БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
15. РАСТИТЕЛЬНОСТЬ ЗЕМНОГО ШАРА
Растительный покров Земли является зеркальным отражением климата, накладывающимся на региональные геологические условия и существующие нарушения среды (рис. 15.1). Поскольку климатические условия не имеют резких географических границ, нет и резких границ между растительными зонами, которые определяются доминированием определенных жизненных форм (так называемых формаций, см. рис. 14 42, раздел 14.3.4). В этой главе рассматривается естественная зональная растительность, т. е. тот растительный покров, который развился под воздействием определенных климатических условий без учета воздействия человека, причем с учетом тесно связанных друг с другом факторов субстрата и всевозможных нарушений (исключение — см. раздел 15.2.16). Но это идеальная картина, так как влияние человека на растительность (охота, пастбищное скотоводство, пожары) существует с древнейших времен, и результат этого длительного воздействия не всегда можно распознать. Рассмотрение всех моментов антропогенного воздействия, от лесных кочевий до земледелия сильно расширило бы рамки настоящей главы. По таким же причинам в ней будут рассмотрены в основном поздние стадии сукцессии, так называемые завершающие растительные сообщества, или климаксовая растительность (см. 14.3.2).
Рис. 15.1. Факторы, влияющие на распределение зональной растительности Земли. Для обеспечения наглядности на рисунке представлены не все взаимодействия
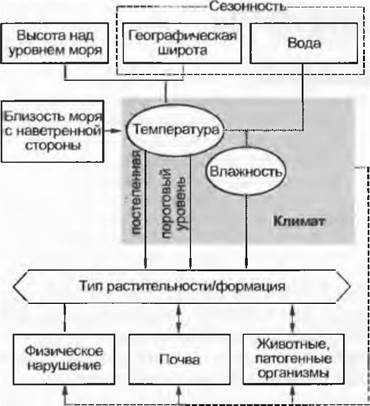
Кроме таких неизменных условий, как географическая широта и высота над уровнем моря, которые обусловливают температурный режим, значительную роль играют также циркуляция атмосферы и морские течения, осуществляющие водный перенос. Обеспечение влагой зависит не только от абсолютного количества атмосферных осадков, но и от соотношения выпавших осадков и испарения их в атмосферу, причем последнее опять-таки зависит от температурных условий. Географическая широта воздействует (астрономически обусловливает) на определенные сезонные смены температурного режима, которые, со своей стороны, прямо или опосредованно определяют сезонный характер водного режима. В высоких широтах доминирующим фактором является сезонная смена температур, в более низких — водного режима. В отношении воздействия температуры важны как ее постепенные изменения, так и пороговые уровни (например, мороз — или он есть, или его нет). В холодных, но все-таки безморозных областях (например, на некоторых побережьях морей умеренной зоны) могут существовать тропические виды, а в сравнительно более теплых, но континентальных и вследствие этого с возможными морозами областях — нет. Большую роль играет также близость моря. Значительно воздействуют на растительность такие типичные для некоторых зон нарушения, как тропические циклоны или пожары, животные — крупные млекопитающие, как стравливающие травяной покров (англ. grazer), так и поедающие листву и ветки кустарников и деревьев (англ, browser), а также и зональные почвенные процессы, которые происходят под влиянием многих биотических и абиотических факторов. Несмотря на многообразие действующих факторов окружающей среды (см. рис. 15.1) современную растительность земного шара (см. форзац) можно «спрогнозировать» на основе очень немногих параметров (в основном, только температуры и осадков) с помощью математических моделей (см., например, модели А. Вудварда или Р.Лссманса). Это говорит о преобладающем влиянии климата на растительность (см. рис. 14.42) и открывает возможности для проектов прогноза на будущее.
Поскольку зоны растительности соответствуют климатическим зонам, чаще всего их описывают, исходя из представлений климатологии. Г. Вальтер дал определение так называемых зональных биомов, свойственных крупным климатическим областям Земли (см. 15.2). У них разное пространственное размещение, в одних случаях их границы четко обозначены (например, зона бореальных хвойных лесов), а в других они представляют собой некое сборное понятие, объединяя похожие, но при ближайшем рассмотрении очень различные типы растительности. Эта классификация, которая будет представлена ниже, является таким образом практичным, но, в силу обстоятельств, довольно грубым вспомогательным средством. Первая часть этой главы — краткий обзор растительности Центральной Европы, это та растительность, с которой читатели непосредственно знакомы. Во второй части описывается растительность земного шара, сгруппированная в 9 зональных биомов, причем в пределах некоторых зональных биомов выделены подгруппы по высоте над уровнем моря (горные, или «оробиомы») или по степени аридности.
15.1. Растительность Центральной Европы
Центрально-европейскую флористическую область можно определить, как гумидную, прохладно-умеренную, с зимними холодами, в ботанико-географическом отношении она простирается с запада на восток от Ирландии и северо-западной Испании, постепенно сужаясь, до Урала (см. рис. 14.19). На севере охватывает южную часть Скандинавского полуострова. По Альпам и Карпатам проходит граница с другой флористической областью на юге (климат, см. 15.2.9).
15.1.1. От низменных равнин до горных лесов нижнего пояса
Климаксовая формация гумидной, прохладно-умеренной зоны — летне-зеленые лиственные леса (см. рис. 14.42, 15.2.9). В течение вегетационного периода, длящегося 5 — 6 месяцев (с конца апреля до начала октября), продуктивность этих лиственных лесов почти так же велика, как и во влажных тропических лесах (но если пересчитать на весь год, то вдвое меньше). Эти леса характеризуются синхронным распусканием и сбрасыванием листьев, устойчивыми к зимним холодам почками, и сопутствующей флорой, находящейся под воздействием сезонных колебаний светового режима под пологом леса. Хвойные деревья принимают участие или даже преобладают в составе древостоя только в условиях континентального климата (особенно на северо-востоке) и в более холодных горных лесах (см. 15.1.2),
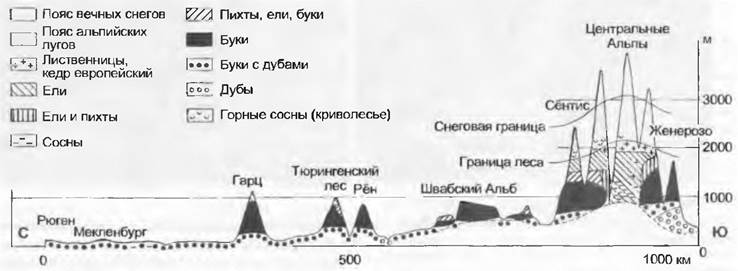
До начала эпохи земледелия Центральная Европа была полностью облесена (см. рис. 11.308). Только в Альпах, Судетах и Карпатах у лесов существовала верхняя климатическая (обусловленная температурными условиями) граница; на самых высоких вершинах Шварцвальда, Бемервальда и Гарца можно было увидеть пояс «борьбы за существование», который был в большей степени обусловлен положением на открытых хребтах, чем высотой над уровнем моря (рис. 15.2). Климатические ограничения роста лесов в наиболее сухих и теплых ландшафтах, обусловленные засушливостью, также отсутствовали, а препятствовавшие росту лесов морские ветры ограничивали только облесение островов и очень узкой полосы побережья Северного и Балтийского морей. Не занятыми лесами по естественным причинам оставались только такие места, где для роста деревьев была слишком маломощная (и поэтому очень сухая), или слишком переувлажненная, или засоленная почва. Но девственные центрально-европейские леса не представляли собой сплошной покров не только в силу этих причин. Мозаично расположенные среди них открытые поверхности, на которых происходило становление современной луговой флоры, возникали вследствие естественных циклов развития лесов, стихийных повреждений насекомыми, естественных лесных пожаров, ветровалов, на них паслись крупные травоядные млекопитающие, например, зубры. Несмотря на это, наибольшая часть воздействий на леса все же приписывается деятельности человека, которая привела к тому, что в настоящее время облесена лишь четвертая часть этого крупного региона (и во многих случаях только в форме искусственных посадок).
Рис. 15.2. Высотные пояса растительности Центральной Европы на профиле с севера на юг. Высотные пояса поднимаются по направлению к югу и по мере увеличения общего поднятия рельефа В континентальных внутренних Альпах бук исчезает, доминирует ель
С флористической точки зрения центрально-европейский регион характеризуют прежде всего основные доминанты листопадных лесов, это Querem robur, Fagus sylvatica, Acerplatanoides, Fraxinus excelsior, Coryfus avellana, Anemone nemorosa и многие другие. Тесные исторически обусловленные флористические связи с другими частями пояса лиственных лесов Голарктики проявляются в том, что те же самые или близкородственные виды произрастают в китайско-японской и атлантическо-североамериканской флористических областях (например, Fagus, см. рис. 14.17). Флора центрально-европейской области была очень сильно обеднена вследствие четвертичных оледенений, большая часть произрастающих здесь в настоящее время видов смогла возвратиться сюда в позднеледниковое время или во время раннего послеледниковья из южных (в основном, более восточных) рефугиумов (см. 11.3). Это является одной из важных причин, обусловливающих тесные флористические связи со Средиземноморской (субсредиземноморской) областью (см., например, рис. 14.21).
Флористическую дифференциацию центрально-европейской области можно распознать именно по связям ее видов — океаническим или континентальным, бореальным или субмеридиональным. Это дает основание к расчленению ее на атлантическую, субатлантическую, центрально-европейскую и сарматскую провинции. На более высоком уровне Альпы и Карпаты на основании особенностей их флоры и растительности выделяются в качестве самостоятельных подобластей (альпийской, карпатской), главным образом за счет верхних горных поясов растительности (см. рис. 14.19, 15.1.2).
Характерные виды атлантической провинции, например, Ulex europaeus, Myricagale, Erica tetralix, E. cinerea, Helleborus foetid us, IIex aquifolium (см. рис. 14.25), большой частью ареала заходят в Средиземноморскую область и могут поэтому называться атлантическо-средиземноморскими.
Для субатлантической провинции свойственны виды, ареал которых простирается далее на восток, например, Cytisus scoparius, Lonicera periclymenum или Digitalis purpurea. В раст ительном покрове атлантической и субатлантической провинций, характеризующихся более мягкими зимами, почти полностью отсутствуют хвойные. Здесь, помимо лиственных лесов, составленных преимущественно дубом и березами, болот и лугов на бедных подзолистых почвах, большую роль в растительном покрове играют сообщества кустарничковых пустошей из вересков — Calluna и атлантических видов рода Erica.
Центрально-европейская провинция характеризуется прежде всего видами, распространенными в основном на континентальном востоке, но ареал которых частично ограничен океанической западной частью Европы: Abies alba, Fagus sylvatica, Caprinus betulus, Quercus petraea, Tilia platyphyllos, Galium syivaticum и др.
Сарматская провинция охватывает восточную часть центрально-европейской области. Среди основных видов здесь можно назвать характерные спутники дубовых лесов: Euonymus verrucosa, Potentilla alba, Melampyrum nemorosum. Преобладают смешанные сосново-дубовые леса из Quercus robur и Pinus sylvestris, бук отсутствует. На юго-востоке они частично переходят в лесостепь.
К центральноевропейским лиственным и хвойным лесам нижнего пояса причисляются:
(1) Буковые леса и смешанные леса со значительным участием бука (см. рис. 15.5, D, E, F) с ясенем, горным кленом, липой, на юге также частично с пихтой и др. Эти леса преобладают в западных среднегорьях, а кроме этого в более низких поясах всех средневысотных гор и в известняковых массивах Альп. На равнинах они встречаются особенно часто в области молодых конечных морен, на почвах, богатых питательными веществами (рис. 11,259).
(2) Дубово грабовые смешанные леса приурочены к тем участкам нижнего пояса гор на более богатых почвах, где бук, который они обычно вытесняют, находится на границе своего распространения или вблизи нее (к примеру, в северо-западной Германии и в сухих внутренних ландшафтах).
(3) Теплолюбивые смешанные дубовошироколиственные леса покрывают часто склоны южных экспозиций и сухие пологие горные склоны. В них произрастают такие субсредиземноморские виды, как Quercus pubescens (дуб пушистый), Acer monspessulanum, Cornus mas и многие травянистые растения, господствующие южнее или восточнее.
(4) На бедных питательными веществами кислых почвах нижних уровней процветают дубовые леса с Calluna vulgaris (вереском) и другими неприхотливыми растениями в нижнем ярусе.
Таблица 15.1. А — D —леса гумидных тропиков (А — С — тихоокеанское побережье Панамы; D — Папуа-Новая Гвинея) с характерными окнами, где идет возобновление; Е, F — досковидные корни стабилизируют, а лианы разрушают; G — эпифиллы скрывают жилкование листьев; Н — большой ущерб наносят травоядные (в данном случае листоядные муравьи)

Таблица 15.2. А — горный лес из Gynoxis, Пасо дела-Вирген, Эквадор, 4000м; В — реликтовый лес из Polylepis sericea, Мерида, Венесуэла, 4050 м; С — богатый видами облачный лес Ранчо Гранде, север Венесуэлы, 1 100м; D — туманный лес, г. Каинди, Папуа-Новая Гвинея (граница туманов, 1 800 м); Е — туманный лес из Guercus, Лас Нубес, Панама, 2 200 м, F — горный дождевой лес близ Пасо дела-Вирген, Эквадор, 1 900 м; G — Tillandsia usneoides в туманном лесу гор Мерида, Венесуэла, 1 800 м; Н — дерево с эпифитами в горном дождевом лесу Пасочоа близ Кито, Эквадор, 2800 м

Таблица 15.3. А, В — кроновая розетка Espletia, Парамо-эль-Анжель, северный Эквадор, 3 600 м; С — растения — шерстяные свечи, здесь — Lupinus allopecuroides, Гуагуа Пичинча, Кито, Эквадор, 4 300 м; D — Tnchocereuspasacana, перевал Кумбре, северо-западная Аргентина, 3 050 м; Е — Gentiana nevadensis близ пика Боливар, Венесуэла, 4150м; F —Парамос с Espletia timotensis и Hypericum ericoides, Пасо Агуила, Венесуэла, 3 900 м; G — плотная подушка Azorella compacts (Apiaceae); H — клональные гирлянды Festucaorthophylla, перевал Кумбре, северо-западная Аргентина, 3900/4 250 м

Таблица 15.4. Один и тот же лес в Панаме в сезон дождей (А) и в засушливый период (В); С — сезонно-зеленый тропический лес в северной части Венесуэлы, массовое цветение Tabebuja (Bignoniaceae); D — лесмиомбо в западной части Замбии с Brachystegia scipiformis; Е, F — сухой лес из Гуанака, Пуэрто-Рико (200 дней без дождей): с Bursera (Е), эпифитами (Tillandsia, G), суккулентными лианами (Vanille, G) и кактусами(Н)

Таблица 15.5. А — D — различные «открытые» формы саванн в национальном парке Крюгер, Южная Африка; В — река с галерейным лесом; Е — плоды Combertum heteroense, F — Acacia tortilis; G -северо-австралийская саванна с термитниками; Н — мульга-скраб, северная Австралия

Таблица 15.6. А, В — соответственно Larrea tridentata и Camegia gigantea в пустыне Сонора, Аризона; C — Jatropha (Euphorbiaceae) и Opuntia, северо-западная Аргентина; D — бутылочное дерево Beaucaimia (Liliaceae) и колонновидный кактус Cephalocereus (Cactaceae), Мексика; Е, F — суккулентные кустарниковые заросли с Euphorbia cananensis на Тенерифе; G — Северная Сахара с Acacia raddiana (индикатор грунтовых вод); Н — песчаные дюны с клонообразующими злаками

Таблица 15.7. А, В — маквис с Arbutus unedo (земляничное дерево), о-в Самос; С, D — гаррига с Cistus salvifolius, о-в Крит; Е, F — один из многочисленных видов Arctostaphylos и чапараль в Калифорнии; G — западноавстралийский склерофильный буш с Banksia prionotes; Н —разреженный эвкалиптовый лес, юго-восточная Австралия

Таблица 15.8. А, В — лавролистный лес с Persea indica на Тенерифе; С, D — лавролистный лес с Castanopsis и Ainus в Непале; Е — полугумидный эвкалиптовый лес в Квинсленде; F — вальдивийский дождевой лес в Чили; G, Н — Podocarpus latifolius в лавролистном лесу Гуду, у подножия Драконовых гор, Южная Африка (вокруг злаковники, возникшие в результате пожаров)

Таблица 15.9. А, В — лес с дубом и гикорн, Северная Каролина; С, D —лиственный лес с Noth- fagus alpina в средней части Чили; Е, F — лиственный лес с диким киви (Actimdia) в провинции Сычуань, Западный Китай; G, Н — среднеевропейский буково-дубовый лес с Anemone nemorosa

Таблица 15.10. А, В — наивысший пояс горных лесов соответственно в Австралии, Снежные горы, 1 900м, с Eucalyptus pauciflora и в Альпах, Тироль, 1 950м, с Pinuscembra, С, D — горный смешанный лес в Центральной Швейцарии, 1 200 м, с Fagus, Acer, Abies и P/cea; Е — горный лес в Тасмании, 1 100 м, с Nothofagus, Eucalyptus, Arthrotaxis и гигантскими розетками Richea (Ericaceae); F — горный лес в Казахстане, Тянь-Шань, 1 900 м, с P/cea schrenkiana, Sorbus, Crataegus и Populus; G — горные леса в Чили (38° ю. ш.) Nothofagus с подлеском из бамбука, 1 850 м; Н — Araucaria аrаucarа, 1 400 м

Таблица 15.11. А — подушечные растения Silene acaulis, Центральные Альпы, 2 600 м; В — кустарнички Rhododendron ferrugineum, Центральные Альпы, 2100 м; С — горный луг с эдельвейсом (Leontopodium) и чемерицей (Veratrum), Сычуань, Китай, 3400м; D —горный луг в Снежных горах, Австралия, 2100 м, Craspedia sp.; Е —злаковая пустошь, Драконовые горы, 3 050 м, Южная Африка; F — кустарничково-подушечниковая пустошь, Крэйдл-Маунтин, 1 600 м, Тасмания; G — высокогорная лужайка с Kobresia, хребет Нивот, Скалистые горы, 3 600 м; Н — горная пустошь с Pinus pumila, г Нурикура, 2 800 м, Япония

Таблица 15.12. А — горная степь из Stipa и Leontopodium, Тянь-Шань, 2 500м, Центральная Азия; В — низкозлаковая степь, 2 000 м, Вайоминг; С, D — высокозлаковая прерия с Rudbeckia, 500 м, Миссури, Е — полынная степь из Artemisia terra-alba, 800 м, Казахстан; F — патагонская степь, 500 м, с Muhnum spmosum (Apiaceae) и Stipa speclosa (фото О. Sala); G — полынная степь из Artemisia tridentata, 2 300 м, Невада; Н — альфовый злаковник, Атласские горы, 1 500 м

Таблица 15.13. А, В — Большое Солёное озеро с Suaeda depressa. Юта; С — пустыня Мохаве с Yucca brevifolia, Калифорния; D — пустыня с Atriplex, край Долины Смерти, Невада; Е, F — кустарниковые заросли Haloxylon aphylla близ оз. Балхаш, Центральная Азия, G, Н — каменистая и песчаная пустыня с Ephedra и злаком Aristida, Казахстан

Таблица 15.14. А — типичный бореальный лес с Picea obovata, северо-восточная Финляндия; В — решающая роль рельефа в размещении леса и болота, северная часть Швеции; С, D, F — бореальный березовый и ивовый заболоченный лес; Е — пышное высокотравье; G — кустистые лишайники; Н — сухой смешанный березово-сосновый лес

Таблица 15.15. А, В — низинное болото с Eriophorum (пушица) и сразу за ним (фото В) приподнятое плоскобугристое болото; С, D — осоковая тундра соответственно под тающим снежником и с бугорками мерзлотного пучения; Е, F — тундра с карликовыми ивами; G, Н — Empetrum nigrum и Carex bigelowii — два доминирующих циркумполярных вида (все примеры из северной части Швеции)

Таблица 15.16. A — Cocos nucífera, Карибское побережье; В, С — Pandanus и Casuanna, Большой Барьерный риф; D, Е — Rhizophora mangle, Флорида; F — Avicennia marina,Квинсленд; G — сформированный под воздействием ветра береговой лес южнее Сиднея; Н — лагуна с Tamarix, острова Греческого архипелага; I — засоленный марш, Камарг, южная Франция; J — береговые дюны и погребенный под ними лес, Орегон, США; К — дюнный ландшафт, побережье Северного моря; L — Convolvulus, Корсика; М — О — скалистое побережье Средиземного моря (М, N — Иския, О — Самос, N, О — Euphorbia dendroides); P — R — литораль, северная Атлантика (Р — различные водоросли-макрофиты, R — чистые заросли Fucus)

(5) В горы дуб и его спутники поднимаются немного выше, чем бук. Дубовые и смешанные леса с господством дуба обладают более разреженными кронами, чем буковые, и поэтому в них богаче кустарники и летние растения нижнего яруса. Дуб — это «светлое дерево», а бук — «теневое». Из хвойных лесов здесь находятся:
(6) Сосновые леса (из Pinus sylvestris), главным образом на бедных, сухих песчаных почвах плоских и холмистых равнин.
(7) Ель (Picea abies) обычна на равнинах только в Северо-Восточной Европе, в Центральной Европе это дерево среднего и верхнего поясов горных лесов (рис. 15.5, А,В; 15.10, D).
(8) Пойменные леса будут рассмотрены отдельно далее.
Под воздействием текучих или стоячих вод развиваются речные поймы, ряды их зарастания, заболоченные леса и болота (см. рис. 14.35, 14.36, 14.41, 15.3, 15.5, G, Н, I). Их экологическая дифференциация основана на степени затопления в половодье, содержании в почве питательных веществ и обогащенности органическим веществом в анаэробных условиях (торфообразование). В условиях избыточного увлажнения деревья могут перестать развиваться.
Живой мир речных пойм, тянущихся вдоль рек и ручьев, должен приспосабливаться к сильным и нерегулярным колебаниям водного уровня (см. рис. 14.35, 15.5, G, Н). Седиментация (речные наносы) и эрозия (снос грунта) постоянно изменяют естественные ландшафты пойм. Затопление оказывает отрицательное влияние на дыхание корней и является причиной механических повреждений (особенно влияют движения льда, перемещение гальки, песка и речного ила), но оно и обогащает поймы питательными солями и продуктами органического опада. При спаде воды, открытые галечные и песчаные почвы, могут сильно нагреваться с поверхности и просыхать до большой глубины. Интенсивность этих воздействий ступенчато снижается по мере снижения уровня затопления, от низко лежащего прируслового уровня по направлению к свободному от затопления высокому уровню краевой части поймы (см. зонирование растительности, см. рис. 14.35).
В стоячих водах(заводях, затонах) отложение неорганического материала прекращается; зато, по мере поступления отмерших остатков растений и животных, здесь образуется органогенный слой (ил) или торф, в результате чего толщина слоя воды постепенно сокращается. Поскольку водная и прибрежная растительность развивается в соответствии с глубиной воды, постепенно происходит центростремительное перемещение отдельных растительных сообществ, и, наконец, водная поверхность исчезает (заиление, см. рис. 14.36, 15.5,1). В богатых питательными веществами (эвтрофных) стоячих водах за счет обильною здесь планктона образуется специфический вид ила, называемый гиттией, при высоком содержании карбонатов может развиваться белый «известковый морской ил»; он представляет собой ценный материал для интерпретации истории климата, являясь своеобразным «климатическим архивом», поскольку содержит большое количество остатков растений, животных и планктона.
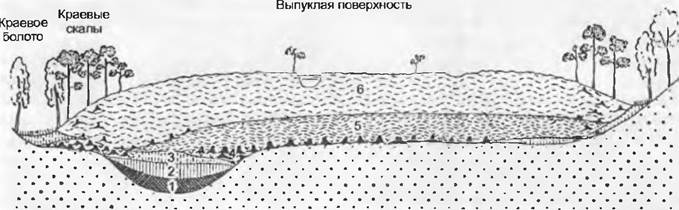
Болотами называют залежи торфа и развивающийся на них растительный покров. Торф — это отложения отмерших мхов и высших растений, которые находятся в стадии постепенной углефикации в условиях отсутствия кислорода, при этом структура их тканей может сохраняться очень долго. При зарастании водоемов или на заболоченных минеральных почвах образуются плоские болота (низинные). Их воды более или менее богаты питательными веществами, образующийся в них торф чаше всего имеет слабокислую или даже нейтральную реакцию (тростниковые, осоковые или лесные болота и заболоченные леса; см. рис. 14.41). В условиях климата, характеризующегося обильными атмосферными осадками, на их поверхности в течение длительного времени могут развиваться влаголюбивые торфяные мхи (виды рода Sphagnum), отмершие нижние части которых пропитываются влагой, при этом верхние слои все время нарастают вверх и растения, существовавшие на этих местах (в том числе и древесные насаждения), отмирают. Такие очень бедные питательными веществами болота, получающие их только за счет атмосферных осадков и воздушного переноса пыли, а именно выпуклые верховые болота(рис. 15.3, см. рис. 15.5, К, L), могут подниматься на несколько метров нал окружающей поверхностью, имея выпуклую форму типа часового стекла. По периферии их возвышенной поверхности имеются краевые болота, по растительности, соответствующие плоским. Выпуклая поверхность таких болот обычно покрыта мелкими бугорками, поросшими большей частью вересковыми кустарничками, кочками влаголюбивых осок, мочажинами. Лишь немногие виды цветковых растений могут существовать на верховых болотах, это, например, Calluna vulgaris, Vaccinium oxycoccos, V uliginosim, Andromedapolifolia (все — вересковые); Eriophorum vaginatum, Trichophorum cespitosum и другие осоковые, а также насекомоядные виды росянок (Drosera).
Близ морских побережий (в условиях сухого степного климата также и во внутриматериковых районах) растительный покров развивается в условиях избыточного содержания в почве соли (галофитная растительность). На территории Центрально-европейской флористической области на побережьях Северного и Балтийского морей в особенности типичны засолённые марши (табл. 15.16, 1) и береговые дюны.
Рис. 15.3. Схема строения слоев центрально-европейского верхового болота (в разрезе), возникшего на месте озера: 1 — ил; 2 — тростниковый торф; 3 — осоковый торф, частично возникший при заболачивании леса; 4 — лесной торф; 5 — древний сфагновый торф; 6 — молодой сфагновый торф. В середине выпуклой поверхности наполненная водой промоина — мочажина. Точками обозначен лежащий ниже минеральный грунт
На немецком побережье Северного моря развитие растительности часто идет по типу зарастания ваттов. Это мелкие участки морского дна, на которых отлагаются богатые питательными веществами илисто-песчаныс осадки, и которые при отливе большей частью пересыхают. Типичную последовательность смены видов вдоль градиента засоления иллюстрирует рис. 13.28. Ниже водной поверхности растут морские травы (Zostera, Ruppia). Наносные илы до границы средне высоко го уровня прилива зарастают видами солероса (Salicomia agg.). На менее регулярно затопляемых местах приморских террас развиваются бескильницевые луга, где доминирует злак Puccinellia maritima. На еще более высоком уровне их сменяют овеяницевые луга с Festuca rubra agg., Armería maritima и др., а затем полностью лишенные засоления сухие луга и пионеры лесной растительности. Развивающиеся на отложениях илисто-песчаных морских осадков луга (см. рис. 15.5, N, О) называют маршами (см. рис. 15.5, М). Можно искусственно содействовать развитию этой растительности путем создания запруд и таким образом создавать новые земли.
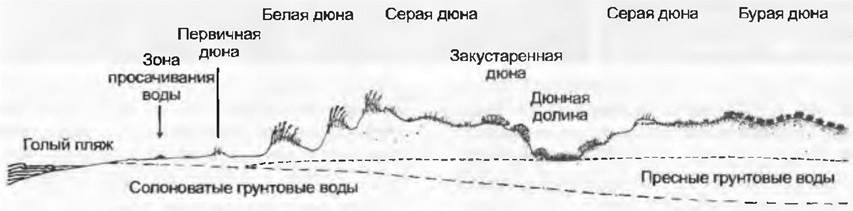
На песчаных морских побережьях образуются дюны (рис. 15.4, табл. 15.16). На еще довольно увлажненных и засоленных участках в зоне просачивания воды развиваются сначала сообщества однолетников (Cakile maritima, Salsola kali, Atriplexprostrata и др.), затем появляются пыреи с ползучими корневищами (сначала Agmpyron junceum, затем Elymus farctus).Создаваемая ими ветровая тень способствует отложению перевеваемых ветром песков, образуются мелкие «первичные дюны». Атмосферные осадки постепенно вымывают соли из этих скоплений песков, и они становятся пригодными для заселения прежде всего песколюбкой (Ammophila arenaria). При этом формирование дюн продолжается. Большинство растений дюн все больше нарастает в слоях вновь отлагающегося ветром песка, пронизывая их корнями и образуя крупные клоны, чем делают эти вторичные «белые дюны» все крупнее и выше (см. рис. 13.24, табл. 15.16, К). Когда дюна становится уже не столь подвержена развеванию ветрами (часто перед ней формируются новые дюны), она полностью захватывается растениями, переходя в третью стадию — «серой дюны». На островах Северного моря на дюнах развиваются кустарничковые сообщества с Salix repens и Hippophae, или с Еmpetrum и Calluna, на Балтийском побережье господствуют сосновые леса. Продолжаются процессы почвообразования, формируются «бурые дюны». Если плотный растительный покров разрушается, образование дюн может начаться снова (подвижные дюны, например, на острове Зильт; также примеры из Орегона в табл. 15.16, J).
Рис. 15.4. Образование и зарастание дюн на побережье Северного моря: от берега моря в глубь страны снижается концентрация солей и усиливаются почвообразовательные процессы «коричневые» дюны в естественных условиях уже покрыты лесами
На поверхностных, бедных влагой почвах в южных районах формируются сухие лужайки, на которых много более восточных и южных видов растений (например, Pulsatilla, Stipa, Artemisia, Astragalus, Fumana, Teucrium). Когда почвы более мощные, они заселяются кустарниками (например, Comus sanguinea, Viburnum lantana), и в конце концов покрываются вышеупомянутыми термофильными смешанными дубовыми лесами. На силикатных скалах, бедных карбонатами песчаных почвах, а на равнинах также на узких не покрытых лесом полосах вдоль побережья (местами и на очень кислых болотных почвах) находятся естественные местообитания кустарничковых пустошей, растительность которых сложена такими низкорослыми вересковыми, как Calluna vulgaris. Особенно типичны развившиеся в условиях океанического климата и под влиянием длительного антропогенного воздействия (выпас скота, выжигание) пустоши северо-западной Германии на бедных песчаных и подзолистых почвах (например, Люнебургская пустошь). Здесь помимо доминирующего вереска (Calluna) произрастает единственное древесное растение, устойчивое к потраве скотом — можжевельник (Juniperus communis). В настоящее время большая часть ландшафтов этих пустошей повторно облесена или используется в земледелии.
Еще интенсивнее используются человеком луга и пастбища. Эти сельскохозяйственные угодья занимают в Германии и Австрии свыше 20 % общей площади, являясь основой для развития мясного и молочного скотоводства (см. 13.9.3). Большая часть лугов занимает облесенные в прошлом участки и благодаря постоянному кошению (сенокосные луга) или выпасу (пастбища) деревья на них не растут. В соответствии с характером почвы и видом использования (см. рис. 13.53) возникают различные типы лугов: малопродуктивные луга (на бедных почвах), они выкашиваются только один раз в году и удобряются слабо (основной вид, произрастающий на бедных известью почвах — Agrostis tenuis, на богатых — Bromuserectus). Тучные, или высокопродуктивные луга с разнообразным видовым составом (на богатых почвах) косят 2 — 3 раза в год, и потом часто еще используют как пастбища. Они требуют постоянного и значительного удобрения (основные виды — на низменных участках Arrhenaterum elatius, на возвышенных — Trisetum Jlavescens). Болотистые луга, как правило, не удобряют, а сено с них используют только в качестве подстилок в хлевах. На их длительно увлажненных почвах господствуют различные осоки (Сагех), это так называемые «кислые луга», а на периодически влажных почвах — молиния (Molinia caerulea). К сухим почвам приурочены закустаренные умеренно сухие луга (выгоны), с высокимвидовым разнообразием составленные Festuca ovina agg., Bromus erectus, Brachypodium pinnatum. Сейчас имеется также много сеяных лугов бедных видами, в которых доминируют Lolium perenne и луговой клевер. Почти треть площади Центральной Европы и большую часть пригодных для земледелия участков занимают в настоящее время интенсивно используемые культурные земли (пашни, сады, фруктовые плантации) и примыкающие к ним засоренные угодья (см. рис. 13.50,15.5, Р — R).
15.1.2. Верхние горные леса и альпийский пояс
В верхних горных поясах Альп и Карпат (верхнем горном, субальпийском, альпийском, нивальном) существует характерная флора от тысячи и более видов сосудистых растений, частично эндемиков. Их родственные связи указывают на происхождение большинства видов от близких форм равнин южной Европы, остальные связаны с горами Европы и Азии, или даже с Арктикой. Наличие эндемиков свидетельствует об относительной самостоятельности развития альпийской флоры и возможности ее существования на периферии ледников в периоды оледенений. Наконец, широко распространенные и многочисленные в настоящее время виды с дизъюнктивными бореально-монтанными (среднегорными) или аркто-альпийскими ареалами подтверждают факт интенсивного флористического обмена, имевшего место на холодных этапах четвертичного периода и в послеледниковое время между Альпами и Карпатами, с одной стороны, и южно-европейскими (Crocus, Dianthus, Helianthemum) или азиатскими (Primula, Leontopodium) горами, а также с циркумарктическими (Oxyria, Saxifraga) и бореальными (Empetrum, Vaccinium) пространствами — с другой. Европейско-(монтанно)-альпийским элементом являются, к примеру, Soldanella, Aster и Geum.
Растительность центрально-европейских гор можно поделить на высотные пояса, или пояса растительности (рис. 14.44, 15.2). Существенными факторами являются понижение температуры с высотой, укорочение вегетационного периода, увеличение длительности существования снежного покрова и другие особенности горного климата. Они четко соответствуют растительным зонам (умеренной, бореальной, арктической; рис. 14.22) и связанным с последними растительными формациями (рис. 15.6, 15.7).
Рис. 15.5. Культурные и естественные ландшафты Центральной Европы: А — высотные пояса на склоне альпийской долины (Хайлигенблют, Каринтия). Нижняя часть пояса горных лесов преобразована в культурный ландшафт. Граница леса образована елью и лиственницей на высоте 2000 м; В — древний культурный ландшафт в горном районе (Лехталерские Альпы, 1 600 м); С — рельеф и экспозиция в альпийском поясе способствуют развитию очень разнообразной мозаики сильно различающихся микроэкотопов. Лето и зима существуют рядом (2 500 м, горный перевал Фурка, Швейцария). Буково-дубово-грабовые богатые видами лиственные леса на верхнем Рейне в горизонтальной и вертикальной проекции — пример центрально-европейской лесной экосистемы пояса холмов с высоким биоразнообразием (12 видов деревьев); D — фаза весеннего распускания листвы; Е — середина лета; F — начало осеннего расцвечивания листвы. Молодая (G) и сформированная (Н) затопляемая пойма проточных вод и пруда (I) — пример растительности стоячих вод (см. рис. 14.36); J — L — атлантическая пустошь с Ulex europaeus и верховое болото со следами торфоразработок в Западной Ирландии. Раскопанный пень сосны (К) указывает на то, что 1 600 лет назад здесь был лес. Запруженный марш (М) и мероприятия по увеличению земельных площадей на ваттах путем создания дамб (N, О) на ютландском побережье Северного моря (амт Рибе, Дания); Р — сильно структурированный культурный ландшафт в южном Эльзасе (Лейменталь); Q — «жизнерадостные» аграрные пространства в Мархфельде близ Вены; R — полуестественный злаков- ник и засолённый луг на паннонской низменной равнине (юго-восточнее Кешкемет, Венгрия)

В Альпах, частично также в более высоких среднегорьях, можно выделить следующие высотные пояса (самый нижний уже описан в разделе 15.1.1, данные по их высотам, в метрах, приведены для Альп):
✵ равнинно-холмистый: пояс плоских и холмистых равнин, до 300 — 500 м;
✵ субмонтанный (низкогорный): переходный пояс самых низких горных лесов, до 400 — 700 м;
✵ монтанный (среднегорный): нижний (600-1 100 м), средний (1000 — 1500 м) и верхний (1 400 — ок. 2 000 м) пояса горных лесов;
✵ субальпийский: пояс редколесий и криволесья, ок. (1 700) 1 900 — 2 200 (2300) м;
✵ альпийский: сомкнутые кустарничковые и травяные пустоши, до 2 500 — 3 000 м;
✵ субнивальный: пятна растительности и отдельные растения, до примерно 3 000 — 3 300 м;
✵ нивальный: снежная зона, открытые участки выше климатической границы снегов: в защищенных местообитаниях пионерные сосудистые растения до 4 450 м.
Границы отдельных высотных поясов также колеблются в пределах отдельных горных цепей в зависимости от рельефа, экспозиции и субстрата. Во внутренних частях гор растительные пояса располагаются выше, чем на краевых цепях (эффект увеличения массы). Граница леса, а именно верхний предел сомкнутых высокогорных лесов (англ. timberline или forest line) не является собственно «границей», но лишь нижним краем переходной зоны, в которой лес постепенно перестает быть сплошным, чередуясь с языками безлесной альпийской растительности (см. рис. 15.5, А, табл. 15.10, В). Линия, которая соединяет последние группы деревьев, обозначается как граница деревьев (англ. treeline), а верхняя граница распространения отдельных карликовых или подушковидных деревьев — как граница древесных видов (англ. tree species line). Эти переходные зоны вместе называют лесным экотоном (англ. treeline ecotone). Очень неоднозначно употребляемое понятие «субальпийский» как раз подходит в первую очередь к этой переходной зоне, которая благодаря наличию леса еще не является «альпийской», а представляет собой мозаику обоих элементов.
В смешанных хвойных лесах (океанического) монтанного пояса доминируют ель и пихта (Picea abies и Abies alba). В континентальных районах и на более высоких уровнях (большей частью на бедных подзолистых почвах с развитым кислым горизонтом грубого гумуса) пихта отступает. Типичные виды нижнего яруса в этом случае — черника и брусника (Vaccinium туг- tillus, V vitis-idaea), папоротники (например, Blechmtm spicant), злаки (например, Calamagrostis villosa). В верхнегорных лесах Центральных Альп ель с высотой сменяется кедровой сосной (Pinus cembra) и летне-зеленой лиственницей (Larix decidua).
В субальпийскойпереходной зоне на открытых участках между фрагментированными лесами развиты кустарничковые сообщества с Rhododendron и Vaccinium, закус- таренные опушки, естественные луга, дернистые лужайки на местах схода лавин, а также кустарниковые сообщества из ольхи зеленой (Alnus viridis = A. alnobetula) или горной сосны (Pinusmugo). Отдельные экземпляры деревьев, изуродованные в результате возрастающих повреждений, обычно вместе с этими кустарниками образуют так называемое криволесье (этот пояс называют еще «зоной борьбы за существование»).
Альпийские растительные сообщества (рис. 15.5, С, табл. 15.11, А, В) кроме Альп и Карпат в обедненном виде имеются еще в Судетах. В нижнем альпийском поясе также господствуют кустарничковые пустоши, особенно с видами Vaccinium, а на наветренных склонах (см. рис. 12.9, 12.13) с очень устойчивой Loiseleuria procubens (мелколистный стелющийся вересковый кустарничек). Рядом с ними и выше находятся сообщества дерновинных растений, которые с высотой все более изреживаются и обедняются: на кислых почвах близ границы леса (на пастбищах и ниже) часто доминирует белоус (Nardus stricta), в верхней части кустарничкового пояса — осока искривленная (Carex curvula), на карбонатных почвах — сеслерия (Sesleria varia), а на подверженных ветрам гребнях подушковидная осока (Carex fuma). Важнейшие растения щебнистых осыпей и скал относятся кродам Androsace, Draba, Gentiana, Minuartia, Oxyria, Saxifraga, Silene и др. (см. 15.2.11). В мульдах, где долго залеживается снег, образуются нивальные лужайки с очень характерными растительными сообществами из низкой ползучей ивы (Saliх herbacea) и видов Soldanella. Наконец, до нивального пояса доходят только немногие цветковые растения, например, лютик (Ranunculus glacialis), виды рода Saxifraga (высотный рекорд для Альп: Saxifraga biflora встречена на высоте 4450 м, на вершине Дом-де-Мишхабель, кантон Валлис).