БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
13. РАСТЕНИЯ В ЖИЗНЕННОМ ПРОСТРАНСТВЕ
13.7. Рост и баланс углерода
Успех вида растения при заселении жизненного пространства зависит в конечном счете от его способности создать и поддерживать устойчивую популяцию. Для этого необходимы четыре условия:
✵ быть способным к росту и продуцированию биомассы при данном состоянии ресурсов;
✵ выдерживать типичные для данного местообитания стрессовые ситуации;
✵ переносить нарушения со стороны фитофагов, патогенных организмов или механических факторов;
✵ успешно воспроизводиться.
Энергия роста растения определяет шансы для его регенерации после стресса или нарушения, преуспевания в конкуренции за ресурсы с другими видами, а также воспроизведения полового потомства или клональных единиц распространения. Поэтому пониманию процессов, непосредственно влияющих на рост, представляется приоритетная роль в экологии растений. После рассмотрения экологии роста следуют разделы, посвященные продукции биомассы и углеродному балансу в экосистеме. Предпосылкой к ним служат физиологические и биохимические основы фотосинтеза и дыхания (см. 6.5, 6.10).
13.7.1. Экология фотосинтеза и дыхания
Фотосинтетическое связывание СО2 и выделение СО2 при дыхании составляют основу круговорота углерода на Земле (см. 13.7.6). Оба процесса помимо внутренних факторов сильно зависят и от факторов среды. Как фотосинтез, так и дыхание, в том числе дыхание, происходящее при процессах разложения после гибели растения, перемещают сходные количества углерода, поэтому в углеродном балансе им придается одинаковое значение. Тем не менее о фотосинтезе известно очень много, а о дыхании сравнительно мало. Дело, вероятно, в том, что фотосинтез происходит в хорошо определяемых и легкодоступных органах (как правило, в зеленых листьях), в то время как в дыхании, напротив, участвуют все органы растения, в том числе и подземные, и оно во многом зависит именно от типа органа.
Поскольку фотосинтез инициируется вектором света, а точнее плотностью потока фотосинтетически активных фотонов (РFD), имеет смысл относить его значения к проекции поверхности ассимилирующих органов (листовой поверхности). Интенсивность дыхания R не зависит от направленных величин, и поэтому предпочтительнее выводить ее из количественных показателей тканей (чаше всего из сухой массы). Для определения интенсивности дыхания листьев, во всяком случае на экосистемном уровне, выбирается также отношение к поверхности. Выбор величины отношения имеет решающее влияние на результаты и вытекающие отсюда окончательные выводы. Связыванию СO2 противостоят одновременно потери СO2 через световое дыхание и митохондриальное дыхание в тканях листа. Наблюдателю, как правило, доступен только чистый результат — степень нетто-фотосинтеза А (ассимиляция). Поскольку митохондриальное дыхание на свету частично подавлено и для поддержания аппарата фотосинтеза при сильно колеблющемся количестве излучения важные функции приобретает световое дыхание (см. 6.5.6), не имеет экологического значения и не рекомендуется измеренную в темноте интенсивность дыхания и даже «потери» на световое дыхание добавлять к нетто-фотосинтезу и указывать брутто-фотосинтез.
Большая трудность в охарактеризовании зависимостей А и R от внешних факторов состоит в том, что эти зависимости очень варьируют во времени и в свою очередь зависят от других переменных. Таким образом, для вида не имеется определенной «нормы реакции», а целая совокупность таких функций и их быстрая сменяемость вслед за внешними условиями «регулируют» функциональную зависимость. Так как общие принципы были представлены в разделах 6.5 и 6.10, в этом разделе ограничимся экологически значимыми взаимодействиями (рис. 13.30; для СO2 см. рис. 13.45).
Рис. 13.30. Интерактивная зависимость фотосинтеза листьев от света, температуры и содержания СO2 у С3-растений (схематично): А — изменение зависимости от плотности потока фотонов (PFD), если вместо 360 ppm СO2 предлагается 600 ppm LKP световой компенсационный пункт при 360 ppm СO2; В— изменение зависимости от температуры при постепенном понижении PFD (до 5 % от PFDsat), нормальном содержании СO2. Кривая зависимости от С02 имеет форму, идентичную кривой зависимости от PFD (ср. рис. 13.45). Дальнейшие пояснения см. в тексте
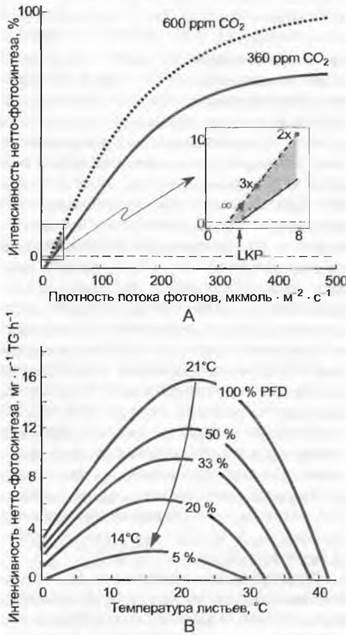
Функции на рис. 13.30 и 13.45 охарактеризованы через пограничные значения и частные отрезки кривых. Ход по абсциссе называется в случае зависимости РFD и СO2компенсационными пунктами света и СO2 (оба даны для солнечных листьев С3-растений и случайно совпадают по значениям, 20 °С, около 20 — 30 мкмоль фотонов • м-2 • с-1, или ррm СO2). Линейный начальный отрезок (initial slope) обеих этих функций насыщения обозначается как эффективность использования квантов (от англ. quantum use efficiency, QUE) и эффективность потребления СO2 (СO2 uptake efficiency, CUE). Линейный подъем кривой PFD — это лимитирование нормы путем
световой реакции фотосинтеза (регенерация акцепторов СО2, RubР), плато кривой (насыщение) — лимитирование темновой реакцией (связывание СO2, карбоксилирование). На кривой СO2 все как раз наоборот, а именно начальный подъем показывает лимитирование темновой реакцией, плато — лимитирование путем подготовки эквивалентов редукции (световая реакция). Концентрация СO2 и РFD взаимодействуют таким образом, что с возрастающей концентрацией СO2 компенсационный пункт света сдвигается влево (в сторону нуля) и насыщение РFD для А наступает при более высоких значениях РFD. С экологической точки зрения это означает, что лучшее использование света происходит при повышенном содержании СO2 в воздухе, особенно в тени (см. 13.7.6). Световой компенсационный пункт и световое насыщение весьма адаптивны. Теневые растения компенсируют при РFD <10 (до 3) мкмоль фотонов • м-2 • с-1, а насыщаются при 100 — 150 мкмоль фотонов • м-2 • с-1 (примерно 5 — 8 % от полного света полуденного солнца). Большинство светолюбивых растений достигают 90 %-го насыщения РFD при его значениях 400 — 600, при особой толщине и прочности листьев насыщение наступает только при РFD >1000 мкмоль фотонов • м-2 •с-1 (показатели для С4- и САМ-растений см. 6.5). Зависимость А от СO2 показывает также поведение при акклиматизации, продолжительной экспозиции в повышенной концентрации СO2, и тем не менее она не остается постоянной (см. рис. 13.45).
Температурная зависимость А — комплексная функция из ускоряющих (функция карбоксилазы RubisСО) и тормозящих воздействий (функция оксигеназы RubisСО + митохондриальное дыхание), в результате чего строится кривая нормального распределения. Hижние и верхние пограничные значения (А = 0) и оптимум (Аmах) типичны для определенных климатов. В умеренных, бореальных и арктоальпийских регионах при полной активности листьев температурные рубежи морозоустойчивости совпадают с температурными рубежами А (-2... -8 °С, в горах часто около -5 °С). Это означает, что А прекращается тогда, когда листья непоправимо повреждаются
морозом до состояния гибели. Верхний температурный рубеж находится между 40 (адаптированные к холоду растения) и 45 °С (адаптированные к теплу), т. е. также всего несколько градусов отделяют его от летального перегрева (см. 13.3.2). Оптимум колеблется между 15 °С (крайне адаптированные к холоду растения) и почти 30 °С (адаптированные к перегреву растения). Еще более низкие оптимумы отмечаются у криптогамных растений холодных биомов.
Оптимум может сдвинуться при акклиматизации за короткий промежуток времени (несколько дней) на 5 К (и больше). Определяющим для адаптации является фактический (микро) климат. Поскольку сообщества горных растений со стелющимися побегами на солнце получают относительно высокие температуры, неудивительно, что их фотосинтетический температурный оптимум очень близок к растениям низменностей (20 — 25 °С). В умеренных и холодных районах температурный оптимум для А очень широкий (90 % А находится в области>10 К). У тропических растений А = 0 уже на границе холода (3 — 7°С) и оптимум сравнительно узкий.
Большое экологическое значение имеет соотношение освещенности и температуры. Обсуждавшиеся выше зависимости РFD и Т действительны для оптимальной температуры и, соответственно, светового насыщения. Хотя такие функции зависимостей можно получить в лаборатории, в природе они проявляются весьма ограниченно. Дело в том, что в течение периода роста (если пренебречь исключениями) при высоком РFD тепло, а при сильно ограниченном освещении прохладно. На это хорошо настроен фотосинтетический аппарат растений. Интерактивное действие РFD и Т таково, что при низком РFD температурный оптимум для А также достигается при очень низких температурах (например, 12 °С вместо 22 °С). При малом освещении растения уже при относительно низкой температуре достигают наиболее возможной при таких лимитирующих условиях интенсивности фотосинтеза. Поэтому фотосинтез почти никогда не лимитируется температурой, но часто — освещенностью. Измеренный при световом насыщении, оптимальной температуре и «нормальной» концентрации СO2 листовой фотосинтез обозначается сейчас как максимальная интенсивность фотосинтеза Аmax. Ранее использовавшееся для этого понятие емкости фотосинтеза Аcар теперь употребляется для обозначения максимально возможного фотосинтеза при насыщении СO2.
Охватить дыхание, близкое к реальному, и найти удобные условные величины, которые сами по себе не дают расхождений, — труднейшие задачи функциональной экологии. Различают три вида митохондриального дыхания: поддерживающее, или обменное, дыхание (англ. maintenance respiration; далее речь пойдет только о нем), так называемое дыхание роста (от англ. growth respiration), связанное с новообразованиями в тканях, и корневое дыхание при усвоении питательных веществ (англ. nutrient uptake respiration). Интенсивность поддерживающего дыхания R сильно зависит от активности тканей, но она перекрывается соотношением сухого веса и содержания углерода в ткани, т. е. своей пространственной плотностью (чем плотнее ткань, тем меньше расчетные интенсивности дыхания на протопласт), которая со своей стороны влияет на концентрацию азота в расчете на сухой вес (см. 13.6.3). Цветки, тонкие корни и листья (в темное время) имеют высокие значения R на грамм сухого веса, стебли и толстые корни — меньше, а древесные структуры или запасающие органы — очень маленькие. На основе содержания азота (как меры протеина) эти различия часто исчезают или становятся очень небольшими. За главное правило можно принять то, что активное растение примерно половину своей ежедневной ассимиляции СO2 отдает на дыхание и что «мягкие» ткани более всего способствуют этим потерям при пересчете на инвестируемый углерод (сухой вес).
Сравнение респираторной активности разных органов или разных видов растений, или даже особей одного и того же вида из различных условий обитания всегда включает ее воздействие на расчетные параметры, т. е. внешняя среда или развитие столь сильно влияют на специфическое для клеток дыхание, что реальные величины часто не соответствуют расчетным параметрам (например, объемный вес ткани). Наконец, поддерживающее дыхание чрезвычайно чувствительно реагирует на
все мыслимые воздействия, включая внутренние перестройки растения в ходе его развития. ассимиляционную деятельность или ранее перенесенный стресс, и особенно на деструктивные вмешательства, например, на выкапывание корней. Поскольку на корни у многих растений приходится большая часть «общего расхода» на дыхание, особая трудность заключается в том, что установить дыхание собранных в интактную ризосферу корней можно только непрямым образом (например, с помощью изотопов С) и только с очень большими затратами.
Важнейшая для дыхания активных растений климатическая переменная — температура (см. 6.10.3.6). В целом R увеличивается с поднятием температуры (как норма большинства энзиматических процессов) примерно вдвое, если температура в средней области (например, 10 — 20 °С) увеличивается в пределах 10 K(Q10 = 2). С экологической точки зрения воздействие температуры на дыхание характеризуется этим явно недостаточно. То, что здесь было описано, всего лишь частное событие. Едва ли какой-либо другой жизненный процесс реагирует так на изменение условий, как митохондриальное дыхание. Основатель современной экофизиологии, ориентированной на исследования в природе, в Германии О. Штокер был, вероятно, первым, кто с удивлением установил: приспособления дыхания к господствующей температуре настолько значительны, что могут компенсировать даже самые широкие амплитуды температур.
Штокер сравнивал летнее (теневое) дыхание ивы (Salix) в ее естественном местообитании в Гренландии с дыханием ветвей деревьев во влажно-тропическом лесу Индонезии и не нашел между ними никаких различий. Сходные наблюдения проводил А. Писек, который исследовал дыхание хвойных деревьев в долине и на верхней границе леса в Тироле. Однако если измеряют дыхание растений или тканей из теплых и холодных областей, тотчас после отбора проб при одних и тех же температурах (не при тех температурах, которые господствуют в их естественных местообитаниях) видно, что интенсивность дыхания адаптированных к холоду растений всегда явственно выше. В литературе это часто неправильно интерпретируется как «более интенсивное дыхание в холодных областях». В действительности дыхание в холодных областях скорее ниже, прежде всего из-за холодных ночей. Вывод о попытке растений встретить неблагоприятные температурные условия повышением специфической активности можно сделать из того, что число митохондрий с понижением температуры местообитания увеличивается. Дыхание нельзя рассматривать только как нагрузку на углеродный баланс, оно представляет собой жизненно необходимый процесс («потребность»).
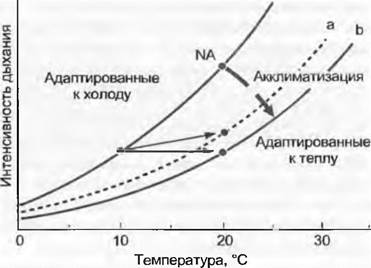
Акклиматизационное приспособление дыхания к новым температурам окружающей среды (рис. 13.31) протекает относительно быстро (от одного до нескольких дней), но не всегда окончательно, как на этом примере. График наглядно показывает, что неуместно на основе известной кратковременной температурной зависимости метаболических процессов составлять прогнозы фактической интенсивности процессов в условиях постоянно меняющейся в термическом отношении среды (например, глобальное потепление климата). Такие прогнозы должны учитывать акклиматизационные возможности растений.
Рис. 13.31. Зависимость дыхания от температуры до и после акклиматизации. Жирная стрелка показывает направление акклиматизации к новой для растения более высокой температуре (20 °С вместо 10 °С), каждая из этих кривых — кратковременную реакцию дыхания на различные температуры (в пределах продолжительности эксперимента, 1 — 2 ч) у групп растений одного и того же вида, адаптированных к холоду или к теплу. В данном примере акклиматизация к теплу либо частичная (а), либо полная (b). В случае b это означает, что помещенная в тепло группа растений дышит при новой температуре после акклиматизации с той же интенсивностью, как и до этого в своем более холодном жизненном пространстве. При идентичных температурах акклиматизированная к 20 °С группа дышит с меньшей интенсивностью, чем группа, акклиматизированная к холоду, что, однако, несущественно в экологическом отношении. NА маркирует теоретическую интенсивность дыхания при новой температуре роста, если никакой акклиматизации не состоялось. Кривая а представляет собой наиболее частый случай
13.7.2. Экология роста
Рост растения в конечном счете представляет собой баланс между приходом и расходом, выраженный в сухом весе, т. е. сумму ассимилированного углерода минус сумму всех затрат на дыхание и прочие потери. Интенсивность нетто-связывания углерода всем растением в определенный промежуток времени зависит от таких факторов, как:
✵ интенсивность фотосинтеза на единицу листовой поверхности (интегрированная из всех листьев);
✵ отношение общей листовой поверхности к общей биомассе растения (от англ. leaf area ratio, LAR);
✵ дыхание всех органов (для разных органов очень различное);
✵ экспорт углерода (например, на симбионтов);
✵ активность отложения углерода (структурный рост или накопление).
Каждый из этих 5 факторов со своей стороны находится в зависимости от многочисленных внешних и внутренних воздействий. Невозможно прогнозировать связывание углерода или рост только по одному из этих факторов. Это простое осознание противостоит доминирующему в течение долгого времени представлению, что рост есть непосредственное следствие листового фотосинтеза и ограничен только интенсивностью его деятельности. Такое злополучное ограничение перспективы имело следствием то, что о фотосинтезе в природе известно очень много, но о 4 других детерминантах роста и их зависимости от внешней среды — очень мало или почти ничего. Хотя все эти 4 дополнительных фактора в принципе могут достаточно эффективно влиять на нетто-продукцию С, из них 3 (кроме последнего) варьируют у определенного вида растений только в некоторых морфологически обусловленных границах. Активность отложения С, регулируемая другими доступными ресурсами (собственно прирост массы), напротив, чрезвычайно вариабельна и в большинстве случаев (за исключением дефицита света) представляет собой непосредственную силу, управляющую ассимиляцией углерода растением. Это становится ясным из простого факта, что фотосинтез беспрепятственно протекает до тех пор, пока продуцируемые ассимиляты находят употребление где-либо в самом растении, т. е. могут быть инвестированы. В ином случае «производство продукции» должно сейчас же прекратиться, поскольку транспортные пути быстро «закупорятся», и, соответственно, хлоропласты будут переполнены ассимилятами (торможение конечного продукта).
С экологических позиций это центральный пункт понимания роста растений.
Высокая активность отложения углерода индуцирует высокую интенсивность фотосинтеза, незначительная активность снижает интенсивность. Если отделяют накопленный углерод от растения, например, растущие картофельные клубни, или яблоки, интенсивность фотосинтеза в листьях падает. Если растение лишается части листьев, интенсивность фотосинтеза в оставшихся листьях повышается.
Активность отложения углерода в растении зависит от наличия ресурсов (воды и питательных веществ) в почве, от температуры и стадии развития самого растения, которая в свою очередь определяется двумя предыдущими факторами и многочисленными другими (например, фотопериодом). Многочисленные литературные источники подтверждают, что активность отложения углерода реагирует на все окружающие воздействия, за исключением света, что она чувствительнее, чем фотосинтез в листьях. Ростовые процессы (клеточное деление, увеличение объема и дифференциация клеток) реагируют задолго до того, как фотосинтез сколько-нибудь значительно ограничивается недостатком воды, дефицитом минерального питания или низкими температурами. Поэтому не будет преувеличением утверждать, что в большинстве случаев, кроме опять-таки лимитирования света (и естественно ситуаций после потери листьев), рост, а значит потребность в ассимплатах управляют фотосинтезом, а не наоборот.
Просто удивительно, как давно это известно и как мало это нашло применения в общебиологических парадигмах. В 1864 г. Е. Краус опубликовал в журнале «Флора» результаты классического эксперимента, который он провел в лаборатории Юлиуса Сакса в Вюрцбурге. Тогда еще фотосинтетическую активность листьев определяли, наблюдая за образованием газовых пузырьков погруженными в воду побегами. Сотрудникам Сакса уже было известно, что интенсивность фотосинтеза время от времени не контролируется потребностями в ассимилятах и скоростью экспорта последних, и это ведет к усиленному образованию ассимиляционного крахмала, что можно было доказать с помощью йода и калий-йода. Краус поставил в высшей степени современный вопрос — на какой из двух процессов сильнее воздействуют низкие температуры: фотосинтез (образование газовых пузырьков) или использование ассимилятов (аккумуляция крахмала)? Ответ ему дали кубики льда, брошенные в водяную баню: в охлажденной водяной бане, образование пузырьков едва снижалось, количество крахмала увеличивалось по сравнению с теплой водой на контроле. Несмотря на все справедливые возражения против такого эксперимента с современных позиций, ход размышлений и наблюдения отлично иллюстрируют дилемму: что лимитирует усвоение углерода — его отложение или источник питания? Новые результаты для растений прохладных регионов соединили все это в единую картину: при 0°С листовой фотосинтез еще достигает около четверти максимально возможной продуктивности и только при температуре около -6°С прекращается (см. рис. 13.30, В); напротив, рост, т. е. активность отложения при температуре чуть выше 0°С прекращается, а при температуре ниже 5 °С происходит очень медленно. Соответственно, в растениях холодных биомов аккумулируются углеводы (крахмал, фруктоза), не связанные с определенными структурами, а за более длительный период и липиды. Полная аналогия имеется и в сезонном сухом периоде, так как рост также гораздо более чувствителен к дефициту влаги, чем фотосинтез.
Дыхание часто реагирует и на воздействия внешней среды (особенно на температуру) в гораздо большей степени, чем фотосинтез, частично сдерживая активность отложения (дыхание роста), однако определяется с большим трудом из-за специфики в разных органах. Так, практически невозможно в реальных условиях измерить корневое дыхание, как правило, самый большой источник потерь углерода. Как только тонкие корни вычленяются из своей микросреды и отделяются от симбионтов, изменяется и их дыхание.
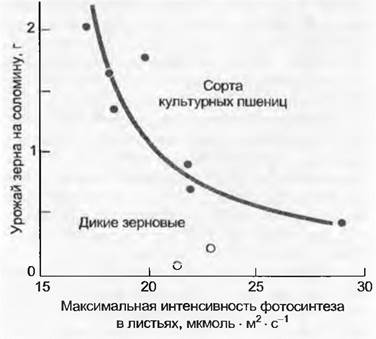
Значение отложения углерода и вместе с тем тип инвестиций ассимилятов — центральный вопрос не только экологии, но и сельскохозяйственной науки. Не говоря о земледельческих мероприятиях, повышение урожая зерновых культур сводится, в сущности, к управлению ассимиляционными потоками в сторону желаемого конечного продукта, а не к более высокой активности листьев (рис. 13.32). Выдающиеся исследователи неоднократно доказывали, что повышенный урожай зерна не связан с повышением интенсивности фотосинтеза в листьях.
Рис. 13.32. Максимальная интенсивность фотосинтеза Аmах в листьях и урожай зерна. Классический пример факта из практики сельского хозяйства о том, что продуктивность фотосинтеза на единицу листовой поверхности не определяет урожая. На этом примере для разных сортов пшеницы связи урожая с фотосинтезом даже негативны. Листья двух дикорастущих видов рода Aegilops имеют нисколько не меньшие значения Аmaх, чем современные высокоурожайные сорта (каждая точка измерения представляет собой среднее из многих индивидуумов сорта (вида). Определяющие урожай факторы среди прочих — это морфология стебля и колосьев, распределение сухого вещества (harvest index), адаптация листьев к слабому освещению (благодаря чему они еще способны продуцировать даже в нижних слоях сообщества), длительность жизни листьев и внутренняя ритмика развития (замедленное старение) всего растения
Чистый подсчет прибыли за короткий промежуток времени сам по себе мало может способствовать в экологическом отношении пониманию ростового процесса. Еще труднее это сделать при более длительном периоде наблюдений, когда встает вопрос о длительности функционирования всех органов и тканей (вопросы амортизации). «Прибыль», которую получает лист, образуется из баланса между живой продукцией фотосинтетически связанного СO2 и затратами на построение самого листа, после оттока переотложенных веществ перед его гибелью. Пренебрегая вопросами издержек, урожай есть продукт работы за единицу времени, умноженной на срок ее проведения, обе величины одинаково значимы. Хотя по соотношениям излучения и содержания протеина (содержания азота) на единицу листовой поверхности можно дать относительно хороший прогноз интенсивности фотосинтеза (см. рис. 13.20), измерения связаны с большими затратами. Напротив, относительно длительности жизни листьев в литературе можно найти только очень скудные данные, хотя их можно легко получить и без технических затрат.
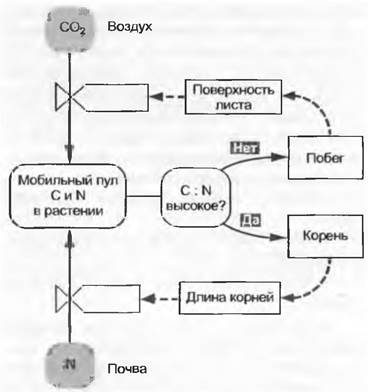
Рост определяется тем, куда и в какой форме инвестируются ассимиляты углерода. Переотложение фогоассимилятов в новые листья выражается в сложных процентах (дневные проценты!). Инвестиция в зеленые стебли с точки зрения баланса может быть нейтральной, в то время как затраты на запасающие органы более низкие, а на тонкие корни — более высокие (прежде всего расходы на дыхание). Такие «решения» не «свободны», они находятся под воздействием трех движущих сил: плана строения, т. е. унаследованного морфотипа; типа развития и соответственно изменения основных пунктов инвестиции в течение жизни; внешних факторов. Ассимиляционные потоки управляются в пределах границ, обусловленных строением и развитием, ассортиментом ресурсов (рис. 13.33; см. 13.7.3): много света— мало листьев; мало света — много листьев; большой ресурс азота — большая масса листьев и маленькая масса корней и т. д.
Рис. 13.33. Простая модель связи углеродного и азотного бюджетов в растении. Исходя из вопроса: имеется ли лимитирование углерода (поступившего, но еще не связанного структурно в фотоассимиляты) или азота (доступные азотные соединения), т. е. мобильного «строительного материала» в отношении C/N, инвестиции их направляются либо в листья в большей степени, либо, напротив, в корни. Сплошные стрелки символизируют потоки массы, пунктирные указывают на воздействия
У разнотравья, злаков, низких кустарников и всходов деревьев некоторые из этих детерминантов роста можно довольно просто установить, используя весы и сушильный шкаф. Они по меньшей мере в такой же степени важны, как и получение СO2, соотнесенное с листовой поверхностью. Непонятно, почему при скудных средствах на исследования (также и в странах со спросом на исследование) прежде всего вводится в действие дорогая газоизмерительная аппаратура, когда можно провести ничуть не менее полноценные исследования остальных параметров роста гораздо более простыми средствами.
Исходя из объема инвестиций углерода в листья, растение может получить больше или меньше углерода, при этом регулируется фотосинтетический «доход» всего растения, без изменений специфики продуктивности фотосинтеза. Кроме света важнейшие параметры, определяющие рост, следующие: температура, доступная вода и питательные вещества. Они определяют, какое количество фотосинтеза может «позволить себе» все растение или сколько ассимилятов вообще может быть «инвестировано». Эта картина мобилизации ассимилятов по потребности находится в противоречии с господствующими до сих пор представлениями, а именно, что рост определяется только имеющимся запасом ассимилятов, а значит продуктивностью фотосинтеза. Последнее происходит только в том случае, когда все остальные факторы роста не лимитированы. Это может быть только при особых ситуациях, в условиях интенсивного сельского хозяйства, или при глубоком затенении, когда связывание СO2 становится единственным определяющим компонентом. Характерно, что многие вечнозеленые растения в течение сухих периодов накапливают резервы (англ. stored growth — запасный рост), несмотря на ограниченное усвоение СO2, и в это время, как правило, не испытывают «голода». С наступлением дождей эти резервы мобилизуются для распускания новых побегов.
Несмотря на то что у С4-растений чаще всего продуктивность фотосинтеза выше, чем у С3-растений, их сельскохозяйственная продукция при высокой влажности почвы далеко не всегда более высокая. Они получают преимущество при малом поступлении воды, прежде всего во время засух. Благодаря компенсаторному росту корней дефицит воды и питательных веществ могут временно не проявляться, но на экосистемном уровне этому устанавливаются естественные границы. Как показывают новейшие исследования в обогащенном СO2 воздухе, стимулирование степени фотосинтеза большим количеством углекислого газа дает очень небольшое увеличение роста, поскольку при таких экспериментах ограничение роста не преодолевалось искусственным возрастанием других ресурсов.
13.7.3. Функциональный анализ роста
Функциональный анализ роста исходит из нескольких основных параметров, которые преимущественно касаются того, как ассимиляты распределяются в растении по различным органам и как они в них размещены. Окружающая среда может влиять на процессы распределения таким образом, что ассимиляты предпочтительно инвестируются там, где имеется их недостаток: при сухости почвы — в корни, при недостатке света — в листья и т. д. Это проявляется в сухой массе органов по отношению к общей сухой массе растения (рис. 13.34). В английской номенклатуре для этого применяются следующие понятия:
✵ LMF — leaf mass fraction (листовая часть);
✵ SMF — stem mass fraction (масса всех побегов);
✵ RMF — root mass fraction (корневая часть).
Аналогично в качестве фракций биомассы могут быть обозначены репродуктивные органы, запасающие органы и др. В последнее время умножаются доказательства того, что эти модели распределения биомассы не всегда соответствуют впервые описанному Р. Броуером «функциональному равновесию» и распределение сухого вещества очень сильно реагирует на факторы окружающей среды. Как правило, к этой проверенной модели добавляются еще физиологические и морфологические коррекции (например, регулирование насыщения азотом и продуктивность фотосинтеза в листьях, кутикула и размеры листьев).
Рис. 13.34. Распределение биомассы и жизненные стратегии растений. В фотосинтезирующих органах находятся очень различные доли от общей биомассы. Здесь приведена leafmass fraction (LMF), как отношение листовой фракции биомассы к общей биомассе (% сухой массы, затемненная плоскость) взрослых растений различных морфотипов, т. е. разных жизненных стратегий

Fla уровне листа и корня нас интересует «стоимость» биомассы, необходимая для создания функционального единства. Так как функция листьев заключается прежде всего в абсорбции света, она описывается лучше через листовые поверхности. У корней это интенсивность пронизывания почвы, следовательно, длина тонких корней, соответствующая единице биомассы. Именно поверхность корней вступает в контакт с почвой, правда, качество (активность) этой поверхности меняется с возрастом корней. В отношении к корневой поверхности нельзя занижать оценку роли очень активных абсорбирующих тонких корней и переоценивать роль уже не абсорбирующих (преимущественно проводящих) толстых старых корней. Поэтому независимо от толщины часто используют длины корней как функциональную меру. Соответствующие параметры обозначаются как:
✵ SLA (specific leaf area — специфическая листовая поверхность, квадратный метр поверхности листьев на грамм их сухого веса, или для наглядности — дм2 • г-1 или м-2 • кг-1);
✵ SRL (specific root length — специфическая длина корней; метр длины корней на грамм их сухого веса).
Вместо SLA часто употребляют обратное этому показателю значение LMA (leaf mass per area). Из сочетания долей массы органов и их специфической «стоимости» получают два важных уравнения функционального анализа роста:
LAR = LMF • SLA
(leaf area ratio — общая поверхность всех листьев по отношению к общей биомассе всего растения, м2 • г-1);
RLR = RMF SRL
(root length ratio — общая длина корней на общую массу растения, м • г-1).
Между тем многочисленные исследования подтверждают, что LAR представляет собой важнейший детерминант роста (рис. 13.35), причем решающими переменными для значения LAR могут быть как LMF, так и SLA.
Рис. 13.35. Регулирование роста путем ассимиляции и инвестиций. Относительная интенсивность роста варьирует независимо от интенсивности ассимиляции СO2 суммой всех листьев (ULR — unit leaf rate), но коррелирует линейным образом с отношением листовой поверхности к общей биомассе (LAR — leaf area ratio). Зависимость от LAR принадлежит преимущественно SLA-компоненту (поверхность листьев на их сухую массу), а не отношению массы листьев к общей биомассе (LMF). Данные по 51 разному травянистому растению при оптимальных условиях роста сведены Fl.Poorter и A. Van der Wert
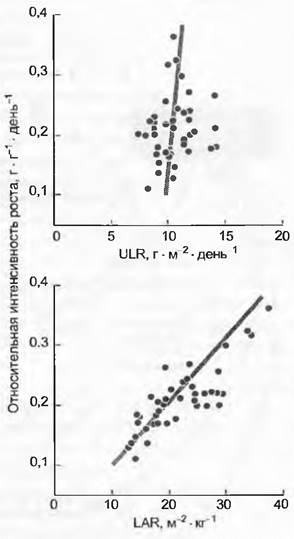
LAR — прежде всего статичная величина. Необходима еще оценка специфической продуктивности фиксации углерода листовой поверхностью. Она будет обозначена как ULR (unitleaf rate) и описывается приростом сухого веса всего растения по отношению к 1 м2 листовой поверхности в день (г • м-2 • день-1). Отсюда вытекает и относительная степень роста RGR одного растения:
RGR = ULR • LAR.
Относительное изменение общей массы М растения по отношению к исходной массе в день получается из:
RGR = 1/М • dM/dt (г • г -1 • день-1, или % • день-1).
ULR представляет собой интегральную меру для ассимиляционной продуктивности листьев по отношению к достигнутому с ее помощью нетто-приросту растения и отражает ассимиляцию углерода более реалистично, чем кратковременное измерение листового фотосинтеза. Недостаток ULR состоит в том, что она определяется только деструктивно (прирост массы за относительно короткие, например, недельные интервалы) и поэтому ее можно установить только для травянистых или во всяком случае мелких растений. Для деревьев в грубом приближении могут быть сопоставлены с LAI прирост древесины и продукция опада. Так как при росте большую роль играет эффект «сложного процента» и ход кривой роста во времени редко бывает линейным, делить годовой прирост растения на длительность вегетационного периода и считать результат этого деления за RGR некорректно. Употребляющаяся в сельском хозяйстве мера продуктивности роста растения за более длительные периоды (несколько недель, месяцы) — расчетный синоним ULR, интенсивность нетто- ассимиляции (NAR — net assimilation rate) — обозначает прирост биомассы на усредненную листовую поверхность в течение одного, как правило, более долгого периода наблюдений. Однако отношение к быстроменяющейся за этот длительный промежуток времени листовой поверхности проблематично.
RGR варьируется в зависимости от вида двумя порядками величин. У деревьев LMF и RMF более-менее условно, так как прирост благодаря включению массивной неактивной ядровой области древесины будет очень мал. По отношению к активной проводящей части древесины LMF и RMF будут иметь почти те же значения, что и у многолетних травянистых растений. Из-за ежедневной «уплаты процентов» на прирост листьев RGR может очень быстро достичь крайне высоких значений, 20 % прироста в день у молодых травянистых растений не редкость. Многие анализы между тем подтверждают, что быстро и медленно растущие растения различаются преимущественно в инвестициях биомассы относительно листовой поверхности и суммарной длины корней. Быстро растущие растения в противоположность медленно растущим имеют большие значения SLA (см. табл. 13.1 в разделе 13.6.2) и SRL. Типичные значения этих параметров приведены в табл. 13.2.
Таблица 13.2. Параметры* функционального анализа роста
Тип растительности |
LMF |
SMF |
RMF |
SLA |
SPL |
LAR |
RGR |
Травянистые растения |
0,25 |
0,45 |
0,30 |
25 |
50 |
6 |
0,15 |
Сезонно-зеленые деревья |
0,02 |
0,85 |
0,13 |
12 |
— |
0,24 |
0,02 |
Вечнозеленые хвойные деревья |
0,04 |
0,83 |
0,13 |
3 |
— |
0,12 |
0,02 |
* Грубые контрольные цифры для уже хорошо развитых, но еще не сенильных особей в природе; у всходов, ювенильных и сенильных особей численные значения могут сильно отклоняться. LMF, SMF, RMF даны в г • г-1, LAR — в м2 • кг-1 и RGR — в г • г-1 • день-1 (все по отношению к сухому весу всего растения), SLA— в м2 • кг-1, SPL — в м • г -1 в единицах веса сухой ткани (листьев или корней).
Рис. 13.36. Модель функциональной связи распределения и ассимиляции Си N с длительностью жизни листьев. Приняты во внимание только внутренние регуляторы роста растения и доступности питательных веществ без включения таких внешних факторов, как климат и влажность почвы, а также процессы развития и биотические взаимодействия (А — продукция фотосинтеза, LAR — leaf area ratio, SLA — specific leaf area)
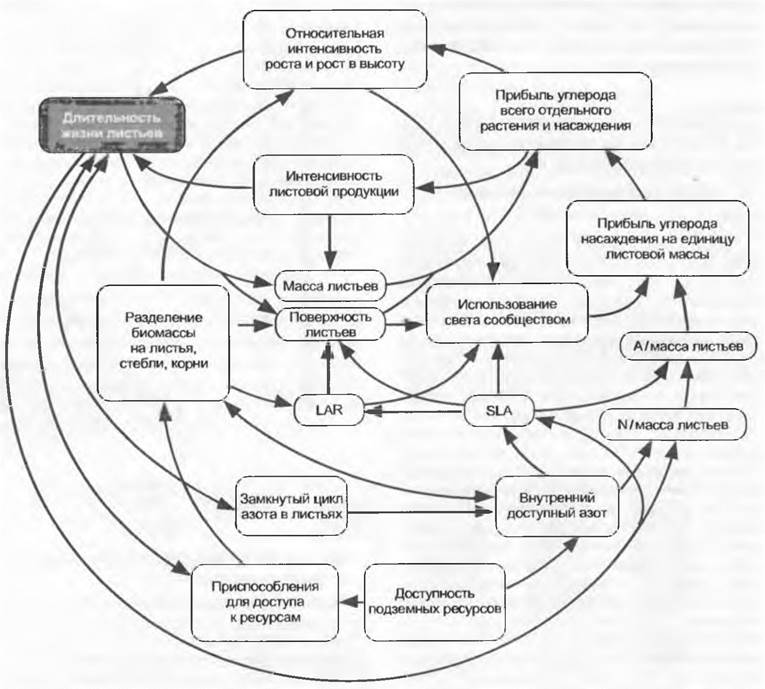
Рис. 13.36 схематически иллюстрирует согласованность важнейших детерминантов функционального анализа роста и при этом показывает (дугообразными стрелками) отношения отдельного листа к прибыли углерода в монокультуре. Эта функциональная схема была бы еще сложнее, если бы попытались включить многогранные (в значительной степени неизвестные) взаимодействия с симбионтами, фитофагами, патогенными организмами и деструентами и понять механизм процесса развития растения (например, цветения), принимая во внимание эти факторы. Комплексность еще более повышается, если и на уровне сообщества также принять в расчет надземные и подземные взаимодействия между различными видами растений и возрастными группами. Рост растительного сообщества поэтому нельзя предсказать (смоделировать) на механистической основе, исходящей из функций одного листа. Для этого отсутствуют данные по бесчисленным параметрам. Такие параметры совершенно непонятны, поскольку они представляют собой переменные, зависящие от других переменных. Часто используемые представления о механической системе из шестеренок и приводных ремней ведут к ошибкам, так как в реальном мире растения «колеса при работающем моторе, его величина и форма, а также число зубцов варьируют, приводные ремни растягиваются в направлении применяемых усилий», и кроме того, рабочая программа (развитие) быстро меняется. Каждый прогноз роста растения имеет поэтому статистическую природу и основывается на тенденциях (вероятностях), которые в такой же степени выводятся из статистических наблюдений в прошлом. В растениеводстве это давно известно. Показанная здесь комплексность проясняет, почему нельзя ожидать того, что путем генетического вмешательства в какой-нибудь частный процесс (например, в процессы, происходящие на мембранах хлоропластов) можно будет в естественных условиях получить прогнозируемые результаты роста.
13.7.4. Устойчивый изотоп 13С в экологии
Часто исходным пунктом для изменения течения науки являются новые методы. Никакое другое научное знание не имело большего влияния на исследования в функциональной (ориентированной на процессы) экологии с середины 70-х годов XX в., как то, что имеющиеся везде в природе изотопы таких важных химических элементов, как водород, азот, кислород (см. 13.5.4, 13.6.4) и углерод, встречаются в растениях или обогащенно, или редуцированно по сравнению с окружающей средой. Такие сигналы распространяются по пищевым цепям, поэтому от этого выиграла экология не только растений, но и животных. Наиболее значимый из них изотоп 13С. Он составляет 1,1% всего углерода в горных породах, атмосфере и живых организмах, 98,9% приходится на изотоп 12С. Образующийся непрерывно в настоящее время, но в крайне ничтожных количествах в верхних слоях атмосферы радиоактивный изотоп 14С, относительно быстрое разложение которого используется для определения возраста органического материала и который также применяется в качестве маркирующей субстанции (меченый атом) в аналитических исследованиях, здесь не рассматривается.
Соединения, содержащие тяжелый изотоп, имеют свойство несколько медленнее диффундировать, чем те же соединения, но с более легким вариантом того же элемента, и во многих случаях скорость процесса замедляется. Говорят, о физическом и биохимическом фракционировании (разъединении) устойчивых изотопов. В случае углерода это касается преимущественно перемещения 13СO2. Здесь фракционирование происходит таким образом, что СO2 с более тяжелым изотопом 13С в растительных тканях дискриминируется от СO2 с более легким изотопом 12С, т. е. 13С встречается в меньшем количестве по сравнению с его содержанием в воздухе (бокс 13.1).
Бокс 13.1. С помощью δ13С по следам углеродного и водного баланса
Измерительный инструмент для идентификации соотношений изотопов — масс-спектрометр, который все более становится стандартным прибором в биологии. Величина пробы лежит в пределах немногих миллиграммов. При этом нужно уловить 0,1 %о разницу в соотношении изотопов 13С/12С. Вместо абсолютных концентраций рассматривается, как правило, относительное отклонение соотношения 13С/12С в пробе от стандартного соотношения. Утвержденное международным соглашением вещество, по которому рассчитываются все прочие отношения 13С/12С, — белемнитовый известняк РееDее-формации, содержание δ13С в котором определено как 0 %с. По отношению к этому значению рассчитывается значение δ13С любого другого вещества по следующему уравнению:
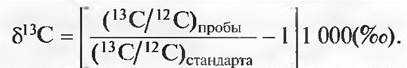
СO2 в атмосфере, из которой в растения поступает δ13С, имеет в настоящее время значение, на -8 %с отличающееся от белемнитового стандарта. Это значение становится все более негативным из-за сжигания ископаемых запасов угля. В XIX в. оно составляло менее -7 %о (реконструкция по воздуху из растаявшего полярного льда). Вместо соотношения изотопов по сравнению с белемнитовым известняком, дающим отрицательные значения δ13С, в качестве альтернативы дискриминации 13С (позитивной) приводится величина А по сравнению с воздухом:
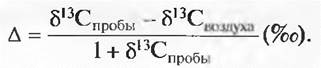
Объем дискриминации 13СO2 при процессах фотосинтеза дает представление о важных этапах восприятия СO2 (устьичная диффузия и карбоксилирование). Поскольку ассимиляты интегрируют свои «подписи» и длительно сохраняются в структурных тканях растения, значения δ13С являются зеркальным отображением условий ассимиляции в течение роста растения в настоящее время в такой же степени, как и тысячи или даже миллионы лет назад. Связь между газообменом и дискриминацией 13С была сформулирована Г. Феркьюэром и многократно подтверждена экспериментально:
![]()
соответственно δ13С пробы = δ13С воздуха + а + (b - а) рi/ра (а = 4,4 %с, фракционирование путем диффузии; b = 28 %с, фракционирование путем карбоксилирования; р1 и ра — внутреннее и внешнее парциальное давление СO2).
Поскольку ра известно, можно через ∆ вычислить значение рi, при этом из ничтожной пробы ткани получают сведения об устьичном ограничении газообмена во время образования ассимилятов. Низкое рi указывает на суженные устьица и, следовательно, на неудовлетворительное обеспечение водой.
Физическая дискриминация, т. е., в сущности, диффузия через поры устьиц, слабая и приводит к очень незначительному количеству 13С под эпидермисом — 4,4 ‰.
Этот СO2, обедненный 13С, служит для связывания фермента RubР-оксигeназы через RubisСо (рибулозо-1,5-бифосфаткарбоксилаза/оксигеназа) — процесса, при котором 13С дискриминируется существенно интенсивнее, а именно до 28 %с. Если происходит первоначально связывание РЕР-карбоксилазой (растения С4 и САМ), то это не приводит к такой дополнительной дискриминации, так как данный энзим не нарушает целостности СO2. В этом случае общая дискриминация ограничивается устьицами (4,4 ‰). Некоторые мелкие неясности возникают у растений. С3 из-за того, что выдыхаемый наружу СO2 из субстрата, уже обедненного 13С, фиксируется опять, что может играть особую роль в незначительном раскрытии устьичной щели. Таким образом, общая дискриминация будет всегда значительной (сильно негативное значение 13С), если доминирует дискриминация путем RubisСо (С3-растения при широко открытых устьицах), и незначительной (менее негативное значение δ 13С), если устьица сильно сужены и препятствуют усвоению СO2, или у С4- и САМ-растений. Поскольку исходное значение для воздуха -8 %с, теоретически 8|3С никогда не будет менее негативным, чем -12 %с (-8 + -4; С4-растения) и никогда не будет более негативным, чем -36 (-8 + -28). Фактически у хорошо снабжаемых влагой С3-растений эти значения в среднем составляют около -28,5 %с (чаще -25...32 %с), а у С4-растений — между -12 и -14 ‰. У САМ-растений они зависят от того, полностью ли идет метаболизм по типу САМ, или при влажной погоде и ассимиляция происходит по типу С3 (большей частью значения находятся в интервале -13...-20 ‰). Экологическая польза такой информации очевидна.
С помощью значений δ 13С можно по малейшим пробам мертвого растительного материала (а также гербарных образцов и фоссимизированных растений) различать растения С3- и С4-типа и, что особенно интересно, выявлять, возникли ли эти структуры в случае С3-растений в условиях водного дефицита (слабонегативное δ 13С) или при хорошем водоснабжении (сильнонегативное δ 13С). По жиру, костям или зубам животных можно узнать, выпасались ли они и когда именно на пастбищах с растениями С4-типа. Пробы почвенного гумуса могут показать, возник ли он из остатков растений С3- или С4-типа (доказательства исторических смен растительности). Этим методом удалось, например, доказать, что ископаемые моллюски с современной территории пустыни Негев тысячи лет назад питались хорошо водоснабжаемыми растениями С3-типа (влажный климат), что в течение истории Земли растения С4-типа всегда преобладали в те эпохи, когда содержание СO2 в атмосфере было низким, а также доказать, что в листьях высокогорных растений всего мира ограничение карбоксилизирования относительно незначительно, в сравнении с близкими видами растений низменных территорий (менее негативное δ13С), что отражается и на почвенном гумусе (рис. 13.37). Также в результате изотопного анализа фоссилизированных отложений бактерий получены первые свидетельства существования на Земле фотосинтезирующих организмов миллиарды лет назад. Поскольку 13С также является полностью безопасным маркирующим веществом (меченым атомом), имеющимся в нашем распоряжении, этот изотоп может служить заменой радиоактивному изотопу 14С.
Рис. 13.37. Изменение значений δ13С в растениях (листьях) и прилегающего почвенного гумуса вдоль 3 000-метрового высотного профиля в Новой Гвинее (затенен 95%-й доверительный интервал). Почва содержит изотопные знаки растений. Растения интегрируют дискриминацию за периоды от месяца до нескольких лет, почвы запасают информацию столетиями и тысячелетиями. Горные растения дифференцируют 13С в меньшей степени, чем растения в долинах
13.7.5. Биомасса, продуктивность, глобальный круговорот углерода
13.7.5.1. Запасы биомассы
Большая часть биологически связанного углерода находится на суше, причем около 1/5 его сосредоточено в растениях и 4/5 — в почвенном гумусе (см. рис. 13.43). Общая биомассаприблизительно на 85 % заключена в деревьях (рис. 13.38).
Рис. 13.38. Распределение заключающихся в биомассе запасов углерода на Земле по крупным биомам. Сравнительные данные относятся к общим запасам углерода, оцениваемым в 559 млрд т (сухая биомасса содержит 46 — 50% С) Запасы биомассы и углерода на единицу поверхности даны в калькуляционных средних единицах. Если рассматривать только ненарушенную зрелую растительность, запасы могут быть и существенно выше

В соответствии с определениями, данными в разделе 12.5.1.4, биомасса не включает отмершие части растений. Однако точное разделение био- и некромассы при такого рода общей статистике на практике невозможно. То, что здесь, следуя литературным источникам, обозначается как биомасса, в действительности является фитомассой (т. е. включает и некромассу). Если задуматься, что эти стволы деревьев представляют собой эволюционный результат конкуренции за свет, а частично и за окружающее пространство («бегство» от фитофагов и пожаров), то это означает, переводя на повседневный язык, что преобладающая часть надземной биомассы удивительным образом состоит из «расходов на рекламу».
Почти половина всех лесных насаждений Земли (около 42 % общей биомассы) находятся в тропиках и субтропиках. Сумма средних запасов биомассы в аграрных культурах достигает примерно 1,6 %, только 0,2 % общей биомассы находится в океанах. В лесах большая часть биомассы сосредоточена в надземной сфере (около 80 %), в злаковых сообществах — под землей (более 60 %, крайние значения — до 90%); 60 — 80% обшей биомассы корней сосредоточено обычно в верхнем слое почвенного профиля, на глубине до 30 см, но небольшая часть корней проникает, как правило, в почву на глубину нескольких метров, за исключением субполярной растительности и переувлажненных местообитаниях (табл. 13.3).
Таблица 13.3. Масса корней в крупных биомах, общая и на единицу площади, а также средняя максимальная и абсолютно максимальная глубина проникновения корней
Биом |
Площадь на Земле, 106 км2 |
Массакорней, Кг ∙ м-2 |
Доля корней до 30 см глубины |
Максимальная глубина проникновения корней*, м |
||
Гт |
% |
средняя |
абсолютная |
|||
Влажные тропические леса |
17 |
4,9 |
83 |
69 |
7,3 |
18 |
Сезонные (суб)тропические леса |
7,5 |
41 |
31 |
70 |
3,7 |
4,7 |
Вечнозеленые умеренные леса |
5 |
4,4 |
22 |
52 |
3,9 |
7,5 |
Листопадные умеренные леса |
7 |
4,2 |
29 |
65 |
2,9 |
4,4 |
Бореальные хвойные леса |
12 |
2,9 |
35 |
83 |
2,0 |
3,3 |
Открытые леса и буши |
8,5 |
4.8 |
41 |
67 |
5,2 |
40 |
Тропические злаковые сообщества |
15 |
1,4 |
21 |
57 |
15,0 |
68 |
Злаковые сообщества умеренного пояса (прерии, степи и др.) |
9 |
1,4 |
14 |
83 |
2,6 |
6,3 |
Тундровая и альпийская растительность |
8 |
1,2 |
10 |
93 |
0,5 |
0,9 |
Жаркие пустыни |
18 |
0,8 |
6,6 |
53 |
9,5 |
53 |
Культурные земли |
14 |
0,2 |
2,1 |
70 |
2,1 |
3,7 |
По подсчетам Р. Дженсона и Д. Кэнделла, общая масса корней земли составляет около 235 Гт сухого вещества, или около 140 Гт углерода. Поскольку в этом источнике из-за специфического подбора относящихся к корням литературных данных включена биомасса корней, проникающих в более глубокие слои почвы, что не отражается в классических таблицах общей биомассы Земли, где обычно не учитывается масса корней ниже 30 см от поверхности, значения оказываются по сравнению с последующими более высокими (85 Гт сухой массы, или около 40 Гт углерода, причем глобальный запас углерода соответственно должен повыситься с 560 до 600 Гт) (см. рис. 13.38 и 13.39). Разница в процентных долях площадей (рис. 13.38) связана с различной ординацией формаций.
* В целом средняя максимальная длина корней составляет для деревьев — 7 м, кустарников — 5, травянистых растений (включая злаки) — 2 — 6, возделываемых растений — 2 м. Без дополнительных данных можно в первом приближении исходить из того, что эти жизненные формы верхушками наиболее длинных корней достигают названных глубин (во влажных прохладных районах глубина может быть меньшей, в жарких и сухих— большей)
Запасы мертвого растительного вещества могут в некоторых злаковых сообществах достигать 50 — 90 % от всей растительной массы (отмершие листья и основания листьев). В лесах может быть аналогичная ситуация, если физиологически более неактивную ядровую древесину рассматривать как «мертвую» (в противоположность активной заболони), что статистически, впрочем, доказать нельзя. Мертвая масса растений в почве, так называемая почвенная подстилка, в злаковых сообществах большей частью невелика, но в лесах она составляет 5 — 10 га-1 (нижнее значение для лиственных лесов, верхнее для хвойных), причем годичная продукция подстилки колеблется от 4 до 5 т • га-1, что приблизительно равно годичному приросту древесины.
Как только растительность полностью покрывает почву, биомасса варьирует без существенного влияния на LAI (см. 12.5.1.5). У сенокосного луга и букового леса хорошего бонитета близкие значения LAI — около 6. Соответственно, количество хлорофилла в сомкнутой растительности на единицу земной поверхности во всем мире имеет близкие значения (2 — 3 г • м-2).
Величина запаса биомассы ничего не говорит об обменных процессах. Хотя в океане находится только ничтожно малая часть биомассы Земли (в основном планктон), эти организмы в сумме поставляют почти такое же количество углерода в год, как и наземная растительность. Данное сравнение показывает, что разница между «фондом» и потоками (реакциями обмена) существенна для понимания углеродного режима, прежде всего для проблем СO2, которые дискутируются далее (см. 13.7.6).
13.7.5.2. Продукция биомассы
Когда растения растут, увеличивая при этом за время роста количество биомассы на единицу площади, говорят о продукции биомассы, или (выражая интенсивность за единицу времени) о продуктивности. Поскольку продукция биомассы растений стоит в основании пищевой цепи, ее называют первичной продукцией. Различают брутто-первичную продукцию ВРР, т. е. количество биомассы, которое в совокупности синтезируется на единицу площади, и нетто-первичную продукцию NPP, которая результируется после вычета текущих потерь на дыхание R в экосистеме:
![]()
Эти продукционные термины, известные из хозяйственной жизни и употребляемые в биологии вообще в большой степени, имеют теоретическую сущность и на практике едва ли употребимы поскольку величины потерь, в особенности тех, которые происходят в подземном слое и через дыхание, как правило, неизвестны. Оценки NPP, таким образом, могут грешить большими ошибками.
Как можно принять из приблизительных контрольных цифр, примерно половина усвоенного растением углерода еще при его жизни отдается как СO2, выделяемое при дыхании. Другая часть, как правило, возвращается в атмосферу при микробном разложении. В глобальном сопоставлении данных для лесов Дж. Райх и К. Надельхоффер продемонстрировали, что ежегодная продукция наземного опада (70 — 500 г/м -2 • а-1) соответствует высвобождению СO2 из почвы («почвенному дыханию»), из чего можно заключить, что надземная и подземная продукции определяются одними и теми же факторами.
NPP — величина, употребляемая чаще всего. еще никогда не была определена даже предположительно по вышеприведенной формуле. Для этого следовало бы знать за весь период наблюдения интегральное количество связанного углерода и все потери на дыхание. Этого избегают и обыкновенно рассматривают изменение запасов биомассы (ДВ) между двумя временными точками (г • м-2 • а-1). Проблема состоит в том, что в течение интервала наблюдений очень много продуцируемой биомассы снова теряется. Отмершие части побегов (V0) еще можно непрерывно собирать и добавлять в заключительных расчетах (естественно без учета потерь летучих веществ типа изопрена — Ех0); труднее реконструировать ту биомассу, которую «консумировали» фитофаги и патогенные организмы (К0), для подземных же органов совершенно невозможно определить потери и «консум» биомассы(V0, К0). В злаковых сообществах больше двух третей биомассы приходится на подземные органы, и большая часть тонких корней недолговечна. Есть оценки, согласно которым еще 5 — 10% ассимилятов попадет в микоризообразующие грибы и там на короткое время задерживаются, что тоже не определимо. Количество корневых выделений в ризосферу (сахара, аминокислоты) в значитель ной степени неизвестно, как и в большинстве случаев потери растворимых органических соединений (dissolved organicmatter — DOM), уходящих в просачивающуюся воду (Еxy). Если NPP, определенная на основе изменений запасов биомассы ДВ, может оказаться неправильной на 100%, то уравнение
![]()
(V — отмершая, потерянная биомасса; К — консумированная биомасса; Ех — экспорт биомассы как надземный, так и подземный) практически нерешаемо. Только в том случае, если все эти величины потерь во всех экосистемах равновелики, данные по NPP, полученные на основании урожая биомассы (∆В), были бы сравнимы, но это совершенно невероятно. Дополнительная проблема— распределение биомассы в течение периода наблюдений. Поскольку новые надземные структуры, строящиеся из резервов в лежащих глубоко в почве органов, в действительности в течение периода наблюдений не были продуцированы (англ. stored growth), биомасса лишь (с метаболическими затратами) переотлагалась снизу-вверх.
Из-за неизвестных величин потерь более корректно вместо NPP говорить о нетто-приросте фитомассы или, как принято в сельском хозяйстве, об урожае (от англ. harvestedyield). Из-за этой неточности значения NPP цитируются как NPP. Почти 46 — 50% сухого вещества приходится на углеродные единицы. Средняя энергетическая емкость (теплотворная способность) биомассы составляет для наземных растений 18,1 кДж, для океанического планктона — 19,3 — 20,6 кДж.
Если прирост биомассы в крупных биомах Земли оценивают, как практикующееся приближение к настоящей «продуктивности», результат зависит от выбранного масштаба времени. Если включать непродуктивное время года в областях, близких к полюсу (зимний покой), т. е. оценивать прирост биомассы за год независимо от фактического времени продуцирования, то продуктивность по направлению к полюсу будет падать. Если сравнивать только периоды активного роста, то продуктивность везде на Земле примерно одинаковая пока достаточно воды. Этот поразительный, часто не замечаемый результат показывает, как исключительно физиологические приспособления повсеместно уравновешивают климатические различия. Широтные различия в продуктивности (не говоря о региональных и локальных различиях в условиях роста) почти исключительно являются результатом различной длительности сезона роста; влияние климата в течение этого сезона очень невелико. Если вместо года взять среднемесячные значения периода вегетации, то высокогорные растения умеренной зоны (Альпы) продуцируют биомассы не меньше, чем в среднем влажный тропический лес, которому в этом не уступают и центрально-европейские буковые леса (рис. 13.39). При особо благоприятных условиях роста, как и в условиях интенсивного сельского хозяйства, аккумуляции биомассы в год в экстремальных случаях достигают свыше 5 кг • м-2.
Рис. 13.39. «Продукция» биомассы в различных экосистемах. Сравниваются показатели за год (справа жирным шрифтом), включая периоды ростового покоя во внетропических областях, или в пересчете на средний месяц вегетационного периода (прямоугольники слева), что свидетельствует об очень разных результатах (данные только для гумидных областей). Это наглядно показывает, что глобальная разница в годовой аккумуляции биомассы едва ли находится под влиянием температурных условий во время сезона роста. Средние данные продукции для лесов и для злаковых сообществ не отличаются (данные в скобках показывают картину большой региональной и локальной вариабельности)
Трудность при экстраполяции запасов и прироста биомассы состоит в том, что очень трудоемкие анализы проводятся только в немногих пунктах Земли. Понятно, что для таких работ часто выискиваются «красивые» и зрелые насаждения. Поэтому глобальная биомасса, очевидно, значительно ниже ее оценки периода Международной биологической программы (1968 — 1974). Глобальный запас земной биомассы был определен тогда в 840 Гт углерода. Новые оценки, проводившиеся на площадях с «не идеальном» растительностью и оголенными поверхностями, лежат в пределах 560 — 600 Гт углерода (если не обращать внимания на быстрое исчезновение лесов в тропиках, оценивающееся в 1— 2 Гт углерода в год, чему противостоит нетто-прирост в умеренной зоне; см. 13.7.6).
Таблица 13.4. Биомассы центрально-европейского смешанного дубово-грабового леса
Организмы |
Масса сухого вещества, т • га-1 |
Зеленые растения |
275 |
Листья древесных растений |
4 |
Ветви |
30 |
Стволы |
240 |
Травы |
1 |
Животные (наземные) |
> 0,0004 (3 — 5 кг • га-1) |
Птицы |
0,0007 |
Крупные млекопитающие |
0,0006 |
Мелкие млекопитающие |
0,0025 |
Насекомые |
9 |
Почвенные организмы |
ок. 1 |
Дождевые черви |
0,5 |
Другие почвенные животные |
0,3 |
Почвенная флора |
0,3 |
Первичные продуценты своей живой и отмершей массой образуют основу для дальнейшей переработки вещества консументами и редуцентами, т. е. для вторичной продукции.Согласно процессам, в пищевых цепях растительная масса в сотни раз больше, чем масса вторичных продуцентов. Среди консументов наибольшая масса свойственна фитофагам, в то время как хищники первого и второго порядка и соответственно паразиты первого и второго порядка с постепенно уменьшающейся зоомассой составляют вершины пищевых пирамид с различными трофическими ступенями. Табл. 13.4 и рис. 13.40 иллюстрируют в качестве примера распределение биомасс и величин продукции центрально-европейского дубово-грабового смешанного леса. При этом видно, что пищевая пирамида соответствует пирамиде продукции, поскольку первичная нетто-продукция достигает здесь, как и в других экосистемах, на порядок более высокие (в десятки и сотни раз) значения, чем значения вторичной продукции.
Рис. 13.40. Годовое излучение Солнца, а также первичная и вторичная продукция в центрально-европейском смешанном дубово-грабовом лесу (ср. табл. 13.4)
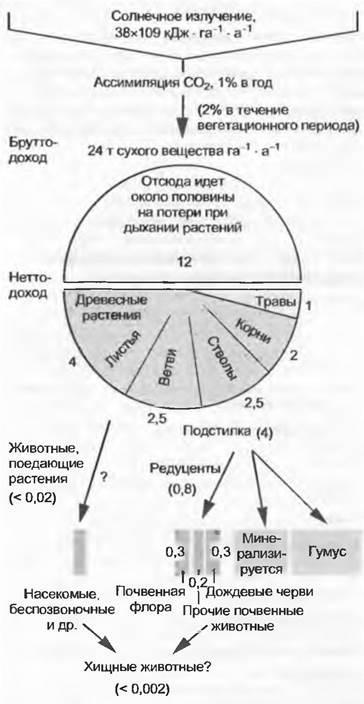
Рис. 13.40 показывает также, что консументы принимают лишь незначительное участие во вторичной продукции, потому что из первичной продукции растений в дубово-грабовом лесу непосредственно потребляется фитофагами только около 2 % (в других наземных биоценозах едва ли более 15%, в среднем около 7%). При этом ежегодно почти 25% продукции аккумулируется как мертвая органическая субстанция в твердом состоянии (детрит: листовой опад, гумус и др.) или в почвенных растворах (например, гумолигниновые кислоты). Такому количественно важному компартименту этой экосистемы соответствуют большая значимость и продуктивность сапротрофных и минерализующих редуцентов (см. 9.1.1, бокс 11.4); их участие во вторичной продукции составляет 95 %.
Несмотря на ненадежность выше приведенной базы данных, оценки «NPP» в глобальном сравнении очень иллюстративны (рис. 13.41). Хотя океаны заключают в себе только 0,2 % глобального запаса биомассы, они по своей площади, составляющей 70 % всей поверхности, имеют почти такие же высокие значения «NPP», как и вся суша. Продуктивность тем не менее сосредоточена в богатых питательными веществами областях и регионах, т. е. в прибрежных, с поднимающимися со дна холодными глубинными водами (upwelling regions). Удаленные от побережья тропические и субтропические регионы мирового океана — продуктивные пустыни (белые поверхности на карте). Примечательно, что максимальная естественная продуктивность в море (специфические прибрежные зоны) и на суше может быть очень сходной, а именно 2 000 — 3 000 г • м-2 • а-1 (точечные пиковые значения «NPP» — до 6000 г • м-2 в переходных зонах от суши к морю, почти как в тропических и субтропических болотах). На суше среднегодовая «NPP» сомкнутой растительности варьирует по географической широте и водообеспеченности от 200 (субполярные области) до 2 500 г • м2 (влажные тропические леса, вся биомасса дана в абсолютно сухом весе). Продуктивность лесов умеренной зоны около 1 000 — 1 500 г • м-2 (см. рис. 13.39). Почти 25 % земной поверхности (около 33 млн км2) показывают годичную «NPP» — не более 500 г • м-2. Для культурных растений верхняя граница может приближаться к 7 000 г • м-2 (орошаемые интенсивные культуры сахарного тростника). Культуры водорослей (например, Scenedesmus) могут в лабораторных условиях давать 10 000 г • м-2 биомассы, но практическое применение их затруднено.
Рис. 13.41. Первичная нетто-продукция биосферы. Данные приводятся в граммах сухого вещества на 1 м2 в год для суши и океанов
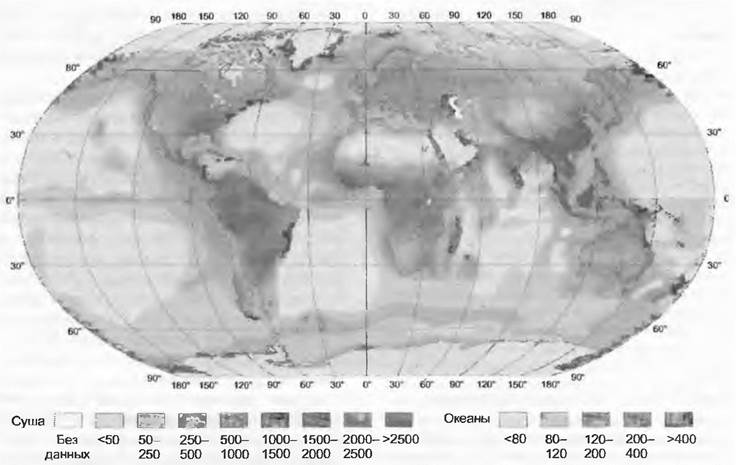
Глобальная «NPP» составляет по новейшим подсчетам около 210 — 250 млрд т биомассы или 100 — 120 млрд т углерода (46 — 50% биомассы), из них по 50 — 60 млрд т углерода в океане и на суше (1 млрд т = I Гт = IPg = 1015 г). На суше приблизительно половина «NPP» падает на тропики. Средняя продолжительность пребывания углерода на суше составляет 22 года, в океане — близко к неделе (планктон <5 мм; значительная величина отношения поверхности к объему).
13.7.5.3. Нетто-продукция экосистемы и биосферы
Нетто-продукция экосистемы NEP (nelecosystem production) — «продукционный параметр», который гораздо более определен, чем NPP и измеряется в большем приближении к реальности. NEP представляет собой баланс нетго-углерода в экосистеме, т. е. разницу в поглощении и выделении углерода, без рассмотрения того, где и как углерод периодически связывается в системе.
NEP более рационально получать за длительные интервалы времени (по меньшей мере за год) и на больших площадях (> 1 га). Базовые данные — измерение потоков СO2 (внос вынос) метеорологическими методами (измерительные вышки над гомогенным, плоским участком ландшафта). Так называемый «eddy covariance» (вихревой ковариантный) метод измеряет с помощью трехмерного ультразвукового ветроизмерительного аппарата вертикальные брутто-потоки «квантов» воздуха и соединяет их тотчас же с открытым (open path) инфракрасным газоанализатором, измеряющим концентрации СO2с очень высоким временным разрешением. Трудности этого метода измерения NEP заключаются в том, что он охватывает очень маленькую разницу в нетто очень больших потоков. Чтобы доказать, что NEP не равна нулю, требуется очень высокая точность измерений. Выбор «хороших» гомогенных насаждений ведет, как правило, к переоценке неттопотоков на уровне всего ландшафта. Экспорт углерода в других формах, чем СO2, обычным способом измерения NEPне охватывается.
Для функционирования экосистемы большое значение имеет не только поток вещества и энергии между самой системой и ее окружением, но и размещение и интенсивность потоков внутри системы. Годовой поток энергии субтропического родникового озера (рис. 13.42) несет очень небольшую фитомассу (но многочисленные генерации планктонных водорослей) и иллюстрирует, как распределяется энергия первичной продукции вдоль пищевых цепей экосистемы.
Рис. 13.42. Поток энергии через естественную экосистему планктона (субтропическое родниковое озеро Сильвер Спрингс, Флорида)
Значения входа и выхода даются в кДж • м-2 • а-1. Компартименты слева направо; первичные продуценты, консументы (растительноядные — PFLF и хищники первого и второго порядков — FF.) и редуценты (ZS.)
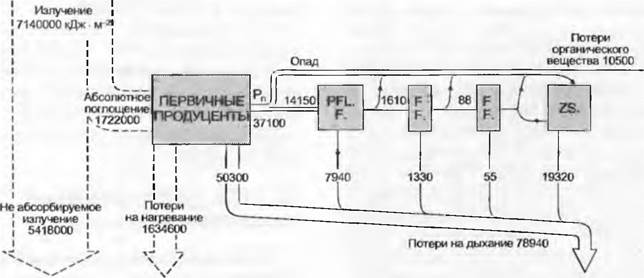
В наземных биоценозах, особенно в длительно существующих лесах, круговорот вещества и энергии по отношению к фитомассе происходит существенно медленнее. Хотя в этом родниковом озере абсорбируется почти четверть излучаемой солнечной энергии, относительно густой фитопланктон может в конце концов, несмотря на отсутствие зимнего покоя, использовать для брутто-продукции только 1,2 % общего излучения. После относительно высоких потерь на дыхание (70%) для консументов и редуцентов остается только 20 %.
Чем более зрелой является экосистема, тем больше ее NEP приближается к нулю. Большое значение при таком балансе углерода имеют запасы углерода в почве (в большинстве случаев 10 — 20 кг • м-2). В гумидных тропиках они составляют часто не более 10 — 20 %, в бореальных хвойных лесах — 60 — 70 %, в тундре — более 90 % всего углерода экосистемы. Когда в быстрорастущих лесных посадках почвенный гумус вначале быстро разлагается (дренаж, удобрение, известкование), NEP может быть негативной, несмотря на очень позитивную «NPP». В молодых экосистемах NEP большей частью положительна, в зрелых приближается к нулю, в старых деградирующих негативна. Только охват всех этих стадий развития, существующих в ландшафте (лесной области), дает информацию о том, имеет ли место закрепление в экосистеме нетто-углерода или нет.
Лес в стадии восстановления, а также спелый лес до рубки всегда имеют позитивные значения NЕР. Дальнейшая судьба деревьев определяет долгосрочное развитие NЕР. Если в обогащенной грубым и мягким гумусом экосистеме после распада мертвой древесины вырастает следующее поколение деревьев, NЕР в течение сотен лет может оставаться позитивной. Если полезная древесина перерабатывается и в конце концов рециклируется (бумага, утиль, сжигание, разложение), то калькулируемая NЕР снова приближается к нулю. Деревянные постройки, например, представляют собой промежуточный фонд углерода.
Итак, объективное суждение о балансе углерода в ландшафте разрывает рамки экосистемы, поэтому для очень больших масштабов и длительных промежутков времени NЕР заменяют на NBР — нетто-продукцию биома. NBР охватывает процессы на ландшафтном уровне, такие, как пожары, ветровалы, массовые нападения насекомых, принимает во внимание все стадии развития растительности, в том числе «окна» внутри нее, и включает также последствия любой деятельности человека. В большей части мира NВР в настоящее время негативна, это значит, что биомы теряют нетто-углерод (корчевка лесов, интенсивная обработка почвы, экспансия густозаселенных и индустриальных территорий), хотя местами еще существуют подвергающиеся экспансии экосистемы с позитивной NВР.
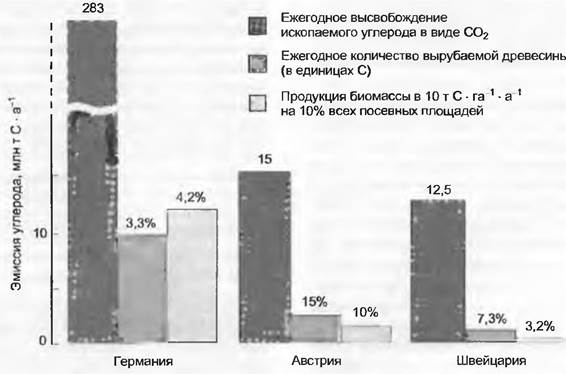
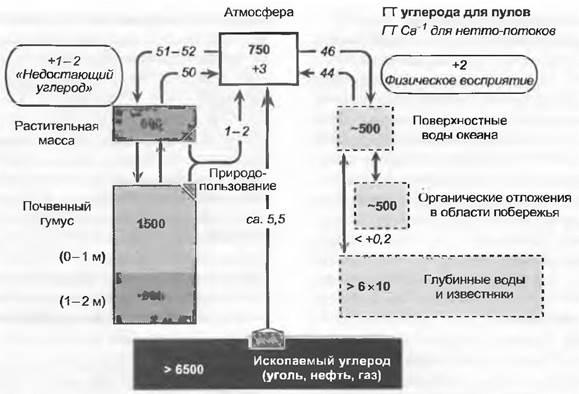
Следующий шаг ведет к биосфере в целом. Ее углеродный баланс приблизительно уравновешен, так как экосистемы Земли в среднем связывают столько же углерода, сколько и отдают. Продолжающееся непрерывно сведение лесов в тропиках поставляет ежегодно в атмосферу 1 — 2 Гт углерода, еще неизученные биотические поглощения связывают опять же 1 — 2 Гт С, причем предполагается, что это связано с удобряющим эффектом повышенного содержания СО2 в атмосфере и экстенсификацией природопользования в различных частях Северной Америки и Европы (вторичные и слабо-используемые леса). Сейчас увеличение содержания углерода в воздухе связано с тем, что человек высвобождает в атмосферу ископаемые запасы углерода в количестве 5 — 6 Гт, часть их растворяется в морской воде, так что ежегодно в атмосфере остается «только» почти 3 Гт дополнительного углерода. Это повышает атмосферный пул углерода (в СО2) с современного значения 750 Гт в год на 0,4%, или почти 1 ppm (современное значение 370 ppm). Продолжение этой тенденции приведет к тому, что в конце XXI столетия общий уровень СО2 по отношению к доиндустриальной эпохе удвоится (см. 13.7.6). Включенные в глобальный круговорот углерода фонды и потоки, в том числе современные антропогенные потоки, иллюстрирует рис. 13.43.
Рис. 13.43. Глобальный круговорот углерода в среде, находящейся под воздействием человека (заштрихованы антропогенные источники С). Размеры «клеток» отражают величину запасов углерода. Только около 40 % высвобождаемого ископаемого углерода остается в настоящее время в атмосфере, остаток растворяется в океане и закрепляется в наземных экосистемах (1 — 2 ГТС «missing carbon» — «недостающий углерод»). Эта величина примерно соответствует ежегодному высвобождению углерода из-за сведения лесов. Большие углеродное пулы в глубоких слоях океана и в известняковых породах играют роль для концентрации СО2 в атмосфере только при рассмотрении очень больших периодов времени (глубинные слои океана>200 лет, существенные взаимодействия с карбонатной геохимией >1 000 лет)
13.7.6. Биологические аспекты «проблемы СО2
В связи с тем, что ископаемые источники углерода, которые большей частью начали возникать более чем 100 млн лет назад и формировались в течение многих миллионов лет, приблизительно за 200 лет (грубо говоря, с 1910 по 2100 г., когда могут быть исчерпаны легкодоступные запасы) в виде СО2 были выкачаны человеком в атмосферу, для растительности возникает совершенно новая ситуация. Биосфера получает новую «диету», в геологических масштабах как бы за одну ночь. Поскольку СО2 — материальный базис фотосинтеза, от которого (если не считать несколько хемоавтотрофных видов бактерий) зависит вся жизнь на Земле, проблема СО2 для экологической ботаники стала центральной. Возможность того, что обогащение атмосферы СО2 также связано с климатом (так называемый парниковый эффект) и соответственно могло бы непрямым образом влиять на растения, здесь не обсуждается, речь идет только о непосредственном воздействии на растения и экосистемы.
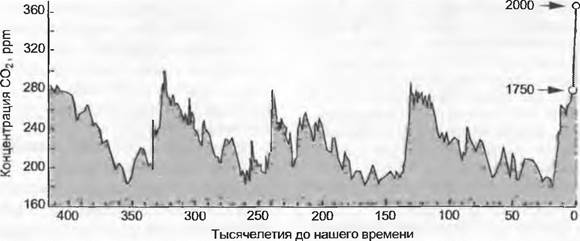
Считается, что уровень СО2 в атмосфере к началу каменноугольного периода был на уровне 2 000 — 3 000 ppm, но потом очень быстро опустился до 300 — 500 ppm, что близко к современному уровню, а также что наиболее низкие значения были в пермском периоде. Достоверно установлено, что в мелу уровень СО2 сначала был очень низким (около 300 ppm), что и объясняет первое массовое распространение растений С4-типа. Механизм концентрации СО2 растениями типа С4 осуществляется только при очень низком уровне СО2, это дает им преимущество перед растениями С3-типа. С помощью ледовых кернов из Антарктиды документирован состав атмосферы за время последних 0,4 млн лет (анализ вмороженных в лед пузырьков воздуха). Как показывает рис 13.44, концентрация СО2 колебалась за это время в пределах 180 — 290 ppm. Максимумы соответствуют теплым периодам, минимумы — пикам оледенений. С 1800 г., т. е. с начала отопления углем, кривая превысила этот диапазон и примерно к 1900 г. поднялась так быстро, что сейчас СО2 в воздухе уже на 30 % больше, чем, когда было посажено ранее большинство деревьев в парках. Почти все сейчас существующие виды растений появились в свободных ото льда областях в тот период, когда уровень СО2 был равен 180 ppm (иначе бы они просто вымерли; в последний раз концентрация СО2 была столь низкой приблизительно 20 000 лет назад). Около 1990 г. они пережили удвоение этого значения (в 2000 г. — 370 ppm). Если не произойдет глобального коллапса мирового хозяйства, концентрация СО2 за последующие 100 лет еще удвоится.
Рис. 13.44. Концентрация СО2 в атмосфере за последние 0,4 млн лет на основе анализа воздушных пузырьков в антарктическом ледниковом щите (бурение на станции «Восток»)
Исходное положение всех рассуждений о биологических последствиях повышения СО2 лежит в кривой зависимости нетто-фотосинтеза от концентраций СО2 (рис. 13.45, см. 6.5.11.2). Она показывает, что С3-растения могут существовать при еще больших концентрациях, чем сейчас, и больше фотосинтезировать. Разумеется, такие кривые являются как бы «снимками данного момента», которые только доказывают, что при обеспечении RubisCo в данный момент и без ограничения отложения ассимилятов процесс карбоксилирования еще не насыщен СО2. Как было изложено в разделах 13.7.2 и 13.7.3, ускорение роста зависит еще от многих других факторов, и только в тех случаях, когда не существует ограничения отложения, концентрация СО2 может быть причислена к стимуляторам роста.
Прирост урожая в тепличном садоводстве равняется примерно 30 % в сезон, если растениям предоставляется уровень СО2 600 ppm или более, что уже перед Второй мировой войной использовалось в немецких и голландских теплицах. Лучше всего удобренные и орошенные пшеничные поля в Аризоне давали на 14 % больше зерна, когда культивировались при уровне СО2 около 600 ppm. Такие цифры должны хорошо смотреться на фоне того, что урожаи пшеницы благодаря новым сортам и оптимальному уходу в последние 100 лет уже увеличились на 300 — 500%.
Когда все другие ресурсы, кроме СО2, относятся к ресурсам, лимитирующим рост, что в естественных условиях происходит почти всегда, остаются три возможности для долгосрочного реагирования:
✵ редукция емкости фотосинтеза (меньше RubisCo и тем самым меньше азота на листовую поверхность, или меньше листьев, т. е. более низкая LAR, см. 13.7.3);
✵ более высокий экспорт углерода (например, более быстрый обмен тонких корней, экссудация корней, экспорт в микоризу, выделение изопренов);
✵ усиление роста с помощью разбавления питательных веществ, особенно N, т. е. продуцирование биомассы с более высоким отношением C/N.
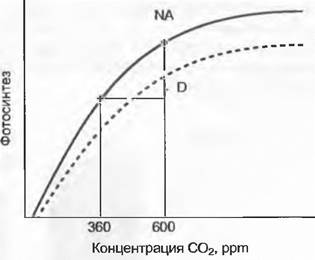
Все три пути идут, как правило, параллельно. Обычно происходит сокращение емкости фотосинтеза, но очень редко полностью, так что при повышенном содержании СО2 на единицу листовой поверхности почти всегда связывается больше углерода (рис. 13.45).
Рис. 13.45. Регулирование зависимости фотосинтеза от концентрации С02 при длительном воздействии повышающегося содержания СО2 (down regulation — пунктирная кривая) зависит от условий роста (активность поглощения ассимилятов) и возраста растений. Точка NA отмечает увеличение нетто фотосинтеза при повышении концентрации СО2 до 600 ppmбез подобного регулирования; D — сохранившаяся нетто-прибыль после регулирования
Многочисленные эксперименты показывают, что экспорт углерода при повышенном СО2 усиливается. Наблюдалось, что повышенное содержание растворимых углеводов в ризосфере ведет к усилению связывания азота почвенными микроорганизмами, следствием чего могут быть симптомы азотного голодания. Увеличение соотношения C/Nнаблюдалось почти всегда и прежде всего в листьях. В большинстве случаев общее содержание структурно не связанных углеводов повышается (например, крахмал, сахар; англ. nostructural carbohydrates — NSC). Со снижением содержания протеина (N) и увеличением содержания NSC (С) изменяется и качество питания. Для травоядных животных доказано, что это отрицательно влияет на их рост и размножение.
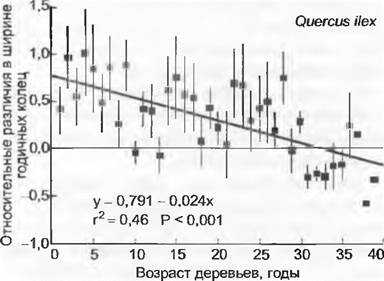
Прирост биомассы в естественной растительности (почти все данные по злаковым сообществам) обычно очень незначителен (менее 15 % при смоделированном удвоении концентрации СО2) или даже равен нулю. Для естественных лесов (85 % глобального запаса биомассы) данных не имеется, но реакция каменных дубов, растущих на границе с естественным источником СО2 в Тоскане, дает первые доказательства возможной реакции: стимуляции роста на самой ранней ювенильной фазе развития при удовлетворительном предоставлении света и питательных веществ. С увеличением возраста реагирование становится все меньшим до того момента, когда эффект сводится к нулю (рис. 13.46) Это могло бы означать некоторое убыстрение жизненного цикла. Более высокий запас биомассы на единицу земной поверхности с этим неизбежно не связан. Немалое число СО2-экспериментов со всходами деревьев дало доказательства этого только на очень ранних стадиях. Большинство видов реагирует на СО2 положительно, но очень видоспецифично, причем различия зависят от синхронного снабжения светом и питательными веществами, поэтому однозначные выводы невозможны.
Рис. 13.46. Воздействие повышенного содержания СО2 на длительный рост деревьев в окрестностях геологического источника СО2 в Тоскане (Раполано). На примере анализа годичных колец каменного дуба (Quercus ilex) можно увидеть, что деревья на ранних фазах развития близ источника по сравнению с более удаленными контрольными деревьями росли явно быстрее (рис. показывает разницу в приросте по отношению к деревьям в стороне от источника). Примерно в 30-летнем возрасте сигнал исчезает (индекс «опыт»/ контроль нулевой; сплошная линия). Полосы дисперсии показывают среднюю вариабельность в каждом случае для 10 деревьев. С помощью углеродного (14С) анализа древесины годичных колец установлено, что подвергавшиеся воздействию источника деревья фактически всегда испытывали вдвое более высокую концентрацию СО2, чем контрольные (геологический СО2 свободен от 14С, путем смешивания его с нормальным воздухом может быть реконструирована средняя концентрация СО2, которую испытывали деревья)
При всех реакциях растений на СО2 было установлено, что различные виды откликаются очень по-разному. В отдельных случаях даже наблюдалось, что различные генотипы одного вида реагируют не одинаково. Это означает, что проблема СО2 в любом случае может рассматриваться как проблема биоразнообразия особого рода. Снабжение СО2 влияет на конкурентные отношения между видами. Можно согласиться с тем, что этот фактор влияет на структуру видового состава в мировом масштабе.
Кроме прямого воздействия СО2 на фотосинтез и последующее употребление ассимилятов имеется еще и непрямое влияние СО2 через водный режим. Устьица часто реагируют на повышенную концентрацию СО2 сужением устьичной щели (см. 8.3.2.5). Реакция зависит от влажности воздуха и почвы, не у всех видов она одинаково сильная, а у деревьев более выражена на стадии проростков, чем у взрослых экземпляров. В злаковых сообществах эта реакция устьиц вызывает более медленное снижение влажности почвы в периоды, бедные осадками, что является причиной увеличения аккумуляции биомассы. Большая часть тех явлений, которые происходят в природной злаковой растительности, могут сводиться к этому непрямому эффекту. В сухие годы данный эффект соответственно проявляется сильнее, чем вс влажные (рис. 13.47). Предполагается, что эффект замедления снижения влажности в насыщенном СО2 пространстве станет реальностью, если условие испарения по сравнению с современными не изменяется.
Антропогенные выбросы СО2 в настоящее время составляют почти 6 Гт С • а-1, и будут увеличиваться далее, так что растительность в течение следующих 100 или более лет не сможет связать поступающий излишек. Гораздо вероятнее, что наземное использование дополнительного углероде переместится в атмосферу. Уже сейчас, еще на крутой ветви кривой функции зависимости фотосинтеза от СО2 при превышении СО2 на 30 % по сравнению с доиндустриальным временем максимальное нетто-связывание составляет только 2 Гт (даже если расчетное поглощение наземного углерода (см. рис. 13.43) приписать исключительно удобряющему эффекту СО2, что, вероятно, переоценивается) — это удивительно малая часть наземной первичной нетто-продукции, составляющей около 60 Гт С • а-1. Вследствие стехиометрического связывания Р и N и других ограничений относительный эффект увеличения концентрации СО2 значительно уменьшается.
Рис. 13.47. Действие повышенного содержания СО2 на участке высокозлаковой прерии. Только в сухие годы повышение концентрации СО2 стимулирует накопление биомассы, это результат воздействия СО2 на устьица, транспирацию и тем самым на влажность почвы (600 вместо 360 ppm в больших «open top» («открытых сверху») камерах).
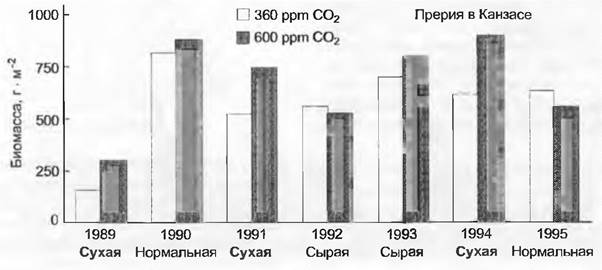
Био-менеджмент усиления связывания углерода растениями рационален и в экологическом отношении полезен, но при этом не следует питать иллюзий, что абсолютные значения его взаимосвязаны с эмиссией С. Только существенное расширение лесных площадей могло бы долговременно способствовать связыванию углерода в биомассе. Но в данный момент лесные площади массированно сокращаются. Облесение выкорчеванных площадей постепенно возмещает потери, но не дает нетто- прироста, пока они не восстановятся до предыдущего состояния, а это происходит со 100 — 200-летней задержкой. Старые деревья содержат существенно больше С, чем молодые, поэтому замена старых насаждений молодыми всегда (!) представляет собой потерю углерода. Следует помнить о том, что в долговременном балансе углерода решающим является не оборот, а- величина общих фондов (средняя продолжительность пребывания С в ландшафте). Быстрорастущие лесные посадки не вносят вклад в восполнение углеродного «долга» местности, хотя и могут служить ценным источником обновляемых ресурсов (замена ископаемым источникам углерода). Чтобы порядок величин замещающего потенциала был очевидным, на рис. 13.48 показано, сколько ископаемого углерода теоретически могло быть замещено биомассой, если бы для этого была введена в действие вся (!) вырубаемая за год древесина в Германии, Австрии или Швейцарии (другое применение древесины и техническое использование заменителей не предполагаются). Цифры свидетельствуют, что проблема не разрешима ни таким образом, ни созданием специальных плантаций биомассы, или биомасса редуцируется до немногих процентов от ископаемого углерода. Это не означает, что использование биомассы вместо ископаемого углерода не приветствуется и не приносит экологических и хозяйственных выгод. Никакой эффект биомассы не может хотя бы примерно приблизиться к потенциалу восстановления даже при маленьких (менее 5 %) квотах сбережения при потреблении горючих веществ, причем технически возможная экономия вместе с той, которая вытекает из действий человека, без сколько-либо значимого снижения уровня жизни может превышать 50 %. Не следует таким образом упускать из вида порядок величин, если в решении проблемы рассчитывать на роль растений.
Рис. 13.48. Потенциал замещения ископаемого углерода биологическими источниками углерода. Ежегодной выработке ископаемого углерода противопоставляется содержание его во всей вырубаемой национальным лесным хозяйством древесине или, как альтернатива, масса углерода, получаемая с плантаций биомассы (например, китайского тростника) на 10% современных посевных площадей