БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
12. ОСНОВЫ ЭКОЛОГИИ РАСТЕНИЙ
12.3. Фактор времени и нелинейные реакции
12.3.1. Фенология и биологическое хронометрирование
Для ботанико-экологических наблюдений применение устойчивых линейных мер времени (час, календарная дата) проблематично в том случае, если нужно сравнивать реакции организмов, связанные с динамикой развития. При определенных обстоятельствах, особи одного и того же вида растений, например, могут зацвести уже через 40 дней после посева, при других условиях — через 80. Для сравнения лучше выбрать определенную фазу развития или фенологическое состояние (например, раскрытие первого цветка), а не определенную дату. Таким образом уделяют внимание фенологии (видимое изменение состояния в процессе развития) и подгоняют эти наблюдения под фенофазы. Характерные фенофазы — прорастание или распускание листьев, цветение, плодоношение, листопад или начало старения (физиология развития, см. 7.7). Последовательность этих моментов постоянна, но длительность интервалов между ними вариабельна.
Фенометрия измеряет изменения определенных параметров в течение прохождения фаз развития растения. Фенологические ряды представляют собой результат интеграции внешних условий и внутреннего состояния растения, иначе говоря — биологический календарь. Благодаря ценности такого рода биологически интегрированных чисел для характеристики хода погоды фенологические даты используются метеорологической службой. Ретроспективно эти даты имеют чрезвычайное значение для суждений об изменениях климата, так как никакие «нормальные» метеорологические измерительные величины не обладают такой чувствительностью. Для разрешения подобных задач (при условиях, не ограниченных засухой) наиболее подходит так называемое термическое время (англ. thermal time) — продукт суммирования температур (обычно среднесуточных) за период определенной длительности (если это число дней, то суммируются суточные температуры; англ. day degrees); также можно просуммировать и среднечасовые значения (degree hours). Нередко можно обнаружить тесные корреляции между такими «весомыми» мерами времени и фенологией.
Часто бывает особенно интересно понять динамику вегетативного развития растения в промежутке между двумя такими фенологическими «крупными событиями», как прорастание и цветение. Неоднократно используемая временная мера для последовательного процесса вегетативного развития — пластохрон. Пластохрон соответствует отрезку времени между появлением двух следующих друг за другом листьев на одном побеге. Возраст развивающегося побега можно приблизительно определить по числу появившихся листьев с момента определенного исходного временного пункта (стадия 5 листьев равна приблизительно 5 пластохронам). Такие измерения играют особенно большую роль в сельскохозяйственных исследованиях для определения возраста побегов. Данные о возрасте (в днях) имели бы меньшую ценность для сравнений.
12.3.2. Нелинейность и частота
Нелинейность связей процессов с окружающей средой является правилом, а не исключением. Типичные примеры — воздействие температуры на дыхание и рост или света и СО2 на фотосинтез (см. 13.7). Полученные для одной области данные измерений не могут быть экстраполированы на другую, как это имело бы место при линейности связей (в пределах определенного доверительного интервала), пока не известен характер кривой.
Особую проблему представляют при этом сильные и нерегулярные колебания внешних условий, поскольку их низкие и высокие измеряемые величины имеют непропорциональные воздействия. Наряду со знанием графической характеристики связи процесса с внешними условиями для их реконструкции или прогнозов также необходимы параметры внешней среды, причем высокодискретные во времени, а не их средние значения. На стандартной кривой нормы реакции средние значения варьирующих во времени внешних условий дали бы фальшивую картину, не соответствующую их реальному влиянию на процесс.
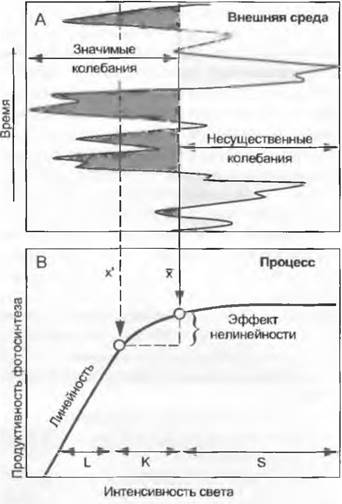
Особенно наглядно это можно продемонстрировать на кривой воздействия света на фотосинтез (рис. 12.3). Фотосинтез в листьях большинства растений приближается к насыщению при 25 % от полного солнечного излучения. Увеличение интенсивности излучения между этим порогом насыщения и максимумом (100%) в листьях, перпендикулярно ориентированных к лучам света, не дает прироста фотосинтеза. При очень слабом свете, напротив, фотосинтез очень чувствителен к малейшим колебаниям освещения. Точное установление варьирующих величин интенсивности излучения в области слабого света имеет очень большое значение, в области насыщения — практически незначимо. Однако арифметическое среднее всех измеряемых величин придало бы одинаковый вес величинам интенсивности фотосинтеза как в области насыщения, так и в области слабого света.
Рис. 12.3. Нелинейные реакции на вариабельную внешнюю среду. Большинство жизненных процессов, например, продуктивность фотосинтеза (В) реагируют на изменения внешней среды нелинейным образом, например, на меняющуюся во времени интенсивность освещения (А). Средние значения (х) таких состояний среды непригодны для того, чтобы прогнозировать интенсивность процесса на основе нормы реакции (В). В этом примере изменения значений во времени в области S несущественны, так как в этой области рассматриваемый процесс насыщен. Изменения слева от х (ненасыщенная область) оказывают воздействие, но в области К нелинейным образом. При очень низких значениях переменных внешней среды отношения вначале могут приобретать линейный характер L (англ. initial slope). Чтобы в этом примере рассчитать суммарное или среднее значение интенсивности процесса, нужно каждое отдельное значение освещенности сопоставить с соответствующим значением интенсивности фотосинтеза или представить состояние фактора среды в форме распределения частот и умножить по классам
Итак, поскольку временные ряды переменных внешней среды в большинстве случаев для целей анализа воздействий не могут быть усреднены, необходимо либо иметь оригинальные временные данные, фиксируемые с достаточно тонким интервалом (например, для освещения — менее 2 мин), либо табелировать результаты по классам освещенности. Из практических соображений по большей части выбирают второй вариант. Распределение частот состояний внешней среды — удобная мера для того, чтобы судить о варьирующих во времени воздействиях на растения. При каждой оценке влияния среды нужно исходить из «временной ткани» его интенсивности (отсюда — гистограмма; histos — ткань) или концентраций рассматриваемого влияющего фактора. Диаграмма частот — это до некоторой степени эмблема функциональной экологии (см. 12.4).