, но лишь ненамного выше той границы, которую устанавливает потребление фосфора. Фосфор не только во многих наземных экосистемах, но и в большей части океанов является важнейшим фактором, влияющим на продуктивность. За исключением отдельных районов Южного океана, где играет роль дефицит железа, достаточная доступность фосфора, прежде всего в Тихом океане, служит предпосылкой для того, чтобы морские цианобактерии могли выдавать в систему азот, что в свою очередь служит предпосылкой для связывания углерода. П. Фальковски с сотрудниками установили, что перенос пыли с суши (дополнительная доставка фосфора) в конечном счете является движущей силой продуктивности океанов, причем далеко от побережий.
Чем суше, тем самым более «пыльно» на суше, со стороны которой дуют ветры, тем больше океаническая продуктивность. Так как большая часть суши в ледниковое время не покрывалась водой, можно даже объяснить низкую в это время концентрацию СО2 в атмосфере (190 ppm) с точки зрения круговорота фосфора и связанной с ним продуктивностью океанов. Если астрономические расчеты для ледниковых периодов достоверны, то нельзя исключить, что подобные взаимодействия моря с сушей были тогда значительно интенсивнее.
На региональном уровне в прибрежных водах океана, на молодых аллювиальных отложениях (и в целом на молодых почвах) обеспечение фосфором большей частью лучше; на зрелых почвах, на участках земной поверхности с малой тектонической активностью (например, в Австралии) или вообще на сильно выветренных почвах оно часто плохое, причем, как выше упомянуто, нельзя заметить какие-либо симптомы фосфорного дефицита в растительности. Растения реагируют на него развитием многолетних листьев, и при таких условиях в особенно большой степени зависят от микоризы. Симбиоз растений и грибов такой же древний, как и сама жизнь на суше, и с тех самых пор снабжение растений фосфором могло решающим образом определяться почвенными грибами.
С азотомдело обстоит принципиально по-другому, так как он в неограниченных количествах присутствует в воздухе, и сколько его может быть связано в экосистеме, вопрос только микробиологической активности (но для этого опять-таки необходимы фосфор и углерод). В настоящее время антропогенное высвобождение растворимых соединений азота достигает таких размеров, что, по расчетам П. Витоушека с коллегами, в 1987 г. оно уже перекрыло объемы естественной фиксации азота. Густонаселенные районы Земли (также и в Европе) сейчас с экологической точки зрения являются областями перенасыщения азотом, даже если это антропогенное поступление из атмосферы в полуестественную растительность составляет 15 — 25 кг N • га/год (это «только» 1/10 — 1/20 обычной нормы расхода азотных удобрений в интенсивной системе сельского хозяйства).
13.6.6. Кальций, тяжелые металлы, соли
Кроме основных питательных веществ — фосфора и азота — на рост и существование растений сильное влияние оказывают другие минеральные компоненты почвы (см. 6.2). Наиболее известно действие карбоната кальция, который благодаря своему сильному влиянию на pH почвы (см. 12.5.2.3; буферизация), а также путем взаимодействия с другими свойствами почвы (доступность элементов, микориза) воздействует на рост непрямым образом. Но часто констатируется различное поведение по отношению к самому иону Са2+. Хотя многие виды могут произрастать и на бедном, и на богатом кальцием субстрате, во втором случае кальций осаждается в виде не влияющего на физиологию клетки оксалата (например, Silene и другие Caryophyllaceae). Настоящие кальцефиты (кальцефильные растения) выдерживают в клеточном соке большие количества растворенного кальция (например, Gypsophila, как исключение из всех Caryophyllaceae). Избегающие кальция (кальцефобные) растения, например, белоус (Nardus stricto), сверхчувствительны к Са2+. Флора и растительность богатых и бедных кальцием местообитаний всегда резко различаются (кальцефиты и силикофиты)
Два вида вересковых, произрастающих в Альпах у верхней границы леса, Rhododendron ferrugineum и R. hirsutum в нормальных условиях встречаются: первый — на силикатных породах с сильнокислыми почвами (pH 4,0 — 6,0), а второй — на карбонатных породах с почвами от слабокислой до слабощелочной реакции (pH 5,8 — 7,2). В области их контакта и на переходных место обитаниях (pH 5,4 — 6,4) могут быть образованы сообщества из гибридов обоих видов.
На богатых кальцием породах формируются как первый этап почвообразования разнообразные гумусно-карбонатные почвы (рендзины) с профилем типа А — С (см. 12.5.2.3). На силикатных и кварцевых породах им соответствуют гумусно-силикатные почвы (ранкеры). Первые обладают высокой степенью буферности к подкислению, последние склонны к подкислению и вымыванию оснований. В Центральной Европе для карбонатов характерны сухие лужайки с Sesleria albicans (= S. varia) и Teucrium montanum, для силикатных почв — такие же лужайки, но с Seduni acre и Scleranthus perennis. В поясе альпийских лугов викарируют сообщества Elyno-Seslerietea на богатых известью и Caricetea curvulae на бедных известью почвах. Наконец, особая флора и растительность (например, с характерным гипсофильным лишайником Acarospora nodulosa) развивается локально на карбонатно-сульфатных почвах с гипсом и (или) ангидритом.
Локальное насыщение потенциально токсичными соединениями тяжелых металлов, таких, как медь, кобальт, никель, марганец, уран, алюминий, магний, цинк, селен и др., в значительной степени ограничивает рост растений, кроме сильно ограниченного набора экофизиологически специализированных, которые обладают толерантностью к таким соединениям и даже иногда аккумулируют их (см. 6.2.2.4; там также указано их значение в качестве индикаторных растений). В связи с этим заслуживают упоминания сильно выделяющаяся на окружающем фоне растительность на серпентинах (силикат магния с Al, Fe и Ni) и галмейная растительность (на цинковых рудах; см. 6.2.2.3).
Насыщение легкорастворимыми солями (особенно NaCl, Na2SО4, Na2CО3, но также и соответствующими соединениями К и Mg) в районах морских побережий и ландшафтов аридных бассейнов в континентальных областях оказывает решающее воздействие на жизнь растений. Это уже многократно было показано при обсуждении морфологических, анатомических и физиологических особенностей галофитов — растений засоленных местообитаний (см. 6.2.2.4).
Самая высокая резистентность к солям развита у водорослей и лишайников литоральной зоны морских брызг; они переживают как высыхание концентрированного соляного раствора, так и выщелачивание дождевой водой. Напротив, растения пресных вод (гликофиты) страдают даже от небольшого количества солей натрия (около 50% морской воды). Факультативные галофиты (например, морская астра Aster tripolium) могут переносить такие концентрации вполне нормально. Облигатные галофиты вообще достигают своего оптимального роста при соответствующих концентрациях солей (например, Salicomia при 75 — 100% морской воды).
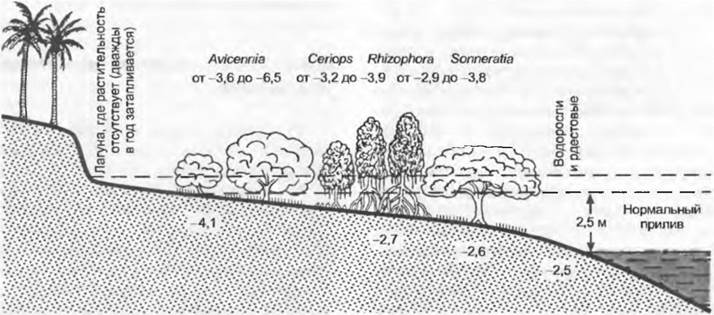
На гумидных побережьях концентрация солей в почве снижается от моря к суше; этому соответствует убывающая солеустойчивость облигатных и факультативных галофитов, которые сменяют друг друга по этому градиенту (например, на побережье западной Швеции; рис. 13.28). Но там, где имеются аридные времена года, даже кратковременно пропитывающаяся морской водой краевая зона у кромки суши особенно обогащена солями, потому что здесь почвенные растворы сильнее концентрированы из-за испарения в сухой период. Такие условия господствуют, например, в манграх (рис. 13.29; см. бокс 4.5, раздел 15.2.16), где почвы от открытого моря в сторону лагуны становятся более насыщенными солями и виды соответственно этому распределяются в сторону возрастания солеустойчивости.
Рис. 13.28. Профиль растительности вдоль градиента засоленности на низком морском берегу в западной части Швеции. Обилие разных видов на профиле длиной 45 м от кромки моря до пастбищного луга, лишь немного испытывающего влияние засоления. На графике сверху вниз заметны 3 отличающиеся друг от друга экологические группы. Максимальная разница высот вдоль профиля составляет 1 м, колебание уровня моря в течение большинства месяцев года не превышает 0,5 м, только один раз осенью оно достигает верхней точки профиля
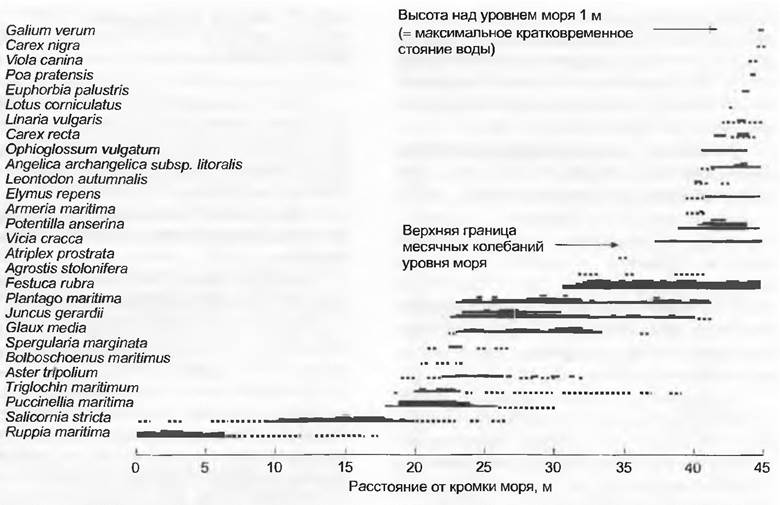
Рис. 13.29. Зонирование мангров на восточноафриканском побережье. Благодаря периодическому высыханию в самой отдаленной от моря точке профиля выявлена наибольшая концентрация солей. Концентрация солей в почвенном растворе 10 см ниже верхней границы почвы (ила) и в выжатом соке листьев дана как осмотический потенциал в МПа. Четыре мангровых рода занимают характерные зоны вдоль градиента, показывающего колебания уровня моря