БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
13. РАСТЕНИЯ В ЖИЗНЕННОМ ПРОСТРАНСТВЕ
13.8. Биотические взаимодействия
Биоценозы Земли характеризуются не только основополагающими пищевыми цепями от продуцентов к консументам и редуцентам, но также и многими другими аспектами совместного существования и конкуренции организмов. Биотические взаимодействия, обычно называемые интерференцией, могут проявляться между индивидами одной популяции (вида) или между индивидами разных видов. Между экологически самостоятельными видами они обычно еще очень слабые, но могут углубляться вплоть до полной взаимозависимости (например, при симбиозе или паразитизме). Для популяционной биологии или популяционной генетики они имеют основополагающее значение. Ниже будут сопоставлены некоторые важные биотические взаимодействия; решающим критерием при этом будут позитивные (+), негативные (-) влияния на интенсивность возобновления или их отсутствие (0), которые оказывают друг на друга два партнера (А и В):

Биотические взаимоотношения между автотрофными растениями простираются от конкуренции до кооперации, причем пространственное вытеснение и борьба за свет и питательные вещества, равно как и за воду в почве, играют роль в такой же степени, как и изменения климата биоценоза (см. 13.3.1) или химические воздействия (аллелопатия, см. 9.5) и др. Некоторые из этих комплексных взаимоотношений можно понять на основе опытов в культуре с двумя или несколькими видами (рис. 13.49), другие же настолько сильно зависят от широкого комплекса влияний внешней среды, что в поле их можно разграничить только приблизительно путем наблюдений (пространственно-временные модели) или с помощью манипулятивного вмешательства.
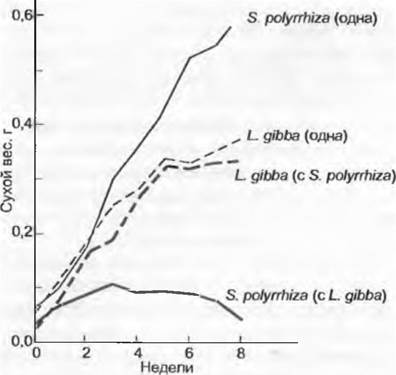
Например, в то время как ряска Spirodela (Lemna) polyrhiza всегда размножается интенсивнее, чем Lemna gibba, последняя в плотной смешанной культуре подавляет Spirodela в ходе конкуренции за свет (рис. 13.49). Насколько сильна корневая конкуренция за питательные вещества, показывает опыт, при котором молодые растения ели в березовом лесу удобряли маркированными фосфорными соединениями. Если березовые корни обрезают, то ели могут усваивать в 5 — 9 раз больше фосфора. Такие («zoot- trenching» — траншейно-корневые) опыты в тропическом лесу также документируют мощную корневую конкуренцию между всходами и взрослыми растениями.
При культивировании важнейших видов на газоне из Lolium реrеnnе высокорослый Trifolium pratense дает более высокую продукцию биомассы, чем низкорослый T. repens. Все виды вместе через год почти удваивают продукцию, но, если удаляют «мешающий» им Lolium реrеnnе, и в целом становятся в три раза более продуктивными, чем один T. pratense. Здесь очевидно явление кооперации, или комменсализма: связывание азота воздуха видами клевера с помощью клубеньковых бактерий явно способствует успеху других видов. Шпалерный кустарничек Loiseleuria (см. рис. 12.9, Е) в арктоальпийских обдуваемых ветрами местах часто существует совместно с кустистыми лишайниками (Cetraria и др.), которым он предоставляет условия для закрепления; в свою очередь возвышающиеся над ковром листьев талломы лишайников тормозят ветер, что создает для Loiseleuria более благоприятные микроклиматические условия.
Всходы и ювенильные растения обычно намного сильнее страдают от конкурентной борьбы, чем устоявшиеся взрослые растения. У лиственницы и сосны многие взошедшие семена выживают на примитивных почвах или на оголенных местах и, напротив, почти не выживают среди яруса высоких трав, мхов или кустарничков, которые разрастаются под более старыми деревьями. Здесь и световые условия неудовлетворительны, и влажность воздуха выше, одревеснение проростков меньше и поэтому значительно сильнее поражение грибами и поедание улитками. Так же и многие другие пионерные древесные растения препятствуют возобновлению лиственницы и сосны и способствуют сукцессии другими видами деревьев.
При аллелопатическом ограничении (см. 9.5) подроста особей других видов (но частично и своего вида) значение имеют продукты обмена веществ. Примеры этому найдены у водорослей Chlorella и Nizschia, у богатых терпеноидами Lamiaceae (например, зоны торможения вокруг видов Salvia в Калифорнии) и Myrtaceae (например, почти свободные от подлеска посадки Eucalyptus). Бросается в глаза «враждебность» к подлеску также у Robinia, Juglans и многих хвойных деревьев.
К отношениям между гетеротрофами и растениями относятся аллелопатия продуцирующих антибиотики актиномицетов и грибов против бактерий и обнаруженное К. Клеем с сотрудниками защитное действие эндофитных грибов в ткани листа против фитофагов. Существенны взаимодействия автотрофных растений с грибными симбионтами в лишайниках (см. 9.2.4) и микоризе (см. 9.2.3), а также с комменсалами, например, с многочисленными сапрофитными бактериями и грибами, которые живут на опадающих частях растений, или с паразитами (многие, бактерии и грибы, некоторые покрытосеменные; см. 9.1.1, бокс 11.4).
Рис. 13.49. Размножение двух свободноплавающих видов Lеmnасеае (Spirodela polyrhiza и Lemna gibbа) в чистой культуре и смешанной культуре в условиях конкуренции
Значительные изменения растительности могут возникать, когда определенные виды деревьев сильно поражаются или полностью элиминируются грибными заболеваниями; за последние десятилетия, например, это касается полевого вяза в Европе (Ulmus minor; гриб — Ophiosloma ulmii, см. 11.2) и восточно-североамериканского каштана (Castanea dentata\изначально поражается до 60 % частей дерева; гриб — Endothia parasitica, занесенный в 1904 г. из Китая; см. Diaporthales). Остается лишь ожидать, что произошел отбор резистентных биотипов этих деревьев, которые могли бы вновь завоевать утерянные видом территории.
Особенно многосторонни и экологически значимы биотические взаимоотношения между растениями и животными. В первую очередь здесь следует назвать фитофагов, т. е. травоядных животных, как консументов первого порядка. При этом причиной очень значительных повреждений и изменений растительного покрова могут стать насекомые (тли, короеды, пяденицы), улитки или млекопитающие (мелкие грызуны, кролики, жвачные животные), поедающие или высасывающие вегетативные органы, цветы и во многих случаях семена. Обусловленное этим «давление» отбора приводит к образованию многочисленных защитных механизмов: колючки, шипы, жгучие волоски, иглы кристаллов, горькие и ядовитые вещества и т. д. (см. 4.2.6, 9.4.1, 11.168). Особый случай — образование животными галлов (см. 9.1.1). Симбиотические связи с животными проявляются у семенных (а также и у низших) растений особенно в области биологии цветения, плодоношения и семеношения (см. бокс 11.4). Многие бактерии и грибы паразитируют на животных. Немногие грибы и покрытосеменные специализировались как «ловцы животных» (см. 9.1.2).
Пастбищные животные при стравливании повреждают в первую очередь молодые проростки древесных растений и благоприятствуют тем самым хорошо регенерирующим злакам и крупным травам лугов. Дальнейшие изменения местообитаний происходят путем вытаптывания (уплотнение почвы, механические повреждения) и удобрения. Как следствие этого распространяются растения — так называемые «вредители пастбищ», которые скот обходит, например, в Центральной Европе Juniperus commimis, Berberís vulgaris, Prunus spinosa, Ononis spinosa, Eryngium campestre, Cardales sp., Cirsium sp., Nardus stricta с твердыми и колючими побегами, а также виды Rumех, Ranunculus, Euphorbia, Apiaceae, Lamiасеае, Liiiales (например, Colchicum autumnale), в них содержатся горькие, ароматические или ядовитые вещества.
История развития многих родственных групп покрытосеменных была успешной, а их формообразование богатым, очевидно, потому, что они выработали действенные химические вещества, защищающие от поедания животными, например, Capparales с их гликозидами горчичного масла, многие Gentianales с индол-алкалоидами или Solanасеае с тропан-алкалоидами (см. 9.4.1). Только определенные группы фитофагов могут потреблять эти защитные вещества без вреда для себя и даже специализируются на соответствующих растениях (например, бабочки Pierinae на Capparales). Бабочка-монарх (Danaus plexippus) встраивает получаемый из своих кормовых растений (Asclepiadaceae) ядовитый гарденолид-гликозид даже в тело гусеницы и взрослого животного и поэтому несъедобна для своих врагов.
В качестве примера еще недостаточно исследованных взаимодействий между растениями и муравьями (см. зоохория) можно привести неотропические виды Acacia (например, A. cornígera). У этого дерева из влажных тропических лесов развит симбиоз с агрессивными муравьями (Pseudomyrmex ferruginea): оно служит им жилым помещением, источником питания и экстрафлорального нектара (см. рис. 11.251) и при этом очень успешно «использует» муравьев, как защиту от всех фитофагов. Муравьи даже подрезают и удаляют разрастающиеся лианы и конкурирующие соседние растения, чтобы их растение-хозяин могло лучше развиваться. Эффективность этого симбиоза видна из сравнения с другими акациями, которые не заселяются муравьями: они сильно подавлены в росте и искривлены. Аналогичные отношения имеются между муравьями и пионерными деревьями рода Cecropia.
Очень существенна для многих растений потеря семян из-за животных.Fagгs sylvatica может успешно размножаться только в урожайные годы с повышенной семенной продуктивностью. Такие годы повторяются через неодинаковые промежутки времени; это следствие того, что паразитирующие в семенах насекомые в своих циклах развития не могут ориентироваться только на урожайные годы. Неотропические бобовые выработали две защитные стратегии против жуков (Вruchidае), поедающих их семена: либо они продуцируют неядовитые, но многочисленные и мелкие семена, из которых хотя бы часть остается нетронутой, либо образуют более крупные и немногочисленные семена, но содержащие ядовитые вещества (см. 11.2, Fabales).
Все эти положительные и отрицательные взаимодействия влияют на рост популяций, участвующих в создании биоценоза. Одни виды становятся доминирующими, другие остаются подчиненными или исчезают, что свидетельствует о лабильном или более, или менее стабильном состоянии равновесия. Такие примеры можно описать математически и симулировать с помощью компьютерных моделей.
Конкурентная борьба между двумя видами становится тем острее, чем ближе их экологические потребности. Надолго их совместное существование в одной и той же экологической нише невозможно (см. 12.1). Поэтому при негативных биотических взаимодействиях мы постоянно находим, что существующие в сообществе виды «уклоняются» от конкуренции: в пределах генетически установившейся в чистой культуре широты нормы реакции видов виды «сдвигаются» в ту или иную сторону при произрастании в смешанной культуре, т. е. под влиянием биотических взаимодействий их распространение таково, что перекрывание с амплитудами и областями оптимума конкурентов оказывается минимальным.
Этот принцип уже обсуждался в разделе 12.1. Он применим, например, и к важным луговым злакам Центральной Европы: по отношению к высоте уровня грунтовых вод (влажности почвы) в чистой культуре они обнаруживают очень близкие оптимумы роста, в то время как в смешанной культуре оптимумы роста отстоят друг от друга достаточно далеко. Bromuserectus и многие другие «ксерофильные» виды в действительности очевидно, не «сухолюбивые», а только более «суховыносливые». Также и Oxallis acetosella не «тенелюбива», а «теневынослива». Многие реликтовые средиземноморские виды лишь потому ограничиваются в своем распространении недоступными трещинами скал, что во всех других местообитаниях они стравливаются козами и овцами.
Экологическая позиция и широкая или соответственно узкая амплитуда экологических потребностей видов (эвритопные, соотв., стенотопные) очень зависят также от их биоценотических партнеров, она относительна. Во всяком случае эти биотические взаимоотношения между видами очень разнообразны, комплексны и системно объединены друг с другом и прочими факторами местообитания; это решающим образом способствует стабильности и саморегуляции экосистем.
Виды, которые в своих исконных экосистемах занимают подчиненное положение и «находятся под контролем», могут в «чужих» экосистемах становиться агрессивными «сорняками», поскольку там отсутствуют их естественные враги. Примерно так произошло с европейско-атлантическим Ulex europaeus в Новой Зеландии, европейским Hypericum perforatum в Северной Америке или неотропической Opuntia inermis в Австралии. Только намеренный занос питающейся Opuntia венесуэльской моли Cacto- blastis cactorum ликвидировал этот вредный сорняк на площади более 120 млн га в течение нескольких лет. Похожая ситуация с водяной чумой Elodea canadensis в Европе. Напротив, интродуцированные животные (например, козы и кролики) в значительной степени уничтожили малоустойчивые к выпасанию островные флоры, например, Гавайских островов, о-ва Св. Елены. Многие виды биоценоза играют несколько экологических ролей, так, Viscum album (омела) для деревьев-хозяев — полупаразит, для разносящих семена птиц — симбионт, для различных насекомых-фитофагов — даже хозяин.
Давление паразитов и других врагов на вид тем сильнее, чем больше этот вид стремится к доминированию и образует большие сомкнутые популяции. Например, гомогенные искусственные еловые посадки намного более восприимчивы к носящим характер эпидемий вспышкам численности вредителей (короедов, пядениц), чем естественные смешанные насаждения ели с другими древесными породами. Богатое видовое разнообразие многих лесов от умеренно-теплых до тропических, видимо, объясняется тем, что каждый древесный вид, разрастающийся за счет другого, вскоре вновь редуцируется из-за богатой паразитической фауны и флоры.