БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
14. ЭКОЛОГИЯ ПОПУЛЯЦИЙ И РАСТИТЕЛЬНЫХ СООБЩЕСТВ
14.3. Экология растительности
Если анализировать отдельные, локально отграниченные группировки растений, то снова и снова обнаруживается совместное произрастание характерных таксонов.
В большинстве случаев речь идет не о случайном соседстве, а о совершенно определенном, обусловленном внешней средой отборе видов из всей совокупности местной флоры (см. рис. 14.1). При этом качественный и количественный состав сообщества часто удивительно точно отражает соответствующие условия окружающей среды.
Растительные сообщества (англ, plant communities) являются комбинациями популяций различных видов, находящихся в зависимости от абиотической среды и биотических взаимодействий. Их формирование подчиняется закономерной последовательности (сукцессии) от пионерных группировок до зрелых «конечных» сообществ, при прерываниях которой (по всей площади или точечных) развитие идет, начиная с характерной промежуточной ступени опять же в сторону типичного для данных условий конечного сообщества — «климакса» (см. рис. 14.14). Нарушения (ураганы, пожары, затопление, вытаптывание животными) могут отклонить сукцессию в сторону другого направления или остановить ее на промежуточной ступени на долгое время. Несмотря на то что биосфера представляет собой сопряженное целое, существующие по соседству растительные сообщества большей частью все же хорошо отличаются друг от друга, имея границы или переходные зоны (постепенные переходы — экотоны, резкие переходы — экоклины; обычно употребляются эти английские выражения). Примеры — переходы от леса к травяной растительности или от растительности влажных местообитаний к сухим. Причины успеха (присутствия) или неуспеха (отсутствия) вида заключаются в его способности успешно размножаться в данных конкретных условиях, причем критическая для существования фаза может соответствовать любому отрезку жизненного цикла — от фазы закрепления всходов до успешной продукции диаспор. это уже сфера деятельности популяционной экологии (см. 14.1.3). Предмет экологии растительности — пространственная и временная структура растительных сообществ и выявление при этом функционально значимых типов и их связей. Это также и построение гипотез для объяснения с
точки зрения происходящих процессов структуры взаимодействий как между видами, так и внутривидовых (см. гл. 13). Одна из важных задач экологии — это хотя бы простое описание и картирование растительности.
В зависимости от задач, территории и масштаба, а также от пространственных и временных подходов имеется много различных возможностей для того, чтобы исходя из полевых описаний определить абстрактные типы растительности различного ранга:
✵ по видовому составу (таксономии) — растительные сообщества;
✵ по господствующему внешнему облику (морфотипам) — растительные формации;
✵ по пространственным связям (сходство местообитаний/взаиморасположе- ние) — комплексы растительности;
✵ по временной последовательности (состояние развития) — сукцессионные ряды.
14.3.1. Структура растительных сообществ
Поверхностная систематизация растительного покрова уже дает возможность осознать многообразие возможных подходов к анализу лежащих в его основе моделей и связей. В смешанном буковом лесу бросается в глаза структура популяции доминирующего бука, в которой чередуются всходы, молодые растения, доминирующие более старые деревья и, наконец, отмирающие (падающие от ветровала или поражения паразитными грибами) или вырубающиеся, а также упавшие гниющие стволы. Подчиненный и мозаично распределенный в зависимости от небольших колебаний рельефа и почв или от световых условий нижний ярус состоит из разнотравья, граминоидов, мхов и лишайников (рис. 14.27). Геофиты, цветущие в весеннее время, такие, как Anemone nemorosa, летом и осенью полностью исчезают с поверхности земли, зато появляются плодовые тела шляпочных грибов, которые связаны с буком в качестве микоризных симбионтов или вместе с бактериями и мелкими беспозвоночными участвуют в процессах разложения листового опада. Другие подчиненные элементы биоценоза находятся на основаниях древесных стволов (листостебельные мхи) и на их коре (аэрофильные водоросли и накипные лишайники).
Рис. 14.27. Части биоценозов (синузии) в центрально-европейском смешанном буковом лесу в конце лета:А — впадины, заполненные листовым опадом, с относительно влажными нейтральными перегнойными почвами (с бактериями, грибами, мелкими беспозвоночными), по краям их — индикаторы влажности и соответственно богатства почвы: а — живучка ползучая (Ajuga reptans), b — лесная фиалка (Viola reichenbachiana); В — микроповышения, свободные от листового опада, с более сухими слабокислыми, бурыми лесными почвами, с наземной моховой и лишайниковой синузиями: с — гипновый мох (Нурnum cupressiforme), d — белый мох (Leucobryum glaucum), е — лишайник кладония (Cladonia pyxidata), f — микроповышения с плодовыми телами млечника (Lactarius vellereus; образует микоризу с Fagus) и с указывающими на кислотность и бедность питательными веществами почвы травянистыми розеточными растениями (гемикриптофитами), g — ожика беловатая (Luzula luzuloides), h — ястребинка лесная (Hieracium murorum), i — щучка извилистая (Deschampsia flexuosa): С — основание ствола с различными листостебельными мхами (Нурnиm, Plagiothecium)] D — кора бука с синузией эпифитных водорослей (Pleurococcus) и накипными лишайниками (Graphis scripta и др.); Е — древесный пень, заселенный грибами (плодовые тела трутовика Coriolus versicolor)

Подобная комплексная инвентаризация может быть проведена только путем разносторонних (и часто очень трудоемких) исследований. На правильно выбранной пробной площади необходимо сначала охватить видовой состав и состав жизненных форм растительного сообщества, а также проанализировать его ярусное строение, чередование крупных и мелких биотопов и периодичность развития. Такие общие описания растительности и ее состояния впоследствии являются предпосылкой для выделения и описания растительных сообществ.
Слово «структура» означает в немецком языке любую форму пространственного и иерархического подчинения или систему связей. Если в англоязычной специальной литературе речь идет о «community-» или «canopy structure» (структуре сообщества или покрова), то подразумевается в большинстве случаев структура геометрическая, в смысле «архитектуры». Применяемое в немецком языке обозначение видового состава растительного сообщества — структура насаждения = таксономическая структура соответствует аналогичному обозначению в английском «community composition» — состав сообщества.
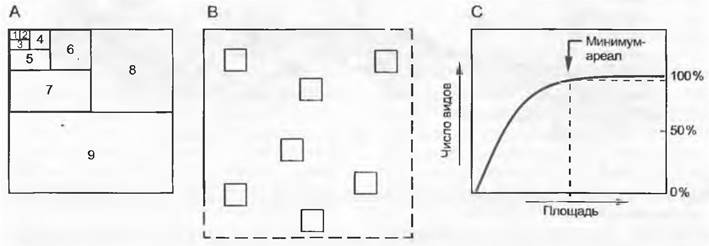
Исходным пунктом для анализа растительности служит инвентаризация ее небольших выборочных образцов. Выбор и размер таких пробных площадей проводят в зависимости от того, какой биоценоз (или какая его часть) должен быть охарактеризован. Чтобы учесть все характерные деревья среднеевропейского леса, необходим минимум-ареал размером около 500 м2, а для богатых видами влажных тропических лесов площадь, возможно, должна быть около 1 га. Напротив, при описании лугов и дерновинных травяных сообществ достаточно уже 10— 100 м2, а для сообществ мхов и лишайников — 0,1 — 4 м2. На альпийских лужайках на гранатных сланцах в северной части Скандинавии на площади около 1 м2можно найти до 50 видов покрытосеменных. Если же площадь увеличить до 100 м2, то количество видов может возрасти до 60, а до 1 км2 — до 80. Таким образом, на площадке размером 1 м2 уже представлено 2/3 видового списка района. Наличие данных по зависимости числа видов от площади повышает ценность данных по биоразнообразию (например, по видовым числам). Диаграмма зависимости числа видов от площади (рис. 14.28) позволяет определить, с площади какого размера (минимум-ареала) можно получить достоверную выборку, содержащую более 95 % общего списка. При этом площади разной величины можно получать путем постепенного расширения стартовой площадки или же путем добавления других площадок, распределенных по общей пло
щади случайным образом. Показания эквивалентны не полностью. Как правило, второй метод объективнее и предпочтительнее.
Рис. 14.28. Минимум-ареал, или наименьшая площадь, которая содержит полный видовой набор растительного сообщества (> 95% всех видов). Это можно выявить путем последовательного удвоения пробных площадей (А — метод одной площадки) или накопления добавочных данных, но на площадках такого же размера (В — метод многих площадок). Результат выражен на кривой накопления (С), которая показывает, при какой величине пробной площади или при каком количестве повторяющихся площадок можно не ожидать существенного (> 5%) увеличения видового богатства. Типичный минимум-ареал при одной пробной площади для малопродуктивного луга или альпийской лужайки — 10 — 25 м2, для травяного яруса в лесах — 100 — 200 м2, для почти ненарушенного умеренного леса — 500 — 1 000 м2, для влажного тропического леса — более 1 га. Характер кривой зависимости видового числа от обследованной площади дает представление о гомогенности растительного сообщества
Для классификации растительных сообществ в целях определения типов сообществ (ранее это называлось фитосоциологией) применяется ряд параметров, среди которых выделяются две группы. Одна группа описывает характер произрастания особей видов в пределах пробной площади, которая должна по меньшей мере соответствовать величине минимум-ареала. Другая группа охватывает показатели, которые выводятся из сравнения большого числа таких площадей.
В пределах пробной площади можно определить или измерить: плотность особей, или обилие (число особей на единицу площади), участие в покрытии (% покрытия почвы вертикальной проекцией особей вида, так называемое доминирование в пространстве) и встречаемость (повторяемость встречи вида на площадках, заложенных внутри пробной площади, %). Высокая встречаемость более важных (обильных) видов указывает на высокую гомогенность растительного сообщества (в противоположном случае — гетерогенность).
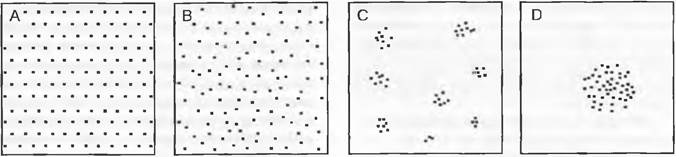
К качественным признакам можно отнести общественность (социальность, произрастание группами или поодиночке) и распределение (регулярное или нерегулярное размещение; признак, близкий к встречаемости. рис. 14.29). В качестве меры оценки «успешности» вида (англ. vigor), с учетом его продуктивности, можно также применять индекс жизненности.
Рис. 14.29. Примеры распределения видов в растительном сообществе, которые находят отражение в различной встречаемости в пределах пробной площади:
А — регулярное распределение (случайно встречается в засушливых областях для «выдерживания дистанции», а также в моноклональных и антропогенных сообществах): В — случайное распределение (например, на неструктурированных рудеральных участках, в других случаях скорее редко); С — распределение группами или кучками (часто встречается на естественных лугах и в первичных лесах); D — очаговое распределение, ограниченное отдельными пятнами (англ. patches) часто при точечных нарушениях или для клонов
Из сравнения большого числа таких описаний можно определить константность вида (вероятность его встреч, как повторяющихся появлений на разных пробных площадях, аналог показателя встречаемости внутри пробной площади). Степень привязанности отдельных видов к определенным сообществам обозначают как их верность. Кроме того, виды с очень высокой верностью (и обычно с более высоким обилием) типичны и называются характерными или характеризующими видами; другие, вполне типичные, также верные, но все же не столь строго связанные с этой ассоциацией, обозначаются как сопровождающие, а остальные — как случайные. Виды, которые делят сообщества одного ранга на подгруппы (например, субассоциации), называют дифференциальными, или разделяющими видами. Не будучи доминантами, они имеют сильную и специфическую сопряженность (верность) с соответствующей подгруппой и высокое постоянство в ее пределах, но не на ближайшей ступени более высокого социологического ранга. Дифференциальные виды,
как правило, хорошие индикаторы определенных экологических условий.
Чаще всего списки видов (так называемые перечни — releves) сопровождаются оценкой доминантности (покрытия) или плотности особей отдельных видов (доминантность и обилие объединяются в единый показатель мощности вида). Такая альтернатива возникает из-за необходимости разделять виды с узкими, вертикально направленными листьями и с широкими, плоско расположенными Виды, образующие листовые розетки, могут, несмотря на высокое покрытие, иметь сравнительно небольшую индивидуальную плотность. Некоторые злаки при высокой плотности побегов достигают сравнительно невысокого покрытия. У клональных растений или растений, образующих большое число побегов, по чисто техническим причинам оцениваются не генетические индивиды (генеты), а побеги (раметы). Значения обилия (= плотности) обозначаются обычно с помощью приблизительной классификационной шкалы (табл. 14.2).
Таблица 14.2. Оценочная классификация значений обилия
Класс |
Покрытие, % |
Обилие |
5 |
> 75 |
Число особей любое |
4 |
50 -75 |
Число особей любое |
3 |
25 - 50 |
Число особей любое |
2 |
5 - 25 |
Особи мелких форм роста, очень многочисленные |
1 |
< 5 |
Достаточно много |
+ |
Скудно |
Особи мелких форм роста, очень мало |
г |
Редко |
Почти единичные, редкие и вне пробной площадки |
Эта ранговая шкала, составленная на оценочной основе, в практике часто для специальных целей еще дополняется или модифицируется в нижней области. По существу, она соответствует видоизмененным показателям оценки покрытия, благодаря которым доминирующие виды часто оцениваются несколько ниже, а редкие — выше. Точные количественные данные можно получить, только срезав всю растительность, пересчитав и взвесив все особи. Очень хорошая количественная оценка покрытия достигается методом «точечных квадратов», при котором в плотную, размеченную координатами рамку опускаются иглы и отмечается первое касание иглой особи одного вида в точке координатной сетки.
Каждое описание растительности снабжается данными, характеризующими местообитание, — это положение в рельефе, высота над уровнем моря, экспозиция, крутизна склона, тип почвы, характер материнской породы, хозяйственное использование и др. Постепенное или резкое изменение видового состава вдоль определенного градиента внешней среды (например, высоты над уровнем моря, влагообеспеченности. степени засоления, pH, светового довольствия) лучше всего может быть представлено в форме трансекты растительности (см. рис. 13.28). Она служит отправной точкой для по возможности наиболее объективного разграничения растительных сообществ. Часто хотя бы приблизительно ценность охарактеризованных признаков заключается в том, что, с одной стороны, эти признаки (особенно отношение покрытие/обилие) позволяют «взвесить» присутствие вида, с другой, благодаря большому количеству повторностей отдельные относительные неправильности все же не нарушают слагающейся достаточно четкой общей картины, это позволяет прибегать к количественным сравнениям.
Таблица 14.3. Спектры жизненных форм (% участия соответствующих видов) некоторых важных формаций и экологических рядов (см. рис. 14.19, 14.42)
Область распространения |
Фанерофиты |
Хамефиты |
Гемикриптофиты |
Геофиты |
Терофиты |
Во всем мире |
46 |
9 |
26 |
6 |
13 |
От теплого до холодного (гумидного) климата |
|||||
Тропические дождевые леса |
96 |
2 |
2 |
||
Субтропические лавролистные леса |
66 |
17 |
2 |
5 |
10 |
Тепло-умеренные лиственные леса |
54 |
9 |
24 |
9 |
4 |
Холодно-умеренные хвойные леса |
10 |
17 |
54 |
12 |
7 |
Тундра |
1 |
22 |
60 |
15 |
2 |
От влажного до сухого (умеренного) климата |
|||||
Лиственные леса |
34 |
8 |
33 |
23 |
2 |
Лесостепь |
30 |
23 |
36 |
5 |
6 |
Степь |
1 |
12 |
63 |
10 |
14 |
Полупустыня |
59 |
14 |
27 |
||
Пустыня |
4 |
17 |
6 |
73 |
Решающим признаком для общего габитуса каждого растительного сообщества являются морфология (форма) видов, а также их жизненные формы и формы роста (см. 4.2.4). По В. Раух (W. Rauh), форма роста — это принцип организации, или план строения, жизненная форма — это то, что может реализоваться от случая к случаю в жизненном пространстве в пределах того широтного пояса, условия которого это допускают. Форма роста дерева таким образом вполне может «спрессоваться» в определенных условиях среды в форму кустарника. Правда, разница в терминах в литературе почти не играет роли, оба понятия используются как синонимы (в немецкоязычной литературе это жизненная форма, в английской — форма роста, growth form). В разных зонах и основных типах растительности Земли существуют очень разнообразные жизненные формы (см. рис. 14.42, табл. 14.3), участие которых в сложении растительности обусловливает ее вертикальную стратификацию. Только самые первичные пионерные биоценозы, а также биоценозы, находящиеся в экстремальных условиях, имеют мало слоев (ярусов) или вообще однослойны. В большинстве естественных лесов есть:
✵ древесный ярус, часто с лианами и эпифитами, также разделяющийся на несколько ярусов;
✵ кустарниковый ярус, включая молодые деревья;
✵ травяной ярус с кустарничками и полукустарниками, включающий всходы деревьев;
✵ моховой, лишайниковый (напочвенный) ярус.
Этому наземному вертикальному расчленению растительности соответствует менее исследованная подземная ярусность корневых систем в ризосфере (см. рис. 13.24). Очевидно, что дифференцированное освоение воздушного пространства и почвенного слоя создает возможность для лучшего использования ресурсов (свет, влага, питательные вещества в почве, см. 14.2.4 2).
Традиционное деление растений на мелко и глубоко укореняющиеся очень упрощено и функционально плохо обосновано. Почти у всех многолетних растений имеются как поверхностные, так и глубоко проникающие корни, но их соотношение варьирует у разных видов и при этом сильно зависит от доступности влаги и питательных веществ. Небольшое число глубоких корней (которые часто не заметны) гарантируют растению даже во время критических периодов минимальное снабжение влагой (по меньшей мере для покрытия очень небольшого кутикулярного расхода влаги через устьичные отверстия), поверхностные корни обеспечивают усвоение большей части питательных веществ из особенно биологически активного верхнего слоя почвы. В периодически сухих районах глубина проникновения корней тесно коррелирует с сезонным ритмом активности побегов. Виды, которые остаются активными (зелеными) на время сухого сезона, имеют более глубокие корни, чем те, которые сбрасывают листву. На центрально-европейских умеренно сухих лугах, расположенных на известняках более 80 % всех корней располагаются в верхних 20 см почвенного профиля, хотя отдельные корни проникают вглубь до 6 м, что видно на отвесных обнажениях. Максимальную глубину корней более чем на 15 м можно наблюдать скорее, как правило, нежели в виде исключения, в большинстве периодически засушливых областей Земли (табл. 14.3).
Растительный покров структурирован также и в горизонтальном направлении. В зависимости от величины свободной от растений поверхности различают открытую или сомкнутую растительность. Даже внутри насаждения, кажущегося однородным, часто можно наблюдать дифференцированное горизонтальное распределение видов в форме определенного рисунка, мозаики или коалиций (определенные виды группируются друг с другом или отстоят далеко, см. рис. 14.29). Уже незначительные повышения или впадины микрорельефа (см. рис. 12.13) обусловливают различия в снабжении влагой и питательными веществами (см. 13.6.2) и отсюда —
в обилии видов. Сам по себе растительный покров также создает различные микробиотопы (см. рис. 14.27). Особое значение имеют окна в насаждении (англ. gaps), образующиеся в результате выпадения особей, так как при этом возникает свободное пространство для поселения новых особей. Самое удачное типичное описание развития лесов ориентировано как раз на динамику в таких окнах (англ. gap dynamics, gap models). В нем речь идет прежде всего о продолжительности существования таких окон и сукцессии, которая в них развертывается. Рисунок 14.30 иллюстрирует цикл от возобновления (А) к оптимуму (В) и разрушению (С) лесного насаждения. Этот цикл зависит от длительности жизни доминирующих особей. В так называемых вырубочных окнах возобновление происходит не сразу, а протекает через стадии недолговечных пионеров, затем быстрорастущих, промежуточных (серийных) видов и только в последнюю очередь — молодых растений видов, изначально доминировавших (см. 14.3.2). В то же время закрепление молодых растений может происходить успешнее непосредственно под защитой других особей (англ. facilitation — облегчение; так, кактусы лучше поднимаются под защитой пустынных кустарников, что хорошо подтверждается на примере кактуса сагуаро — Camegia gigantea).
Рис. 14.30. Циклическая регенерация горного елово-пихтово-букового девственного леса в Восточных Альпах (Ротвальд близ Лунца, 1 000 м): А — фаза омоложения с богатым подростом в «окнах» (места ветрового вывала); В — фаза оптимума с плотно сомкнутыми кронами и преобладанием хвойных пород; С — фаза разрушения перестойного сообщества (много мертвых деревьев, как стоящих, так и упавших, повышается участие бука, вновь появляется подрост). Вертикальный и горизонтальный профили растительности: • ель (боковые ветви с темной заливкой); пихта (боковые ветви без заливки); бук (лиственные кроны изображены схематически); упавшие стволы, подрост— заштрихованы

Наконец, сильно упорядочивающий/ структурирующий элемент — это еще и временная последовательность, периодизация, развития отдельных растений и сообществ в целом. Прежде всего важны такие фенофазы, как разворачивание листьев, цветение, плодоношение, старение листьев (рис. 14.31). В своей совокупности они обусловливают формирование в разные сезоны года различных «аспектов» (внешнего вида) растительного сообщества.
Рис. 14.31. Сезонное развитие (фенология) характерных видов влажного дубово-букового леса в северо-западной части Германии. Черным цветом показано развитие листьев этого года, горизонтальными линиями — перезимовавших листьев, вертикальными — цветков
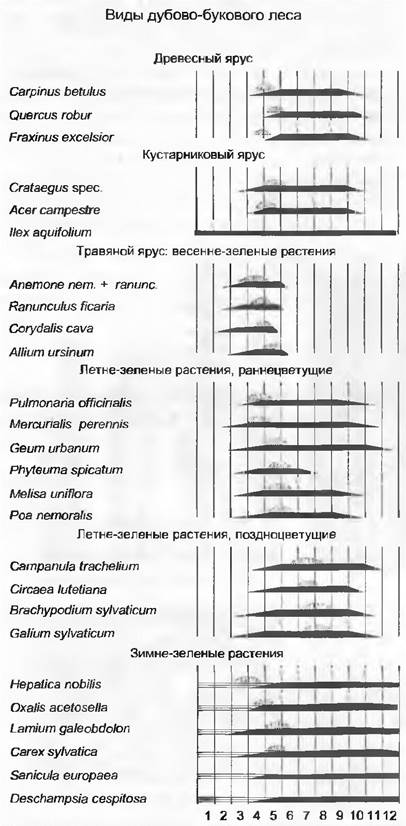
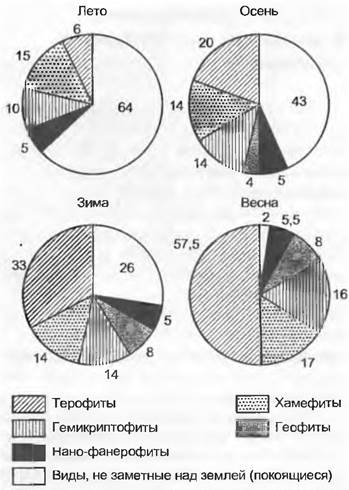
При сравнении сезонного развития видов влажных дубово-буковых лесов (см. рис. 14.31) отмечается, что листва на лиственных деревьях, более «открытых» для зимних морозов, разворачивается поздно. Вечнозеленым остается только IIex. Период до разворачивания листвы или во время разворачивания более благоприятен для цветения, особенно для ветроопыляемых (анемофильных) деревьев. Этот светлый период используют также требовательные к свету геофиты, которые развиваются только в короткое весеннее время. Остальные виды травяного яруса теневыносливы; произрастая непосредственно у поверхности почвы (защита от мороза), многие из них сохраняют листья в течение всей зимы. Для средиземноморской растительности в сезонных аспектах особенно характерно частичное или полное отмирание в летнее время терофитов (однолетников) и геофитов (луковичных и клубневых растений, рис. 14.32).
Рис. 14.32. Сезонные изменения участия растений разных жизненных форм в сложении растительности средиземноморской каменистой пустоши (Вrachypodietum ramos близ Монпелье, юг Франции). Проценты рассчитаны от общего числа видов (111)
Итак, растительным сообществам свойственна пространственно-временная упорядоченная структура. Часто их большое видовое богатство и разнообразие жизненных форм и форм роста пространственно, функционально и сезонно связаны с наличием имеющихся экологических ниш, причем отношения между видами могут быть от взаимно дополняющих до зависимых и нарушения положительно влияют на сосуществование (см. 14.2.4.1).
14.3.2. Формирование и динамика растительных сообществ
Растительный покров находится в состоянии постоянного изменения (см. выше) и появляется на одних и тех же местообитаниях с различающимися в зависимости от сукцессионнойфазы набором видов, спектром жизненных форм и доминантами. Эго хорошо видно из анализа, проведенного через длинные временные интервалы, и наблюдений на постоянных площадках длительного мониторинга (англ. permanent plots, permanent quadrants; рис. 14.33). Но хорошую картину происходящей сукцессии дают и сравнения различных участков сформировавшейся растительности на сходных местообитаниях (рис. 14.34).
Рис. 14.33. Последовательная смена растительности (сукцессия) в течение 4 лет на площадке длительного наблюдения (1 м2), изначально не заросшей. Высохший торф верещатникового болота (Хилден, Рейнланд): а — Agrostis sp.; b — Molinia caerulea, c — Sphagnum papillosum, d — S. auriculatum, e — Erica tetralix, f — Juncus bulbosus, g — J. squamosus, h — Dicranella cerviculata-, i — Carex panacea-, j, k — Eriophorum angustifolium, I — Cerastium sp.; m — Polygala serpyllifolia: n — Rhynchospora alba
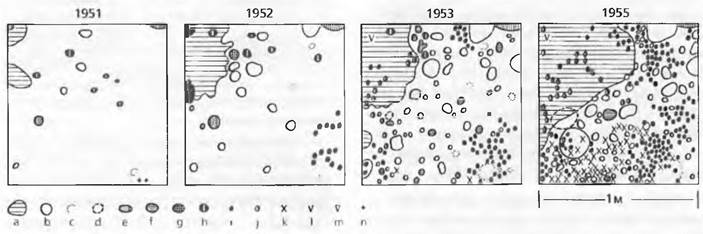
Рис. 14.34. Повторное зарастание лесом залежи в умеренной зоне (Северная Америка; Брукхэвен, Нью-Йорк). Через приблизительно 8 лет разнотравье и граминоиды сменяются летне-зелеными кустарниками, последние через 30 лет сменяются лиственными лесами, которые к 150 годам стабилизируются как климаксовый лес с летне-зелеными дубами и соснами. В ходе этой прогрессивной сукцессии увеличиваются первичная нетто-продукция (А, о—о) и биомасса растительного сообщества (В, □—□) вплоть до стадии климакса; напротив, падают число видов сосудистых растений (С, ▲—▲) после максимума в поздней фазе травяного сообщества и число адвентивных видов (D, ∆—∆), элиминирующихся на кустарниковой стадии в результате конкуренции
Реконструкция развития растительности в течение длительного времени проводится на основе ископаемых остатков (пыльцы) и свойств почвенного профиля (ископаемые почвенные горизонты и «горелые» горизонты). В семиаридных областях информацию относительно исторических смен, доминирующих СЗ- и С4-растений можно проследить путем анализа соотношений изотопов углерода в гумусе (см. 13.7.4). Поучительны опыты, в которых можно наблюдать сукцессию после исключения нарушений (огораживание от травоядных животных, препятствование пожарам).
Для первоначального поселения и, соответственно, изменения видового состава требуются элементы расселения (см. диаспоры), привнесенные извне или имеющиеся в покоящемся состоянии в семенном банке почвы. Установлено, что на 1 м2 пахотной почвы содержится до 50 000 жизнеспособных семян. В зависимости от зрелости системы в ней могут утвердиться популяции только определенных видов. На открытых участках с неразвитыми почвами (например, участки в зоне отступания ледников, галечные отмели, дюны) сначала формируется типичная пионерная растительность. На нарушенных поверхностях (на мусорных участках, на обочинах дорог) вначале поселяются так называемые рудеральные виды (см. рис. 14.13); для регулярно нарушаемых, используемых в сельском хозяйстве территорий характерна сегетальная флора (флора посевов и залежей). Каждая сукцессия связана с каким-либо изменением местообитания. Причины этого могут быть как внешними (аллогенная сукцессия), так и лежащими внутри самого сообщества (автогенная сукцессия).
Так, постепенное становление пойменной растительности вдоль горных ручьев (рис. 14.35) после отложений на поверхности гальки, песка и ила (намыв берега) — аллогенная сукцессия. В стоячих водоемах (рис. 14.36), напротив, в образовании поверхности заселения преобладают органогенные отложения, образованные из самой растительности (зарастание), — это автогенная сукцессия. Из-за периодической стабилизации состояния среды (например, регулирование рек, относительно стабильное состояние водной поверхности вдоль скальных берегов) сукцессия может приостановиться, достигнув стабильного зонирования растительности. Аллогенные смены
обусловлены изменениями климата или субстрата, или нарушениями, автогенные управляются видами, имеющими особое значение в строении сообщества. В закреплении дюн это побегообразующие злаки, например, Еlymus и Аmmophila (см. рис. 13.24; раздел 15.1.1), в зарастании водоемов — тростники и крупные осоки, в формировании среднеевропейских лесов — Fagus sylvatiса. поскольку эти виды «затеняют» другие и в большей степени влияют на формирование органогенного горизонта почвы.
Рис. 14.35. Схема последовательного распределения растительности в среднем течении реки в предгорьях Альп в зависимости от высоты воды и седиментации (наносов)
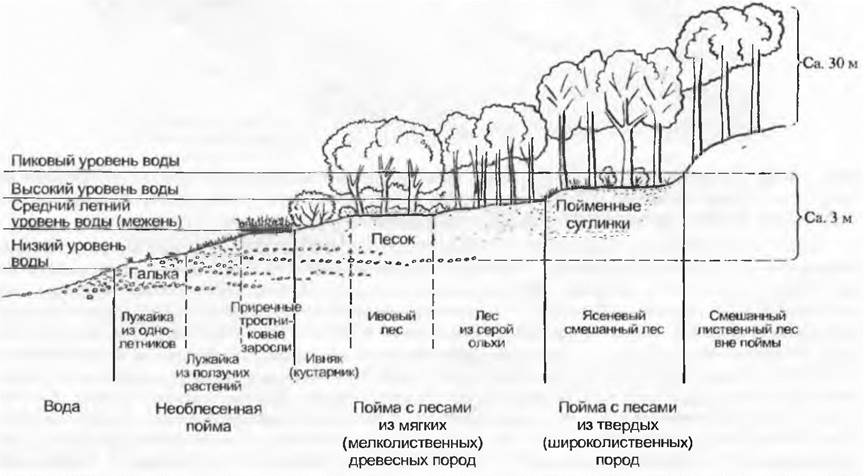
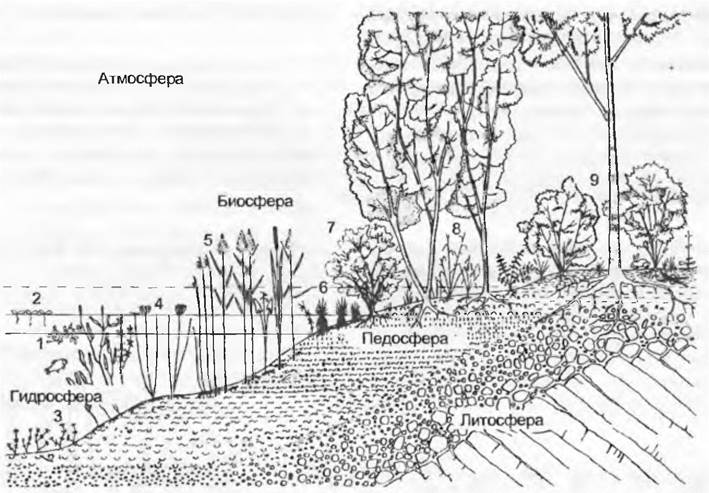
Рис. 14.36. Профиль растительности вдоль зоны зарастания эвтрофного озера в Центральной Европе (даны только названия родов, так как часто они представлены только одним широко распространенным видом в последовательности, соответствующей уменьшению глубины воды): 1 — свободно плавающие подводные растения, Utricularia (см. вставку 4.4, рис. В); 2 — свободно плавающие растения, Lemna (ряска), Hydrocharis (водокрас); 3 — прикрепленные подводные растения, Chara (вертикально растущая зеленая водоросль), Myriophyllum (уруть), Elodea (водяная чума, неофит), Hippuris (водяная сосенка); 4— прикрепленные водные растения с плавающими листьями, Nymphaea (кувшинка), Nuphar(кубышка), Trapa (чилим, водяной орех, в настоящее время очень редок), Potamogetón (рдест); 5— камыши (корневищные растения): глубже всех Schoenoplectus (= Scirpus, камыш), далее по степени обмеления Phragmites (тростник), особенно обычный в эвтрофных водах Typha (рогоз) и в мягкой воде на тростниковых торфах Sparganium (ежеголовка); 6 — пояс крупных осок с крупнокочкарными видами Carex sp., в болотных водах Menyanthes (вахта, трифоль), Potentate palustris (сабельник) и с другими видами зоны зарастания, на краях болот Sphagnum (торфяной мох); 7, 8 — прибрежные деревья и болотистый лес, регулярно затопляемый, с Salix, Ainus и Populus; 9 — в лесу близ самой отметки уреза воды к элементам болотистого леса постепенно примешиваются элементы лиственного леса. Отображены также основные сферы биотопа, причем пе- досфера и биосфера перекрываются
Едва растительность смыкается, в качестве движущей силы сукцессии вступают усиливающиеся взаимодействия между видами. Определенные виды больше не могут обновлять свои популяции, поскольку начинают доминировать другие. Они остаются в сообществе только в качестве стареющих реликтовых популяций, в то время как следующая фаза сукцессии уже дает о себе знать появлением молодых растений подроста. Типичные примеры — береза и сосна в смешанных лесах. Они представляют собой в таких лесах чаще всего стареющие реликты более ранних стадий сукцессии (и таким образом являются показателями бывших нарушений) и как светолюбивые деревья не могут больше возобновляться при полной сомкнутости крон. Только в заключительной стадии развития растительности (климаксовая растительность) возникает относительно стабильное равновесие в видовом составе (см. рис. 14.34), а также между размножением и отмиранием участвующих в сообществе видов. Полностью стабильное состояние все же не достигается никогда. «Зрелые» растительные сообщества, которые никогда не были вовлечены в хозяйственный оборот и не испытывали катастрофических нарушений (сплошная рубка, использование под пастбища, пожары, затопление), всегда представляют собой мозаику разных сукцессионных стадий.
В бореальной зоне и верхнем поясе гор влажно-умеренного климата Северного полушария каждая автогенная не нарушаемая сукцессия происходит в направлении хвойного леса, на плоских равнинах — лиственного леса. Различают первичную сукцессию на свежих, только что освободившихся поверхностях (после отступления ледника, изменения русла реки) и вторичную сукцессию, например, на заброшенных залежах или после недавнего пожара. Соответственно, чему вошло в практику
различать леса первичные и вторичные, это не означает, что первичный лес никогда не нарушался естественным образом, но такие нарушения остались далеко позади. За очень долгое время (для лесов более сотни лет) сукцессионные ряды сходятся, и влияние макроклимата становится все более преобладающим, в то время как другие факторы отходят на второй план (зональная растительность). В Центральной Европе за очень небольшими исключениями существуют только вторичные леса различной степени «естественности» (обратно пропорционально степени окультуривания = гемеробии). В ландшафтах, никогда не использовавшихся человеком, ту стадию развития, на которой сукцессия приостанавливается, предрешает частота естественных нарушений (горные осыпи, лавинные полосы, зоны затопления, дюны, пожары, инвазия травоядных и др.).
Зональные типы растительности могут выступать так же, как и экстразональные, за пределами своей исконной области распространения, если локальный климат соответствует макроклимату этой основной области (например, произрастание субмеридиональных лесов из дуба пушистого на сухих южных склонах в Западной Европе).
Экосистема может меняться тем быстрее, чем меньше ее биомасса (В) и чем интенсивнее в ней потоки вещества и энергии. Если прирост биомассы на единицу времени обозначить как ДВ, то отсюда следует, что длительность полного оборота (turn-over) биомассы В/ДВ = 1. Сообщества планктона или терофитов могут соответственно этому изменяться за дни или месяцы, лесные же сообщества — за десятилетия и столетия.
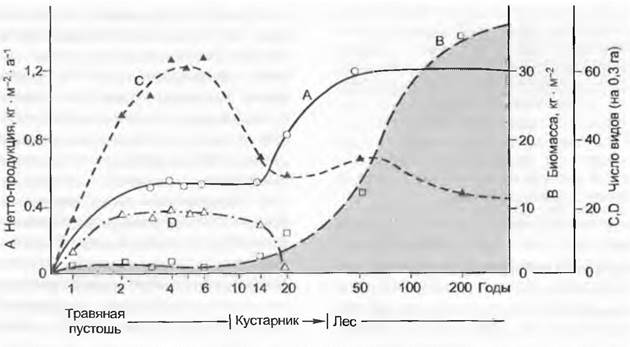
При формировании одного однородного растительного сообщества можно параллельно сменам стадий развития, зрелости и старения установить специфические изменения В, ∆В, дыхания (R) и продуктивности (Рn) насаждения (рис. 14.37). Эти изменения связаны с тем, что с прогрессирующим возрастом насаждения соотношения автотрофных компонентов (листьев) и гетеротрофных компонентов (стволов, ветвей и корней) все более уклоняются в сторону гетеротрофных и насаждение наконец угасает, если не происходит его омоложения.
Рис. 14.37. Фазы развития однородного лесного насаждения. Пропорции дыхания, опада (VA), прироста биомассы (∆В), нетто- (Рn) и брутто-продукция; отчуждение травоядными животными не принимается во внимание
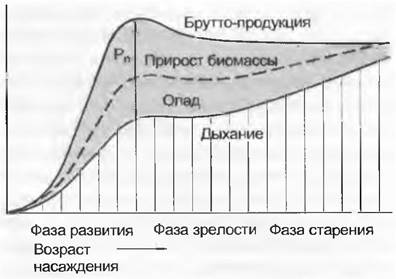
В ходе прогрессивной сукцессии комплексных биоценозов (см. рис. 14.33) также сначала происходит прирост В и Рn, так как ДВ больше, чем VA + VK. Такая экосистема продуктивна, но еще относительно изменчива и нестабильна. На стадии климакса наконец достигается фаза естественного омоложения, которая обеспечивает стабилизацию В на высоком уровне, поскольку прирост ДВ расходуется в ходе биологического круговорота. Отношение Рn и VА + Vк, так же, как и опада и разложения опавшего материала, уравнивается. Такая экосистема называется протективной. Как и климаксовая экосистема, она остается достаточно стабильной и имеет максимальную биомассу с очень небольшим нетто-приростом.
Рис. 14.38. Динамика растительности Центральной Европы под воздействием хозяйственного использования: регрессивные и (вторично) прогрессивные сукцессии на глубоких пологих известняковых склонах у подножия Высокого Венна (ФРГ). Растительные сообщества связаны в «кольца» соответствующих комплексов растительности и занимают определенную «плиту»
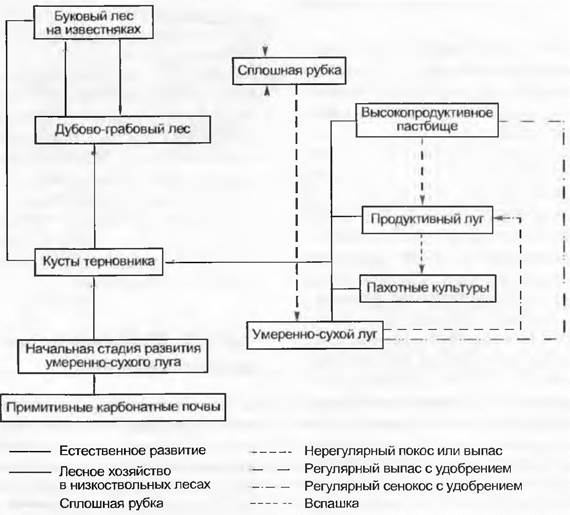
Регрессивные сукцессии идут в сторону от климаксовой растительности и связаны с ее деградацией. Если не принимать во внимание природных катастроф (землетрясения, ураганы и пр.) или катастрофических биологических изменений (например, голландская болезнь вязов, см. Ophiostomа), они почти всегда являются реакцией на вмешательство человека (см. рис. 13.51). Самыми тяжелыми являются те вмешательства, которые непосредственно или через ослабление растительности наносят ущерб почвам (перевыпас, регулярное выжигание).
Природные и антропогенные смены растительности на известковых почвах нижних горных поясов Центральной Европы обнаруживают как прогрессивные, так и регрессивные тренды, которые, будучи циклически связаны между собой (рис. 14.38), все же обратимы. Регрессионные смены, приводящие к ухудшению почв, такие, как перевыпас и повышенная частота пожаров на каменистых пустошах, ведущие к деградации средиземноморских дубовых лесов, в значительной степени необратимы.
14.3.3. Классификация типов растительности
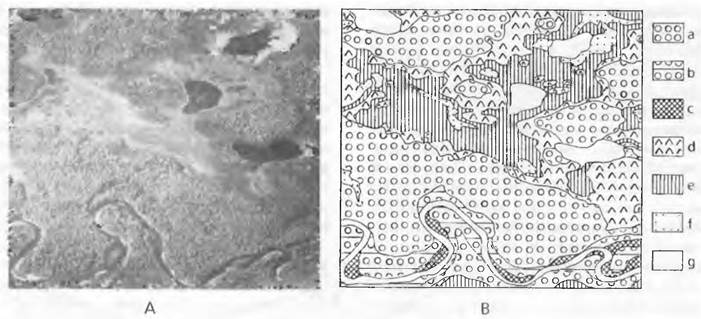
Хотя биоценозы, составляющие биосферу, и представляют собой континуум, все же между разными растительными сообществами возникают отчетливые границы, обусловленные пространственными различиями условий местообитания и различиями в сукцессионных стадиях. В рамках «систематики растительности» типизация конкретных растительных сообществ возможна и полезна. Статистические анализы показывают также, что держатся вместе только совершенно определенные комбинации видов (рис. 14.39). Эти характерные группы видов можно выделить как абстрактные типы растительности, реально отграниченные от других. На аэрофотоснимке лесного заболоченного ландшафта Аляски, еще совершенно не нарушенного человеком (рис. 14.40), даже глазомерно различаются разные единицы растительности, выделенные на соответствующей карте. В переходной зоне (экотон, англ. ecotone) набор видов меняется довольно очень быстро, а в пределах определенного типа растительности колеблется очень незначительно.
Рис. 14.39. Частота совместного произрастания 43 видов (кружки, буквенные символы) лугов в Нидерландах, представленная в виде диаграммы синэкологических корреляций (представьте объемно!). Разные виды часто растут вместе друг с другом (толстые линии связи) и характеризуют определенные растительные сообщества (и, соответственно, местообитания), например заболоченные луга с молинией = Molinia caerulea (М) и Carexpanicea (Ср), Potentilla erecta (Ре), Cirsium dissectum (Cs) и т. д.; болотистые приречныекамышовники с двукисточником = Phalaris arundinacea (Pha) и Glyceria maxima (Gm), Carex disticha (Cd), Caltha palustris (Cal) и т. д.; богатые сенокосные луга с райграсом = Arrhenatherum elatius (Arr) и Dactylis glomerata (D), Trisetum flavescens (Tn) и т. д.; интенсивно используемые пастбища с плевелом = Lolium perenne (Lp) и Cynosurus cristatus (Су), Роааnnuа (Ра), Trifolium pratense (Тр) и т. д.
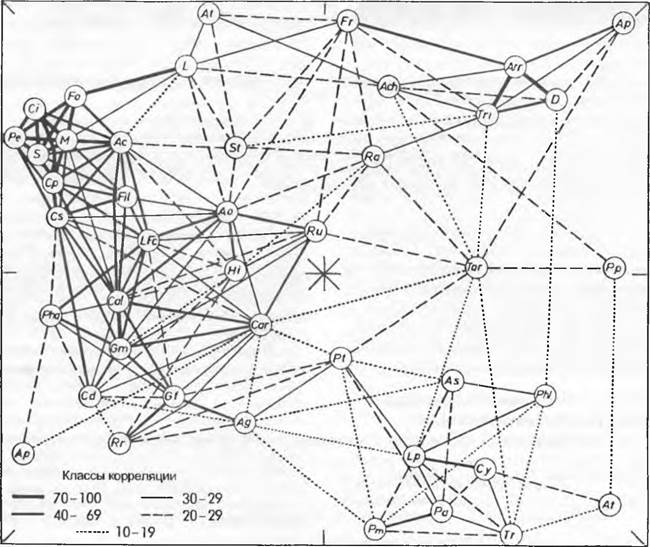
Рис. 14.40. Сопоставление аэрофотоснимка (А) и карты растительности (В) (низменность к северу от Анкориджа. Аляска; площадь 400 х 370 м): а — смешанный лес с березой (Betula resinífera и т. д.) за пределами болот и речной поймы; b — пойменный смешанный лес с бальзамическим тополем (Populus balsamifera) и березами; с — берега реки с ивняками (Salixsp.); d — заболоченные еловые леса (Picea mariana); е — моховые кустарничковые пустоши (Vaccinium uliginosum, Ledum decumbens) и Sphagnum, f— болотистые луга с осоковыми (Carex, Eriophorum и др.); g — водоемы и песчаные отмели
При флористической классификации растительности виды объединяются по степени их сходства в иерархически соподчиненные группы (совокупности видов). Эти группы близких друг к другу по отношению к условиям внешней среды видов и есть типичные для местообитания растительные сообщества. Обычно в таких сообществах имеются виды доминирующие или очень типичные (характерные или верные виды), которые могут быть использованы в названии сообществ. По таким принципам развивалась синтаксономическая система растительных сообществ Ж. Браун-Бланке, которая стала важнейшим инструментом геоботанического картирования и обобщения (табл. 14.4), особенно в Европе.
Таблица 14.4. Синтаксономическая система растительных сообществ
Категория |
Окончание |
Пример |
Класс Порядок Союз Ассоциация Субассоциация Вариант Фация |
-etea -etalia -ion -etum -etosum Без окончания Без окончания |
Molinio-Arrhenathefetea Arrhenatheretalia Arrhenatherion Arrhenatheretum Arrhenatheretum brizetosum Salvia-вариаш Фация c Bromus erectus |
Так, ассоциации союза райграсовых лугов входят в класс европейских сеяных лугов и в соответствии с субъединицами подразделяются на специальные фации; с Molinia(молиниевая). с Arrhenatherum (райграсовая), с Briza (трясунковая), с Salvia (шалфейная), с Bromus erectus (костровая).
В качестве условных синтаксономических единиц используются прежде всего ассоциации и союзы (например, Fagetum, Abietetum, Pinetum для соответственно буковых, пихтовых и сосновых лесов). В лингвистическом отношении для обозначения ассоциации применяется суффикс — etum (лат. коллективный суффикс), прибавляющийся к корневому названию доминирующего рода. Так, например, лиственнично-кедровый лес будет называться Larici-Pinetum, а сосняк с вереском — Erico- Pinetum.
Для синтаксономической характеристики определенного растительного сообщества используются:
• характерные (верные) виды — виды, основная область произрастания которых ограничивается какой-либо из главных таксономических категорий (характерный вид ассоциации, союза, порядка или класса) и особенно хорошо характеризует их с флористической точки зрения;
• дифференциальные (разделяющие) виды — виды, которые наиболее хорошо отделяют один синтаксон от другого, близкого к нему, но сами не ограничены этим синтаксоном, а могут произрастать и в других, «отдаленно» стоящих в системе классификации.
Характерные виды смешанного широколиственного леса (класс Querceo-Fagetea) — Daphne mezereum и Anemone nemorosa, для порядка Fagetalia — это, например, Ranunculusficaria и Mercurialis perennis, а для союза Fagion — Cardamine (= Dentaria) bulbifera и Hordelymus europaeus.
Высокая степень совпадения местообитаний определенных групп видов (см. рис. 14.39) позволяет дать экологическую характеристику этих местообитаний, так что виды часто могут служить показателями условий среды. Редко бывает, когда в распоряжении имеется экофизиологическая характеристика видов, основанная на данных измерений. Тем не менее, учитывая имеющийся опыт многих поколений полевых ботаников, можно охарактеризовать определенные свойства каждого вида. Такой полуколичественный метод представляют таблицы индикаторных значений (система Г. Элленберга или Е. Ландольта). В них индексы от (0)1 до 9(10) присваиваются видам в зависимости от свойственных им отношений к определенным, имеющимся в наличии ресурсам среды, значения которых меняются по возрастающей. Высокий показатель по увлажнению означает, что произрастание этого вида коррелирует с высокими значениями влажности. Типичные индикаторные растения для сильнокислых почв (R от 1 до 2, см. ниже) — это, например, Deschampsia /flexuosа, Vacciniummyrtillus. Причинная связь, указывающая на потребности вида, этим индексом не выражается, но во многих случаях она может подразумеваться. Подобные индикаторные значения действительны не для отдельных изолированных особей, а для растений, которые произрастают в сообществе и участвуют в биотических взаимодействиях. Дополнительная трудность состоит в том, что этот метод из чисто практических соображений привязан к уровню вида в целом, но экотипы одного вида могут различаться между собой по потребностям даже сильнее, чем два различных вида. Несмотря на это, такие оценочные методы имеют большое практическое значение прежде всего благодаря своей простоте и, что довольно удивительно, точности (табл. 14.5).
Таблица 14.5. Индексы для условий Центральной Европы, по Элленбергу
Условия внешней среды (величины) |
Символ |
Индекс (что показывает вид) |
Свет |
I |
1 — глубокая тень, 5 - полутень, 9 — полная освещенность |
Температура |
Т |
1 — альпийско-субнивальный климат, 5 —субмонтанно-умеренный, 9 — средиземноморский |
Континентальность |
К |
1 — наиболее океанический, 5 — промежуточный, 9 — наиболее континентальный |
Влажность |
F |
1 — почва очень сухая, 5 — свежая, 9 — сырая, 10 — вода |
Реакция почвы (pH) |
R |
1 — сильнокислые почвы, 5 — умеренно кислые, 9 — основные (известняковые) |
Содержание азота |
N |
1 — минимальное. 5 — умеренное, 9 — чрезмерное |
Засоленность |
S |
0 — отсутствует, 1 — слабое, 5 — умеренное, 9 — экстремальное засоление |
Виды растений с похожими комбинациями кодовых цифр образуют экологическую группу. Средние значения этих цифр позволяют оценить все растительное сообщество, причем индифферентные (не связанные с определенным сообществом) виды заранее исключаются, а в расчет принимаются только типичные виды,
«взвешенные» по значениям их покрытия. Перечень расширяется путем включения любых других значимых индексов, что дает возможность проводить разнообразный анализ в системе базы данных. Так, можно добавить сведения о морфологии (жизненной форме), способах распространения, времени цветения, чувствительности к нарушениям (например, рудеральный вид или не рудеральный), гемеробии («зависимости от культуры», в смысле указания на условия, не измененные антропогенными воздействиями), времени внедрения во флору (автохтонный вид, неофит), а также ботанико-географические данные (область распространения).
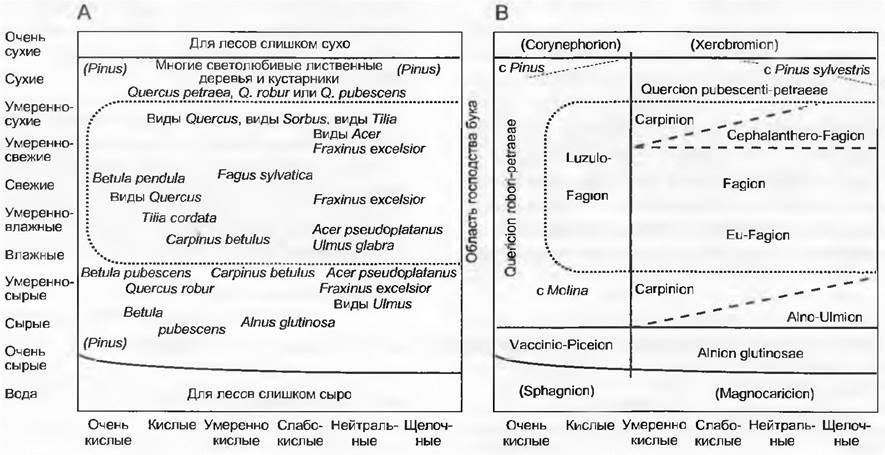
Единицы растительности, более высокие по рангу, чем союз, слишком абстрактны и поэтому в меньшей степени подходят в качестве примера конкретных растительных сообществ. На рис. 14.41 лесные растительные сообщества Центральной Европы представлены в системе координат по отношению к влажности и реакции почвы, причем слева приведены отдельные характерные виды, которые можно сравнить с представленной справа системой синтаксонов.
Для всех комплексов растительных сообществ. т. е. объединений нескольких сообществ, классификация растительности по аналогичной номенклатуре разрабатывается сигма-социологией (симфитосоциологией), что применяется прежде всего в немецкоязычных странах (сигма Σ — знак суммы). Так, как только определенные виды объединяются друг с другом в сообщества естественным образом, приходится разные сообщества объединять вместе (например, прогалина в лесу, лес, берег ручья в лесу образуют вместе комплекс, составляющий одно целое, «сигмету»). По аналогии с синтаксономией сигма-синтаксономия использует понятия характерных и дифференциальных для сигметы сообществ и применяет окончания, соответствующие системе Ж. Браун-Бланке.
Рис. 14.41. Лесообразующие виды деревьев Центральной Европы: А — характерные виды для почв от кислых до щелочных (соответственно, бедных и богатых питательными веществами) и от влажных до сухих (в субмонтанном горном поясе и в условиях умеренно субокеанического климата). Приблизительные количественные соотношения в объединениях сообществ, близких к естественным, набраны курсивом (названия в скобках только в некоторых областях); В — фитосоциологическая система сообществ лиственных лесов Центральной Европы на почвах от кислых до щелочных (соответственно, бедных и богатых питательными веществами) и от сырых до сухих (в субмонтанном горном поясе и в условиях умеренно субокеанического климата). В краевой зоне располагаются еловые (Vaccinio-Рiceion) и ольховые (Аlnion glutinosae) леса; нелесные типы растительности — ксерофильные злаковые луга на бедных и богатых известью породах (Сorynephorion и Хеrobromion соответственно), а также верховые болота (Sphagnion) и сообщества зарастающих берегов водоемов (Маgnocaricion). Линии границ соответствуют в природе зонам с переходными типами объединений сообществ
14.3.4. Физиономическая классификация растительного покрова
Насаждения независимо от флористического состава можно группировать по преобладающим жизненным формам и физиономическим признакам (габитусу, внешнему облику). Такие характеризующиеся как самостоятельные, но в то же время в сущности комплексные типы растительности называют формациями; как пример можно привести влажно-тропические леса, бореальные хвойные леса, вечнозеленые жестколистные кустарниковые заросли, кустарничковые пустоши, луга и т. д. Напротив, простые, составленные только одной жизненной формой и часто не существующие самостоятельно объединения растений обозначают как синузии, например, это корка накипных лишайников на скалах, кустарничковый ярус в хвойном лесу или совокупность шляпочных грибов в осеннем лиственном лесу. Растительные формации вместе с окружающей средой, в которой они существуют, образуют биологические формации или сокращенно — биомы. Им присваивается название доминирующей растительной формации, поэтому биомы и формации по наименованию — синонимы.
С синтаксономическим делением растительности имеются несомненные перекрытия. Несмотря на различия в видовом составе, внутри сходных биомов часто произрастают одни и те же роды, например, в летне-зеленых лиственных лесах холодно-зимних умеренных областей Северной Америки и Евразии это роды Quercus, Fagus, Capñnus, Acer, Tilia и др. И напротив, отмечено конвергентное физиономическое сходство у тех формаций, которые образованы представителями разных семейств, что вызвано сходными условиями внешней среды. Примеры этого— заросли жестколистных кустарников средиземноморского климата, расположенные вокруг всего земного шара, или ряд суккулентных полупустынь Старого и Нового Света (Euphorbiaceae — Cactaceae). Некоторые формации почти не связаны с климатом того места, где они расположены, например, злаковники, которые произрастают как в континентальных степных областях умеренной зоны, так и в тропиках.
Попытка разделить растительный покров всей наземной поверхности биосферы Земли на типы формаций (имея в виду только поздние стадии сукцессий) представлена на рис. 14.42. В качестве координат служат среднегодовые суммы осадков и среднегодовые температуры, причем последние, как видно на климадиаграммах, отражают потенциальное испарение. Отдельные формации подробнее описаны в гл. 15.
Рис. 14.42. Формации континента: попытка классификации кпимаксовой растительности по средним значениям годовой температуры и количества осадков. Условные знаки означают характерные жизненные формы. Линии границ нечеткие, особенно в центральной части диаграммы, так как относительное положение формаций по отношению к климатическим данным в разных областях может довольно значительно сдвигаться

14.3.5. Пространственная структура растительности и биотопов
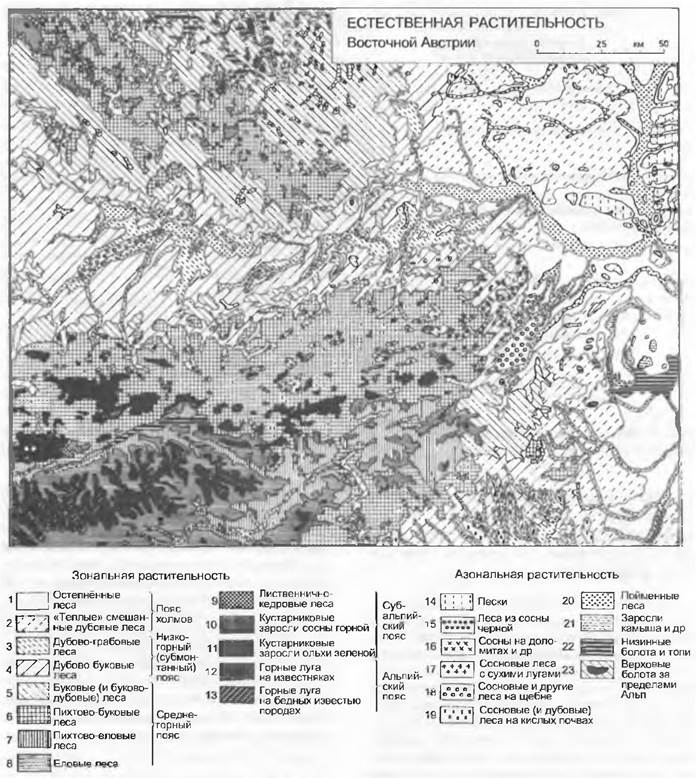
В пределах определенного регионального климата выявляется обусловленное рельефом и эдафическими условиями горизонтальное расчленение естественной растительности (или ее фрагментов), которая сочетается с растительностью, находящейся под воздействием человека (культурный ландшафт). Часто фактическая (антропогенная) растительностьочень отличается от потенциальной, естественной (например, пастбище на месте леса). Изображение этого пространственного расчленения растительного покрова содержат карты растительности. Однако они могут отображать растительные сообщества ранга ассоциации только в очень крупном масштабе. На региональных или тем более межрегиональных картах могут быть представлены только обширные комплексы растительности, что иллюстрируется картой Нижней Австрии на рис. 14.43. Карты такого или еще более мелкого масштаба представляют, как правило, лишь потенциальную растительность, т. е. ту растительность, которая могла бы длительно существовать, если бы не было воздействия человека. В континентальном или глобальном масштабах пространственная структура растительности соответствует крупным климатическим зонам, обусловленным снижением температуры от экватора к полюсу, и наложенному на них градиенту океаничности-континентальности. В таком масштабе отображаются только климаксовые формации потенциальной растительности.
Рис. 14.43. Деление растительного покрова Центральной Европы на примере Нижней Австрии. Потенциальная естественная климаксовая растительность (почти повсюду леса!), соответствующие ей высотные пояса и большие пространства длительно производных растительных сообществ. Флористические районы (подрайоны) среди зональных типов растительности: 1 — 2 — паннонский, 3 — 8 — центрально-европейский, 9 — 13 — альпийский. Вдоль крупных рек (Дуная, Марх и др.) представлены пойменные леса (20)
При выделении высотных поясов растительности в горах мы имеем дело также с комплексами растительности, которые в каждом случае относят к доминирующей потенциальной климаксовой формации или доминирующему растительному сообществу (рис. 14.44). Последовательность этих комплексов растительности отражает изменяющийся с увеличением высоты («высотный») климат. Единственный общий для всех зон климатический признак высотного градиента заключается в падении температуры. Все другие климатические факторы изменяются в зависимости от географической широты (укорочение с высотой вегетационного периода — это явление только удаленных от экватора широт) или с региональными особенностями макроклимата (в некоторых областях Земли облачность, осадки или ветер с высотой увеличиваются, а в других — снижаются). Наблюдаются также сильные различия между краевыми зонами горных систем и их внутренними, чаще всего более континентальными участками, как и между наветренными и подветренными по отношению к основному направлению воздушных масс горными цепями. Принятые в международной практике обозначения горных поясов и соответствующей им последовательности растительности в Альпах даны на рис. 14.44. Существуют также средне- и низкогорные, предгорный (холмистый) и равнинный пояса.
Рис. 14.44. Верхние высотные пояса Альп с характерными мозаично сложенными комплексами формаций
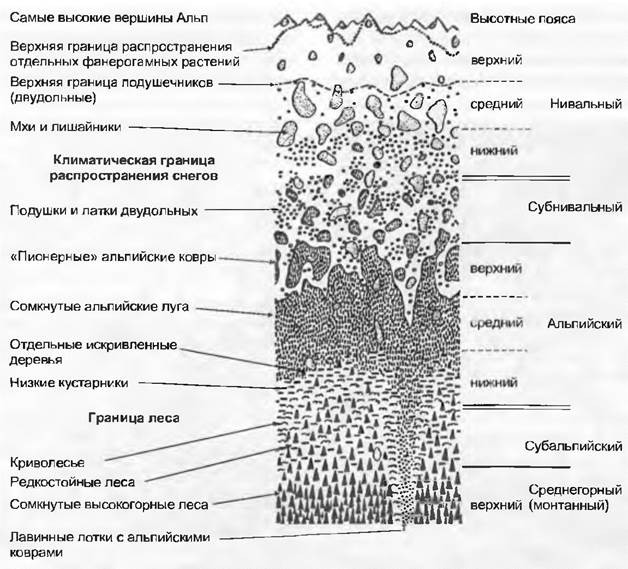
К сожалению, употребление этой номенклатуры высотных поясов в литературе не всегда единообразно. Это касается в основном понятия «субальпийский», которое используется в очень широкой интерпретации. Такая трактовка, которая дана на рис. 14.44, применяется во всем мире наиболее широко и охватывает зону, где горные редколесья верхнего пояса сочетаются с альпийской растительностью (часто ее называют экотоном верхней границы леса, англ. treeline ecotone). Региональные «школы» тем не менее с точки зрения фитосоциологических критериев используют это понятие также для обозначения пояса горных лесов (верхний горный пояс). Понятия «нивальный» и «субнивальный» мало употреби мы за пределами Европы. Понятие «альпийский» относится не только к Альпам (корень alb, alp, или alpo присутствует в словах, обозначающих горы или их склоны в древнеримском языке, возможно, в древнеиндо-германском; см. Ch. Körner, 1999), но и используется во всем мире для обозначения безлесной растительности, расположенной выше естественной климатически обусловленной границы леса (даже когда ее местами уже нет по причине антропогенного воздействия или естественных нарушений). Обиходное употребление понятия «альпийский» в смысле просто «горный» («альпииский ландшафт», «альпийский экономический район», «альпийская культура») не имеет ничего общего с ботанико-географическим определением. В Андах для альпийского пояса употребляется понятие «андский», в Африке — «афро-альпийский».
14.3.6. Корреляционный анализ растительности
На основании инвентаризации растительности и ее классификации, описанной в разделе 14.3.3, при наличии данных по характеру местообитаний и индикаторных
видов можно провести серию ординационных и корреляционных анализов, конечная цель которых — выявить повторяющийся характер в распространении видов и растительных сообществ и объяснить его исходя из конкретных внешних условий.
Градиентный анализ ориентирован на изменение растительности под воздействием меняющихся условий среды. В идеальном случае, но это бывает далеко не всегда, на изученных площадях реализуется один сопряженный градиент. Если упорядочивать, например, обилие видов по градиентам окружающей среды, они сортируются по значениям увеличения или снижения значений градиентов, и этот процесс называется прямой ординацией. Если выбрать только одну переменную, то это — одномерная ординация (наглядный пример: профиль растительности на отрезке от берега моря соответственно степени засоления почвы, см. на рис. 13.28). При двухмерном изображении говорят уже об экодиаграмме (см. рис. 14.42), причем на двух осях откладываются значения двух параметров внешней среды и месту произрастания вида растений иди сообщества придается определенное значение в системе координат X — Y, которое обозначает положение вида или сообщества в двухмерном пространстве (рис. 14.45, 14. 46). При этом возникают узоры (например, скоплений, корреляций), которые можно интерпретировать как предпочтительные условия местообитаний определенных ассоциаций. Поскольку, как правило, воздействие оказывают сразу много факторов среды, поэтому не ясно, какой из них или какая их комбинация являются критическими, ординационное пространство при подробном анализе всегда многомерно и охватить его можно только с применением компьютерных методов. В связи с этим говорят о корреляционном (а также математическом, численном, статистическом, мультивариантном или количественном) анализе растительности.
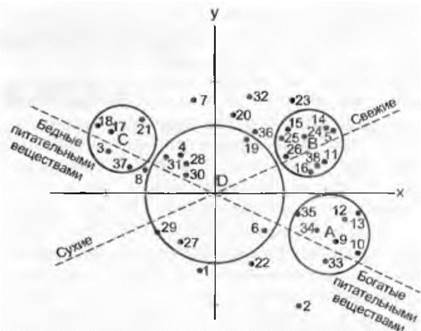
Рис. 14.45. Виды буковых лесов (1 — 38), упорядоченные по их совместному произрастанию в ассоциациях Fagetum во Франции с помощью оптимального двухмерного факторного анализа. Градиенты — влажность и богатство почвы. Хорошо выявляются группы индикаторных видов для богатых питательными веществами (А), свежих (В), бедных питательными веществами (С) и типичных (D) образований
Большое число комплексных методов и расчетных программ разработало специально для параметров растительности в сочетании с данными по местообитаниям (например, корреспондентный анализ СА, канонический корреляционный анализ ССА и как вариант— анализ главных компонент РСА —principal component analysis). Он, как правило, проводится в два этапа. Сначала «отыскивается» и определяется тип растительности, затем для него, точнее для отдельных описаний растительности, определяются значения параметров внешней среды. Оба этапа основаны на анализе корреляций. Благодаря этому становится очевидным, какие факторы внешней среды с большей вероятностью определяют связи растительности.
Рис. 14.46. Растительные сообщества Польши, упорядоченные по их флористическому сходству с помощью оптимальной двухмерной полярной ординации: верховое болото (Sphagnetum medii = Sm) облесенное болото (Pineto-Vaccinietum uliginosi = PVu), сосновый лес (Pineto-Vaccinietum myrtilli = PVm), пихтовый лес (Abietetum polonicum = Ар), заболоченный ольшаник (Cariceto elongatae- Alnetum = CeA), ясенево-ольховый пойменный лес (Circaeo-AInetum = СА), дубово-грабовый лес {Querceto-Caprinetum medioeuropaeum = QC), буковый лес (Fagetum carpaticum = Fc), смешанный дубовый лес (Querceto-Potentilletum albae - QP), заросли лещины (Coryleto-Peucedanetum cervariae = CP). В скобках после каждого сообщества — среднее число видов сосудистых растений
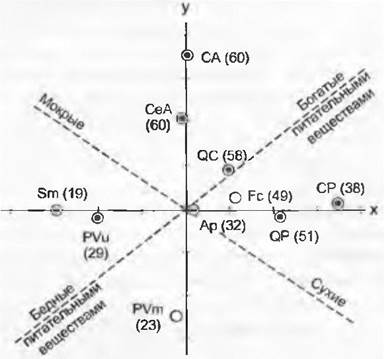
Каждое из растительных сообществ исследуемой местности становится при этом «объектом» («компонентом»), который характеризуется своим видовым набором (= «атрибутом»). То, что вид произрастает в определенном видовом сообществе, является для него специфическим признаком, так же как господствующие условия внешней среды — признак этого вида, определяющийся на втором этапе. В таких корреляционных методах используется, как правило, не простая констатация наличия вида по принципу «да или нет», но вид регистрируется с учетом его обилия (см. 14.3.1). Поэтому для каждого вида имеется условная ось обилий, начинающаяся от нуля и заканчивающаяся 5 (100% покрытия). Описание растительности соотносится с этой осью в зависимости оттого, насколько вид в нем обилен. Поскольку видов (и шкал обилия) много, возникает многомерное (корреляционное) пространство, в котором отдельные описания растительности подчинены одновременно многим осям отдельных видов, что почти невозможно себе мысленно представить, но что приемлемо для математической обработки. Каждое описание растительности (каждый объект) соответствует точке в этой комплексной многомерной координатной системе. Если провести обработку 100 описаний, то и точек будет 100. С помощью математических методов эту многомерную систему можно свести к нескольким осям и затем представить облако точек (описаний) в двух- или трехмерном пространстве. Многомерная система при этом свернется таким образом, что большая часть флористического разнообразия отразится на немногих осях. Совокупности видов (описания растительности) упорядочатся по отношению друг к другу при этом в некую диаграмму соответственно их флористическому сходству (расстоянию на диаграмме). Такая диаграмма отражает совокупности групп описаний (объектов), так называемые кластеры. Они соответствуют типам сообществ, а метод отвечает требованиям неиерархической классификации. Облака точек кластера распознаются с первого взгляда как резко отграниченные друг от друга группы (= типы растительности) или же переходят друг в друга более или менее постепенно.
Если вместо номеров описаний растительности проставить в корреляционном пространстве соответствующие значения pH, типов почвы, влажности, положения в рельефе и т. д., можно таким же образом установить корреляцию между обилием видов и условиями местообитания. По изменениям условий местообитания между разными описаниями отображают направления градиентов среды (например, слева на оси будут более низкие значения pH, справа — более высокие). Вращая координатную систему, также находят такое положение, при котором одна из осей достигает максимальной дисперсии «нагрузки» («первая» ось имеет наибольшее значение для дальнейшего толкования). Если определенный градиент лежит параллельно первой оси, то из этого можно заключить, что данный экологический фактор оказывает наиболее сильное влияние на видовой состав. Вторая ось. расположенная под прямым углом к первой, показывает наибольшую оставшуюся дисперсию, т. е. следующий по важности фактор, и т. д. С помощью этого «непрямого градиентного анализа» и с применением приспособленного для этих задач компьютера обрабатывают очень большой массив данных.
Компонентный анализ также представляет собой процесс сокращения числа пространственных измерений путем перевода многомерных облаков точек в двух- или трехмерное пространство. Полученные оси являются результатом анализа, а не установлены априори, как при одно- или двухмерной (графической) ординации. Таким способом можно выяснить, при каких условиях среды два вида могут произрастать вместе, имея высокое обилие (т. е. установить их общую преференцию местообитания). Это можно сделать с применением компьютерных программ путем сравнительного анализа всего видового состава, из чего рассчитывается и строится матрица связей (сеть корреляционных плеяд, растровая диаграмма, см. рис. 14.39). Такой анализ сходства используется при цифровой классификации близкородственных растительных сообществ и выражается потом в виде дендрограммы (кластерный анализ).