БОТАНИКА ТОМ 4 - ЭКОЛОГИЯ - 2007
12. ОСНОВЫ ЭКОЛОГИИ РАСТЕНИЙ
12.5. Экосистема и ее структура
Под экосистемой понимается совокупность взаимодействующих абиотических и биотических компонентов в какой-либо ограниченной области. Абиотическая матрица (местообитание) обозначается как биотоп, а живая составляющая — как биоценоз. Эта сильно упрощенная схема разъединяет то, что само по себе неразделимо, поскольку биоценоз постоянно изменяет и создает биотоп; даже его первоначальное пребывание создает свойственное ему жизненное пространство (биотоп), поскольку в абсолютно неживой форме его не бывает. Еловые леса ведь не растут изначально на свойственных им лесных почвах, но такие почвы формируются там, где достаточно долго росли ели. Эта дихотомия — биотоп-биоценоз символизирует тем не менее изменения в физико-химических условиях вместе со связанными с ними организмами.
12.5.1. Структура биоценоза
12.5.1.1. Иерархическая структура
Биоценоз — это совокупность всех живых организмов экосистемы: растений, животных, грибов и микроорганизмов. Для совокупности всех растений существует понятие фитоценоза, или растительного сообщества (англ. plant community). Каждый вид такого «товарищества» (фитоценология, см. гл. 14), как правило, представлен многими особями, совокупность которых именуется популяцией (размножающееся товарищество). Популяция включает все возрастные состояния вида, а также те особи, которые присутствуют незаметно в почве в виде семян. Популяция состоит из генетически различных особей или генетически различных групп индивидов, возникших клональным способом (генетически тождественных в пределах клона), — триб (англ. ramets). При этом индивидуум (особь) воплощает собой определенный генотип или в случае генетически идентичной отдельной раметы — часть генеты (англ. genet — синоним клона). Поскольку возникшие из семян особи также могут быть генетически идентичны (апомиксис, см. 10.1.3.3), клоны, содержащие генетически однородные особи, встречаются чаще, чем можно было бы предположить исходя из их морфологии (например, апомиктический вид Taraxacum officinale). Аналогичная иерархическая структура имеется и в сообществах животных, грибов и микроорганизмов.
12.5.1.2. Таксономическая структура
Характер экосистемы создает присутствие определенных видов растений. По ним можно судить о местных условиях существования, поэтому часто в начале анализа экосистемы устанавливается список видов (индикаторные виды, см. 12.5.2.2; индикаторное значение, см. 14.3.3). Выявление числа видов и относительного обилия (частоты) отдельных видов — важная биологическая инвентаризация, обозначающаяся в фитоценологии (см. 14.3.3) немецкоязычных стран как структура сообщества (Bestandesstruktur). Поскольку английское слово «structure» (структура) почти исключительно используется для обозначения пространственной структуры (см. 12.5.1.5), целесообразно применить здесь понятие «видовой состав». Понятие биоразнообразия обычно применяется для количественной оценки числа видов. Тем не менее оно также включает внутривидовое генетическое разнообразие и надвидовой уровень, а именно многообразие растительных сообществ (см. 14.2.4.1).
12.5.1.3. Функциональная структура
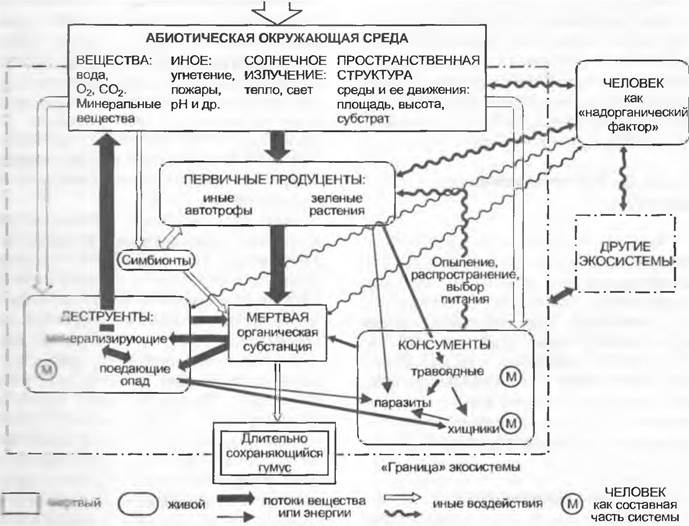
Все фотосинтетически активные организмы объединяются понятием первичных продуцентов. Им противостоят консументы (поедающие живые организмы) и деструенты (редуценты, разлагающие органическое вещество) Консументы функционируют непосредственно как травоядные животные (фитофаги, питающиеся растениями) или опосредованно как хищники (первого, второго или третьего порядка). Мертвые органические остатки в конце концов разлагаются самыми различными деструентами. К ним в особенности относятся дегритофаги (детрит — мертвый опад; поедающие опад клещи и черви) и минерализаторы (бактерии и особые грибы). Такие пищевые цепи, или лучше пищевые сети (англ. food web), связывают друг с другом членов каждой экосистемы, как и потоки энергии и круговорот веществ. Эти связи могут быть обратными и делают возможной в ограниченном объеме саморегуляцию, направленную против внешних изменений (рис. 12.4).
Рис.: 12.4. Концепция экосистемы. Упрощенная схема совершенной экосистемы, т. е. широкой саморегулирующейся структуры взаимодействий живых организмов и окружающей среды (объяснения см. в тексте)
Внутри растительного сообщества можно различать функциональные группы, или функциональные типы (англ. functional types). Теоретически их так же много, как и видов (см. введение к гл. 12), если же рассматривать функционально очень различающиеся возрастные состояния одного индивидуума (например, проросток — дерево), то их даже больше, чем видов. Концепция функциональных групп направлена на то, чтобы свести часто огромное разнообразие видов к небольшому числу «сходных по функционированию» категорий, что необходимо прежде всего для моделирования экосистем и теоретических обобщений (генерализации). Существует много таких попыток группирования; приведем некоторые важные. Наиболее простая и, пожалуй, наиболее используемая — это группирование по морфотипам (травы, кустарники, деревья или глубоко укореняющиеся в сравнении с поверхностно укореняющимися розеткообразуюшие граминоиды и т. д.) и фенотипам (однолетние, многолетние, летнезеленые, вечнозеленые и т. д.; фенология. см. 7.7. и 12.3.1). При группировке по физиотипан исходят из особенностей обмена веществ, как, например, по типам фотосинтеза: С3-, С4-, или САМ-типы (см. 6.5.7 — 6.5.9), по потребности в свете соответственно положению в сообществе (теневые и солнечные растения), специализации к субстрату (кальцефилы и кальцефобы) или свойствам резистентности (к засухе, засолению почвы, морозу, жаре). Симбиотипы основываются на способности к симбиозу с азотфиксирующими бактериями или специальными микоризными грибами. Критериями для объединения в группы могут служить и
различные другие формы мутуализма и паразитизма.
Концепция, изначально инспирированная зоологией, — это группирование по господствующим жизненным стратегиям (англ. life strategies), ставшее классическим разделение растений по образу жизни на г-стратегов (первая буква слова «reproductive», что подчеркивает их устойчивость) и К-стратегов (первая буква слова «kompetitiver», что подчеркивает их преимущественно вегетативный способ размножения); с течением времени было развито много различных концепций стратегий растений. Особенно известна система треугольника Грайма, или CSR-стратегов (конкуренты, стресс-толеранты, рудералы). Она выводится из двумерной матрицы, в которой растения группируются по их толерантности к стрессу и нарушениям (малым или большим); одна из четырех комбинаций (сильный стресс и много нарушений) рассматривается как несуществующая, и остаются три категории. Они маркируют в углах треугольника экстремальные ситуации. Каждое растение занимает свое место между этими экстремами. Расстояния от угловых пунктов называются радиусами; например, стресс-радиус свидетельствует о степени стресс-толерантности вида. Как и каждая попытка типизировать многообразие жизненных ситуаций, эта концепция также критиковалась как слишком упрощенная.
П. Грабб (Р. Grubb), напротив, считает, что категория стресс-толерантных растений должна быть отнесена к трем различным стратегиям, поскольку их поведение в течение жизни может изменяться. Ученый различает виды, которые сохраняют свой тип стратегии на протяжении жизни от всходов до взрослого растения (low flexibility strategy — стратегия низкой пластичности), виды, которые только в состоянии всходов хорошо переносят стресс (switching strategy— переключающаяся стратегия), и наконец виды, которые могут перестраиваться и в известной степени менять тип и переходить в другую категорию, когда условия очень благоприятны (gearing strategy — приспособительная стратегия). Признаками стресс-толерантности наряду с другими являются медленный рост, многолетние органы, низкое репродуктивное усилие, сравнительно толстые листья с низким содержанием азота.
Перечень примеров типизации и группирования можно продолжать еще долго. Опыт показал, что функциональные группы часто обнаруживают предполагающиеся для них общие функции только очень ограниченно и видоспецифические различия в пределах функциональных групп часто значительнее, чем между самими группами. При группировании может оказаться так, что особи разного возраста будут отнесены к различным функциональным группам, что затрудняет попытки группирования видов. Для разных целей предпочитают и разные способы группирования или даже создают новые группы (например, группы видов, устойчивых к озону, тяжелым металлам или застойному увлажнению). Морфо- и фенотипы могут по-прежнему представлять собой наиболее практичные функциональные группы для нетаксономического описания структуры фитоценоза.
12.5.1.4. Материальная структура
Для наименования и разделения растительной субстанции в экосистеме установлены международные правила (все данные без исключения приводятся в сухом весе после высушивания при 80 — 100 °С). Живая, надземная и подземная растительная составляющая экосистемы называется биомассой растений. В биомассу включаются также омертвевшие внутренние ткани (одревесневшие структуры), которые все же относятся к живому растению. Наружные отмершие части растений (надземные и подземные) обозначаются как некромасса (англ. standing dead). Совокупность всех живых и отмерших наружных частей растений называется фитомассой. Ей противопоставляются отделившиеся мертвые части растений в экосистеме — подстилка (англ. litter), причем различают напочвенную подстилку (лежащую на поверхности почвы) и подземный опад (например, отмершие корни). Органические остатки, в которых простым глазом больше нельзя распознать структуру органа, относят к гумусовому комплексу (англ. soil organic matter, SOM), он охватывает все переходы от грубого гумуса до комплекса молекул гуминовых кислот в органической составляющей почвы. Органическая субстанция животного происхождения (большей частью мелких почвенных животных) и микроорганизмов очень незначительна (<0,1%) и обычно прибавляется к углеродной составляющей почвы или вообще игнорируется, что, конечно, ничего не говорит о важной функции этих организмов в экосистеме.
12.5.1.5. Пространственная структура
Способ освоения воздушного пространства и почвы определяет не только общий внешний облик, но и свойства экосистемы. Морфология побегов и корней доминирующих видов растений, прежде всего геометрия или архитектура сообщества, придает каждой экосистеме свое неповторимое своеобразие, определяет, где происходит преобразование энергии, откуда поступают вода и питательные вещества. Важнейшие признаки надземной структуры — высота доминантов, индекс листовой поверхности и вертикальное размещение листовой поверхности, подземной — типы корневых систем, максимальная глубина проникновения корней и вертикальное распределение корней в почвенном профиле.
Индекс листовой поверхности LAI (англ. leaf area index) — безразмерная единица, которая показывает, сколько квадратных метров листовой поверхности приходится на 1 м2поверхности почвы (фактическая листовая поверхность независимо от ее ориентации в пространстве; при толстых или не плоских листьях это их наибольшая площадь проекции). В сомкнутых растительных сообществах на хорошо развитых почвах с достаточным водоснабжением LAI достигает значений 5 — 8. Для большинства посевных растений максимальное значение LAI достигает почти 4. Плотность листовой поверхности LAD (англ. leaf area density), измеряемая в метрах в минус первой степени (м-1), дается для всего сообщества в целом, если LAI разделить на высоту сообщества или на высоту отдельных ярусов (отношение величины листовой поверхности в м2 к 1 м3 объема сообщества). LAI и LAD определяют профиль освещенности в сообществе (см. 13.1.3).
Большая часть тонких корней находится обычно близко к поверхности почвы (<1 м), часто даже в верхних 20 см почвенного профиля, там же сосредоточена большая часть минерализаторов и микоризных грибов. Часть корневых систем может достигать значительной глубины (см. табл. 13.3). Такие глубоко проникающие корневые системы служат в основном для обеспечения водой. Ярусное распределение корней отдельных видов в почвенном профиле — классический пример дифференциации ниш, которая в значительной степени определяет биоразнообразие. Глубоко проникающие корни могут также служить для увлажнения верхних почвенных горизонтов (гидравлический лифт, см. 13.6.4). Многообразие типов корневых систем часто превосходит разнообразие типов побегов и является существенным структурным признаком каждой экосистемы.
12.5.2. Биотоп: местообитание и факторы внешней среды
12.5.2.1. Местообитание и местопроизрастание
Факторами местообитания (в определенный период времени при фиксированных данных местности) называют климат, рельеф, почву и биотические воздействия других присутствующих организмов (например, затеняющие лесные деревья). Им противостоят непосредственно действующие и сильно варьирующие в короткие промежутки времени факторы внешней среды: фактическое количество солнечной радиации, тепло, влажность. химические факторы, а также механические и биологические нарушения. Растения реагируют на них ростом, развитием, структурными проявлениями и резистентностью (рис. 12.5).
Рис. 12.5. Факторы местообитания и внешней среды. Проявляющиеся на местности вторичные факторы местообитания оказываются комплексом из первичных факторов местообитания и факторов окружающей среды, которые действуют непосредственно на различные структуры растения и процессы. Они оказывают обоюдное влияние. В дальнейшем можно установить разнообразные обратные связи между растением и средой
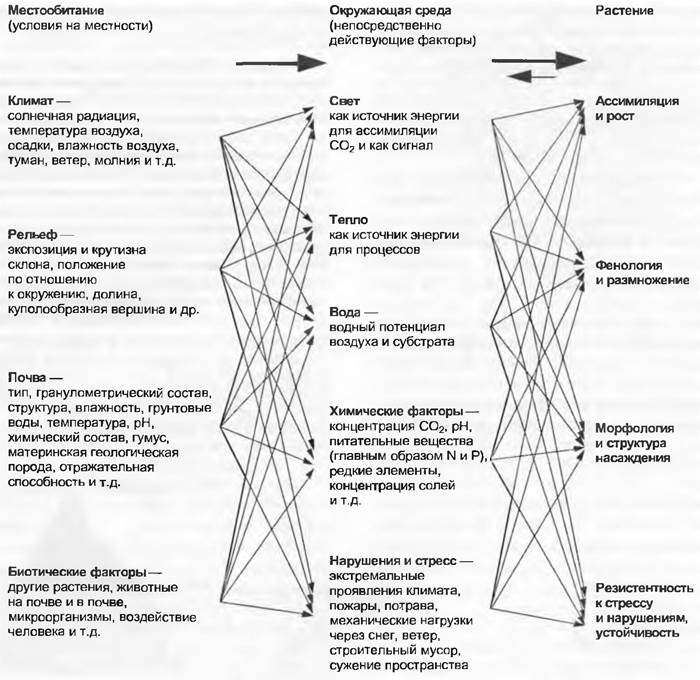
Под местообитанием понимается поверхность, которая характеризуется единообразными факторами. Понятие «местопроизрастание» означает конкретное место, на котором растение фактически растет (= местонахождение). Фактические условия жизни местопроизрастаний в пределах одного и того же местообитания могут существенно варьировать через проявления микроклимата, локального микрорельефа и свойств почвы, через другие виды растений, произрастающих в близком соседстве, и через животных. Твердо установлено, что местопроизрастания не являются случайными, виды растений закономерно существуют в подобных местопроизрастаниях, и там они чувствуют себя «дома», в этом случае говорят об их естественной среде(habitat). Естественная среда — достаточно узкое понятие, применяется к определенному виду растений или группе видов, в то время как местообитание — более широкое понятие, ориентировано на местные условия независимо от того, какие виды там произрастают. Виды растений могут населять также и несколько различных типов естественной среды. Эти три понятия часто смешивают или употребляют в качестве синонимов; однозначного перевода слова «местообитание» на английский язык не существует (большей частью употребляется лапидарное «site»).
12.5.2.2. Климат и микроклимат
Под климатом понимаются усредненное за много лет состояние атмосферы и средний ход метеорологических условий. Погодой обозначают наблюдаемую в данный момент ситуацию. Различные климаты Земли обусловлены прежде всего количеством и распределением по временам года притока тепла и осадков. Эти различия можно представить наглядно в форме климадиаграмм (рис. 12.6).
Рис. 12.6. Климадиаграммы: А — для теплоумеренного климата с континентальным влиянием, с зимними дождями и летними засухами; В — для умеренного климата с океаническим влиянием, с осадками в течение всего года; С — для влажного тропического климата с выраженным периодом дождей и (относительно) сухим периодом. Приведены температуры воздуха в тени на высоте 2 м. Абсцисса: месяцы, ордината: одно деление = 10 °С и соответственно 20 мм осадков а — станция (место); b — высота над уровнем моря; с — число лет наблюдений; d — годовая температура, °С; е — среднегодовое количество осадков, мм; f — средний дневной минимум температур самого холодного месяца; g — абсолютный минимум температур (= самый низкий из измеренных температур); h — средний дневной максимум самого теплого месяца; i — абсолютный максимум температур (= самый высокий из измеренных температур); к — годовой ход среднемесячных температур; I — годовой ход среднемесячного количества осадков; m — сухие периоды (пунктировано); n — влажные периоды (вертикальная штриховка); о — периоды со среднемесячным количеством осадков > 100 мм (масштаб на 1/10 уменьшен); q — «холодное» время года (месяцы со среднедневным минимумом ниже 0°С); r — месяцы с абсолютным минимумом ниже 0°С, т. е. с поздними или ранними заморозками (косая штриховка)
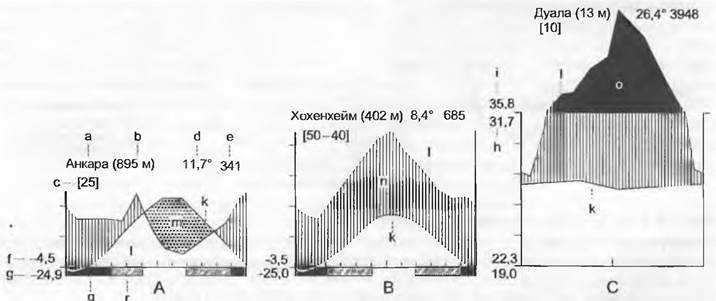
Наглядность климадиаграмм обеспечивается определенным масштабированным в соотношении 2:1 изображением осадков и температуры и равным образом нанесением штриховки, основанным на конкретном опыте. Для биологии ценность климадиаграмм заключается в визуализации динамики климата именно по временам года, вместо средних или суммарных значений. Температуры означают не только тепло, но и потенциальное испарение, на их основании возможно делать выводы о сезонном водном балансе (например, о сухих периодах). Информация дополняется введением данных по суммам осадков, температурным экстремумам и координатам местообитания. В тропиках среднемесячные температуры почти не меняются (в отношении температур суточные колебания больше значат, чем смена времен года), сезонность, если она имеется, выражается только в количестве осадков.
В зависимости от географической широты изменяются солнечное излучение, а с ним и температура, обусловленная температурой сезонность и потенциальная эвапотранспирация (возможное испарение с поверхности почвы и растений при хорошем снабжении водой). Там, где годовая сумма осадков значительно превышает потенциальную эвапотранспирацию, господствует гумидный климат, если осадки заметно ниже потенциальной эвапотранспирации, климат семиаридный или аридный. При этом для растительности важнее распределение осадков по сезонам года, чем их сумма.
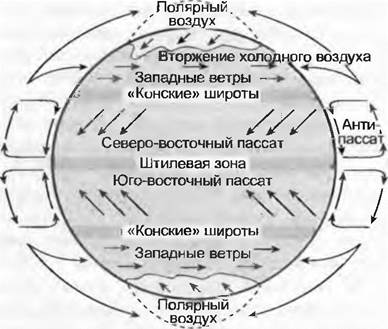
Помимо географической широты климат зависит от глобальной циркуляции воздуха (рис. 12.7) и морских течений. Экваториальная зона низкого давления с восходящими потоками воздуха (конденсация и зенитальные дожди) — влажная, субтропическая зона высокого давления с нисходящими воздушными потоками в континентальной области (так называемые конские широты) — сухая (область пустынь). Благодаря устремляющимся к экватору приземным воздушным течениям возникают пассатные ветры, которые особенно в Южной Азии изменяются, подвергаясь муссонной циркуляции (максимум дождей отмечается летом). В умеренной зоне Северного и Южного полушарий благодаря перемешиванию теплых и холодных воздушных масс образуются циклоны, которые вследствие вращения Земли двигаются в виде господствующих западных ветров на восток (циклонапьные осадки и затяжные дожди характерны для гор краевых областей континента, внутренние области остаются сухими). Воздух полярных областей содержит мало влаги, соответственно, осадков очень мало, но все же больше очень низкой потенциальной эвапотранспирации. Близкие к побережью (морские = океанические) области отличаются меньшими годовыми амплитудами климата, чем отдаленные от побережья (континентальные).
Рис. 12.7. Схема воздушных течений на Земле в горизонтальной и вертикальной проекциях в период равноденствия
Важный фактор для снабжения влагой — сезонное перемещение высоты стояния солнца. Средиземноморская область благодаря этому попадает зимой в зону западных ветров, а летом в субтропическую зону высокого давления. В тропиках Северного полушария максимум осадков сдвигается летом на север, а зимой на юг, вследствие чего, например, в окраинных частях тропиков возникают ясно выраженные влажный и сухой периоды.
Зональный климат сильно преобразовывают и морские течения. Если бы не Гольфстрим, климат в Северной Германии был бы такой же, как на Лабрадоре. Холодное течение Гумбольдта приводит к относительно малому количеству осадков на западном побережье Южной Америки к югу от экватора (экстремальный вариант — пустыня Атакама). Аналогичная ситуация наблюдается и в Юго-Западной Африке (пустыня Намиб). Периодически повторяющаяся аномалия давления и температур в экваториальной части Тихого океана (Эль-Ниньо) возникает примерно раз в пять лет, она вызывается устойчивыми направленными на запад пассатами и сопутствующими им морскими течениями и приводит к наводнениям на западном побережье Южной Америки и к засухам в областях Индомалайзии, где обычно преобладает гумидный климат. В каждом подобном случае экологическое воздействие значительно.
Характерным образом меняется климат и с высотой над уровнем моря. В горах средние температуры падают с увеличением высоты на 0,55 °С через каждые 100 м (причинами этого служат в особенности более слабое нагревание воздуха через поверхность почвы, меньшая плотность воздуха и увеличение теплового излучения). Это приводит к образованию характерных высотных тепловых и растительных поясов. Давление воздуха падает примерно на 10 % через каждые 1 000 м, благодаря чему снижается и парциальное давление СО2 и О2, с падением давления также увеличивается разреженность газов. Все остальные климатические параметры не обнаруживают единообразного высотного изменения. Радиационный климат находится в сильной зависимости от облачности. Во влажных областях есть горы, где количество солнечного излучения сильно падает с высотой (например, Новая Гвинея). В Альпах при безоблачном небе прирост интенсивности радиации и увеличение облачности с высотой держатся почти в одном соотношении, отчего доза (интенсивность излучения за единицу времени) не увеличивается. Ни ветер, ни осадки не придерживаются единообразного общего шаблона, нет у них и определенных изменений, связанных с высотными поясами, хотя имеются градиенты, специфические для данного региона (так, в Альпах и Скалистых горах количество осадков с высотой возрастает, а в некоторых частях Южных Анд — падает). В центральной части горных массивов климат обычно несколько другой (более сухой и теплый), чем в краевых, а потому высотные градиенты также различны (так называемый эффект поднятия воздушных масс).
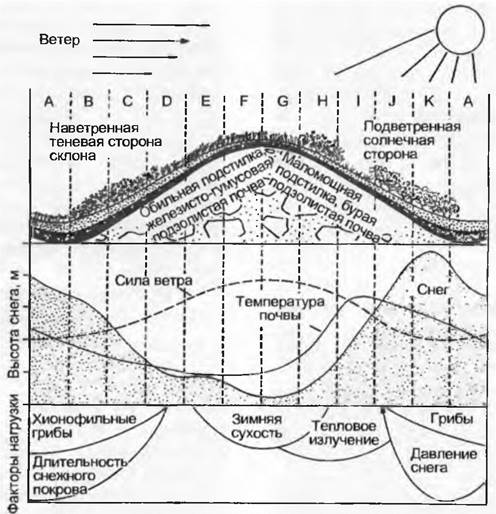
Благодаря рельефу, экспозиции, структуре почвы и растительного покрова реальный климат, в котором живут растения, не всегда соответствует тому, который фиксируется метеостанциями (рис. 12.8). Этот микроклимат может так сильно отличаться от макроклимата, что на уровне конкретного растения периодически исчезают различия между климатическими зонами. Особенно сильно этот эффект проявляется в горах, где низкие, очень сомкнутые растительные сообщества сильно препятствуют теплообмену со свободно циркулирующим атмосферным воздухом, поэтому днем благодаря теплоте излучения на поверхности травостоя могут возникать тропические температуры. Чем ниже и гуще насаждение, тем больше выражено это расхождение в климате (на лужайках оно сильнее, чем в лесу). Тепловая отдача в ясные ночи понижает температуру поверхности насаждения по сравнению с температурой воздуха, что может повлечь за собой непредвиденные повреждения от заморозков. Существенно, что растительный покров сам действует как преобразователь климата. При этом модифицируются все компоненты климата.
Рис. 12.8. Микроклимат в наземном жизненном пространстве. Весенняя ситуация на опушке леса в Нидерландах (полдень после ясной ночи, 3 марта 1976 г.). Пример сильного пространственного разнообразия микроклимата, воздействующего на растения, по сравнению с температурой воздуха (макроклиматом). Интенсивность солнечной радиации различается в зависимости от угла падения на облучаемую поверхность
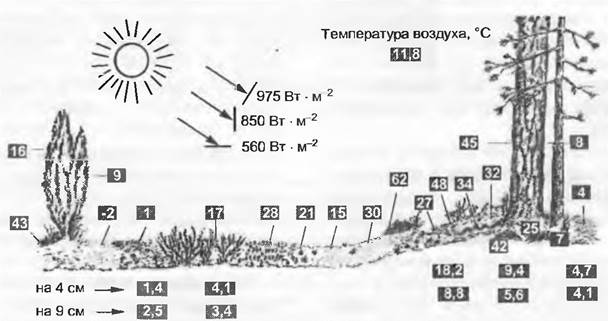
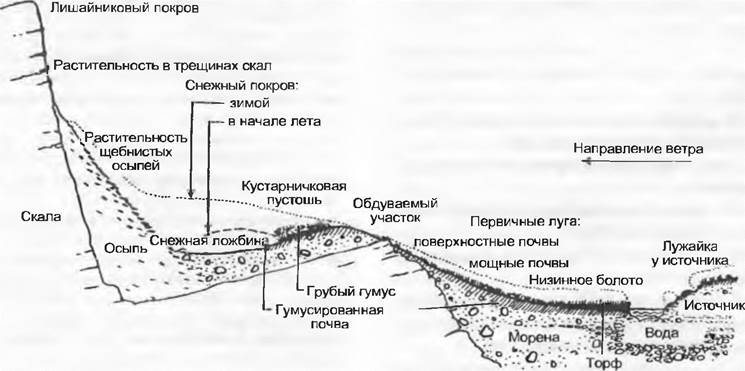
В качестве примера комплексного совместного воздействия рельефа, микроклимата и его вторичных последствий рассмотрим ситуацию на кустарничковой пустоши в Центральных Альпах (рис. 12.9).
Рис. 12.9. Совместное действие рельефа, микроклимата и других модифицированных биологических факторов на примере профиля через альпийскую кустарничковую пустошь в Центральных Альпах (см. также рис. 12.13). Реакции на различающиеся условия жизни в микроместообитаниях при одном и том же макроклимате — характерное чередование растительности: А — лужайка в днище мульды с Soldanella и мхами; В — заросли альпийского рододендрона (Rhododendron ferrugineumy, С — рододендроновые заросли с Vaccinium myrtillus; D — кустарничковая пустошь с доминированием Vaccinium uliginosunr, Е — луазелеуриевая пустошь (Loiseleuria procumbens); F — лишайниковая пустошь с оголенными участками вследствие ветровой эрозии; G — несомкнутая растительность с розеточными и подушковидными растениями и Juncus trifidus, H — кустарничковая пустошь с Arctostaphylos uva-ursi и Vaccinium vitis-idaea; I — пятно, обусловленное перегревом; J — заросли карликового можжевельника (Juniperus communis ssp. alpina) c Calluna vulgaris, Vaccinium vitisidaea, К — рододендроновые заросли с можжевельником
Структура рельефа оказывает влияние на количество солнечной радиации, а также на обусловленное ветрами и крайне неравномерное распределение снега. В мульдах с длительно сохраняющимся снеговым покровом почва увлажнена хорошо, но вегетационный период сильно сокращен, что усиливает возможность поражения ослабленных растений разнообразными хионофильными грибами. На обдуваемых ветрами куполообразных вершинах зимой часто вообще отсутствует снежный покров, поэтому растения здесь подвержены воздействию низких температур и сильной солнечной радиации (возможно морозное иссушение). На солнечных склонах близ поверхности почвы температуры летом особенно высокие, что может привести к оголенным участкам в растительном покрове. На наветренной стороне подобные пустые пространства часто обусловлены ветровой эрозией.
Распределение различных видов растений очень точно отражает условия небольших по площади местообитаний в пределах кустарничковой пустоши. Сравнивая одни виды с другими, можно в общих чертах использовать их как экологические индикаторы (см. 14.3.3) и придать им соответственно эталонированию по фактически измеренным величинам индикаторные значения для определенных факторов местообитания. Тем самым для растительных сообществ и их биотопов можно без трудоемких измерений получить полуколичественные данные для действующих там экологических факторов.
В водных экосистемах температура и доступная радиация также сильно отклоняются от климатических данных метеостанций (рис. 12.10). Весной и летом верхние слои воды преимущественно нагреваются. Благодаря своей малой толщине теплая вода остается летом на поверхности (эпилимнион), в то время как холодный и более плотный гиполимнионлежит под ним (рис. 12.10, В). Охлаждение осенью и зимой совместно с действием ветра способствует перемешиванию слоев, что имеет решающее значение для снабжения кислородом и питательными веществами всей толщи воды.
Рис. 12.10. Климатические отношения в водном жизненном пространстве: А — солнечное излучение; В — распределение температур по слоям во время летних месяцев в эвтрофном озере умеренной зоны (Мондзее, Зальцкаммергут)

12.5.2.3. Почва
Почва формируется в результате совместного воздействия факторов почвообразования: материнской породы, живых организмов, климата и рельефа, которые с течением времени воздействуют на процессы почвообразования. Важнейшие процессы почвообразования — выветривание, переотложение, образование гумуса, минерализация и оструктуривание. В зависимости от топографического положения образуется таким образом живая педосфера, представляющая собой часть биосферы. Эдафон — это совокупность присущих почве живых организмов. Ризосфера охватывает все пространство, занятое корнями, и представляет собой зону соприкосновения растительного покрова и почвы. Почвы — открытые пористые системы, состоящие из твердой, жидкой и газовой фаз, в которых происходит обмен веществом и энергией с литосферой, атмосферой, гидросферой и биосферой. Большое значение, особенно для плодородия почвы, имеет образование глинистых минералов и гумуса. Гумус — органический компонент почвы, возникающий под действием эдафона путем разложения и переработки растительного опада и его перемешивания с минеральными составляющими почвы (рис. 12.11).
Рис. 12.11. Разложение листовой подстилки и образование гумуса (мулль) в буковом лесу на бурой лесной почве: А — листовой опад: В — окончатое выгрызание (ногохвостки и др.), при котором открывается эпидермис (начало поселения бактерий и грибов); С — переход к прогрызанию отверстий; D — прогрызание отверстий и разрушение скелетных жилок (равноногие рачки, многоножки и др.), помет животных; Е — кульминация микробного разложения (бактерии, грибы), дальнейшее поедание сапрофагами (моховые клещи и др.); F — поглощение разложившейся массы, перемешивание ее с минералами и образование глинистогумусного комплекса с помощью детритофагов (дождевые черви и др.); G — состояние после повторного прохода через кишечник (ускоряемого бактериальным разложением!) и формирование зернистой структуры — мулльгумуса (А — Е — почти 1 :3, F — G — около 150х)
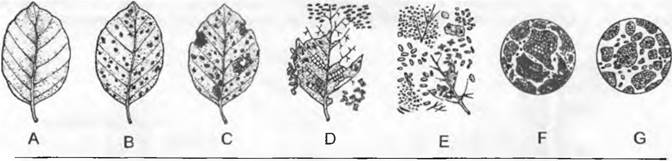
В пересчете на массу и обмен веществ в умеренно-гумидной зоне важнейшими почвенными организмами являются дождевые черви и бактерии. Дождевые черви при живой массе 20 — 80 г • м-2 могут переработать 10 — 40 т мелкозема на 1 га за год. В субтропиках и тропиках с сухим периодом доминирующие животные-деструенты — термиты (в Танзании, например, насчитывали до 200 термитников на I км2); они принимают решающее участие в перемешивании и насыпании почвы (при распаде колоний). В степных областях о постоянном перемешивании почвы заботятся разнообразные роющие грызуны (полевки, земляные белки и др.).
Обогащение органической субстанции в почве ведет в зависимости от насыщенности основаниями к понижению значений pH, в экстремальных случаях до крайних значений около 3. Все факторы, сдерживающие разложение растительного материала, такие, как трудноразлагающаяся хвойная подстилка, неблагоприятные климатические условия или бедные основаниями породы, способствуют образованию грубого гумуса и при этом подкислению почвы. С этим тесно связаны мобилизация и доступность минеральных питательных веществ. Большинство растений с помощью микоризообразующих грибов имеют возможность лучше использовать запасы питательных веществ почвы и при этом увеличивать продукцию биомассы.
Взаимодействия между почвой и растительностью многообразны и очень комплексны, так что простых выводов относительно причинно-следственных связей сделать нельзя. Определяющие факторы структуры этих взаимодействий — исходная подстилающая порода и климат. Растения не только поселяются на определенных почвах, но и влияют на почвообразование. Этот процесс часто ведет к смене видов. Качество и количество подстилки (например, хвои или листвы) существенны для динамики верхнего слоя почвы.
Доступность оснований, которая из-за буферности кислот в почве может сильно варьировать даже в пределах одной ступени pH в какой-либо системе буферности, в экологическом отношении важнее, чем значение pH в данный момент. Важнейшие буферные системы соответствуют следующим значениям pH: карбонатная — 8,6 — 6,2; катионообменная — 5 — 4,2; алюминиевая — менее 4,2; железистая — менее 3,8; кремниевая — всей амплитуде значений. Особенно важно содержание кальция. Помимо значительного буферного воздействия он влияет на физические свойства почвы, например, на оструктуренность (комковатая структура), и тем самым на водный, воздушный и тепловой режимы.
Значения pH почвы в верхнем слое находятся в следующих примерных границах: 2,6 — 4,5 в сильнокислых почвах верховых болот и кустарничковых пустошей; 3,5 — 4,5 в кислых лесных почвах; 4,5 — 6,0 в богатых, умеренно- и слабокислых почвах, смешанных лиственных лесов и пахотных почвах; 5,0 — 6,5 в почвах низинных болот; 6,0 — 7,5 в буковых лесах на карбонатных породах; 6,5 — 8,0 в пойменных лесах; 7,0 — 8,5 в более или менее щелочных почвах степей на известняках, до 10 и более под аридной галофитной растительностью (сильнощелочные содовые почвы — это уже солонцы).
Перекисление почвы объясняется не только образованием гумусовых кислот, но и также выделением кислот корнями и микроорганизмами, диссоциацией угольной кислоты и вымыванием оснований. Поскольку рост растений и активность почвенных организмов находятся под воздействием сезонных колебаний осадков и температуры, значения pH подвержены типовому сезонному ритму. Подщелачивание почвы обусловлено прежде всего обогащением солями сильных оснований и слабых кислот (например, Na2CO3, СаСO3).
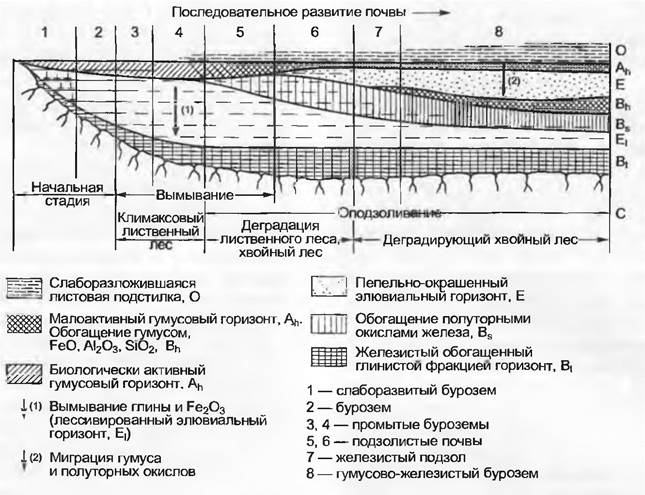
В лесах большая часть органического опала скапливается на поверхности (подстилка). Это направленное поступление опада приводит к очень сильной вертикальной дифференциации почвенного профиля (рис. 12.12). Под травянистой растительностью и в типичных степных почвах органический опад преимущественно поступает при отмирании тонких корней, что наряду с активностью роющих животных и пониженной из-за сухости инфильтрацией объясняет значительно меньшую вертикальную дифференциацию содержания гумуса. Средняя продолжительность пребывания углерода в почве для определенных компонентов исчисляется тысячелетиями, поэтому богатые гумусом почвы имеют весьма значительный возраст, и отсюда их разрушение в определенные периоды времени окончательно и необратимо.
Рис. 12.12. Развитие почвы в атлантической климатической области Европы. Стратификация почвы наглядно показана посредством ее профиля, который изменяется с течением времени. Почвы «созревают», но могут также и деградировать. График показывает последовательность стадий почвообразования
Типизация почвенного субстрата ориентирована: 1) на исходный материал (например, известняк, силикаты); 2) на текстуру, так называемое деление на классы по гранулометрическому составу; 3) на содержание гумуса. Принятые в Германии классы гранулометрического состава перечислены в табл. 12.1.
Таблица 12.1. Классы гранулометрического состава почвы, принятые в Германии
Фракция почвы |
Величина частиц*, мкм |
Скелетная почва |
> 2 000 |
Мелкозем |
< 2000 |
Песок |
63 - 2 000 (50 - 2 000) |
Суглинок |
2 - 63(2 - 50) |
Глина |
< 2 |
* Величина в скобках дана согласно международным классам.
С экологической точки зрения кроме размера частиц почвы особенно важна ее структура, которую определяют одновременно объем пор и классы пористости, имеющие решающее значение для накопления воды (табл. 12.2). Песчаные почвы имеют крупные поры, хорошо проветриваются, быстро дренируются, почему и имеют малую влагоемкость (легкие, теплые почвы); суглинистые и глинистые почвы, напротив, тяжелые, холодные. Соединения коллоидных глинистых минералов и гуминовых веществ (очень сложные гигантские молекулы из многочисленных ароматических циклов частично со связанным азотом и алифатическими боковыми цепями) образуют глинисто-гумусовые комплексы, с отрицательно заряженными поверхностями которых связаны обменные катионы. В ненарушенных почвах с этими агрегатами столь тесно ассоциируются корневые волоски, микоризы и микроорганизмы, что чрезвычайно затрудняет вымывание питательных веществ. Из-за химического связывания азота в частично крайне инертных гуминовых веществах большое количество азота находится в недоступной для растений форме (отношение C/N в гумусе 10/20, в зеленых листьях 30/50, см. 13.6.1), поэтому данные об общем запасе азота в почвах ничего не говорят об обеспечении им растений. Максимальная «загрузка» почв доступными для растений минеральными питательными веществами определяется в значительной степени содержанием глинистых частиц и гумуса. Согласно недавно выведенной биогеохимической модели, продуктивность Земли в конечном счете лимитирована содержанием глинистых веществ в почвах. Отсюда встает вопрос: какое количество углерода во всем мире может быть связано в биосфере?
Таблица 12.2. Классы пористости
Обозначение |
Размер пор, мкм |
Свойства |
Широкие грубые поры |
> 50 |
Вода быстро просачивается |
Узкие грубые поры |
10 - 50 |
Вода легко доступна |
Средние поры |
0,2 - 10 |
Доступность воды от средней до затрудненной |
Тонкие поры |
< 0,2 |
Вода недоступна для растений |
Классификация почв сильно ориентирована на строение почвенного профиля, т. е. на формирование горизонтов, которые обычно обозначаются прописными буквами (см. бокс 12.1). Различают органогенные и минеральные горизонты.
Основные органогенные горизонты:
L — подстилка, в значительной степени не разложившиеся растительные остатки (англ. litter);
F — горизонт ферментации или образования модера (структура тканей различима);
Н — гумусовый горизонт, органические остатки без структуры тканей.
Основные минеральные почвенные горизонты:
А — верхний почвенный горизонт (сильно гумифицированный);
Е — горизонт вымывания (элювиальный горизонт);
В — минеральный горизонт иллювиирования (характеризуется новообразованиями минералов и обогащением);
G — горизонт, находящийся под воздействием грунтовых вод;
S — горизонт, находящийся под воздействием застойных вод;
С — исходная материнская порода, из которой образовалась почва.
В наименованиях типов почв руководствуются такими бросающимися в глаза признаками, как, например, окраска (буроземы, черноземы), или последовательностями легко различаемых горизонтов. Смена типов почв происходит при изменении фактора почвообразования. Если в условиях умеренно-гумидного климата факторы почвообразования не меняются, то из слабоструктурированных примитивных почв (А — С) позднее формируются иллювиальные почвы (буроземы типа А — В — С) или типы почв, сформированные под воздействием процессов переотложе- ния (подзолы А — Е — В — С). У недавно сформировавшихся почв типа А — С (например, рендзин на известняковых породах или ранкеров на силикатах) горизонт А лежит непосредственно на материнской породе. Почвообразующие факторы могут сильно варьировать на небольшом пространстве, что приводит, к мозаике почв (рис. 12.13). Горизонт А считается донором, горизонт В — реципиентом для мобилизуемых в ходе развития почвы веществ.
Рис. 12.13. Возникновение растительной мозаики благодаря различающимся в соответствии с климатом и рельефом свойствам почв. Пример, как и на рис. 12.9, взят из альпийского пояса, поскольку там эта вариабельность почв проявляется на особенно незначительных по площади участках
Обычная последовательность горизонтов в умеренно-холодных хвойных лесах, а также под тундровой растительностью следующая.
Гумус присутствует в форме грубого гумуса (англ. mor), который лежит на минеральных горизонтах почвы и в котором можно различить горизонты L, F и Н различной мощности. В горизонте А, поверх которого лежат различные формы грубого гумуса, происходит перемешивание гумусовых веществ и минеральных составляющих почвы. В холодных и влажных областях за горизонтом А следует более или менее отбеленный и бедный гумусом или вообще лишенный его горизонт вымывания (Е), характерный для подзолов. В этом горизонте глинистые минералы выветрены в наибольшей степени, и продукты их выветривания переоткладывались, как Fe и АI, содержащие гумусовые золи. В экстремальных случаях горизонт Е состоит из одного только кварцевого песка. В подзолистых почвах горизонт В поэтому не только является горизонтом выветривания, но и обнаруживает заметные признаки обогащения веществами, особенно содержащими железогумусовые коллоиды. Данный горизонт при известных условиях может быть настолько пропитан этими веществами (ортштейн), что станет трудно проницаемым для корней. Переход от горизонта В к С большей частью постепенный.
Этот пример последовательности горизонтов в молодых примитивных почвах еще не выражен. Соответственно стадии развития в почвах типа А—С горизонты Е и В отсутствуют, они начинают развиваться только в ходе формирования почвы при соответствующих условиях почвообразования.
В умеренных условиях, примерно в таких, как лиственные леса, гумус часто находится в форме, которую называют модером и которая характеризуется маломощными горизонтами L, F и Н. При очень благоприятных условиях разложения возникает форма гумуса, называемая мулль, в которой горизонты F и Н совершенно отсутствуют. Мулль не представляет собой форму модера, поскольку под слоем подстилки непосредственно лежит минеральный горизонт А, в котором гумусовые вещества и минеральный мелкозем смешаны между собой. В таких условиях подзолы не формируются, а доминируют почвы из серии буроземов, в которых нет полуторных окислов и органометаллических соединений.
В областях с большим количеством осадков и в почвах с затрудненным просачиванием воды образуются переувлажненные так называемые псевдоглеевые почвы, ко
торые в отличие от глеевых почв постоянно переувлажнены грунтовыми водами.
В сухих, теплых континентальных климатах (лесо)степей и прерий формируются преимущественно черные почвы (черноземы). Это очень богатые питательными веществами плодородные почвы типа А — С с мощным, окрашенным в черный цвет гумусовым горизонтом, переходящим непосредственно в минеральный субстрат (часто это лёсс). До глубины проникновения атмосферных осадков в этих почвах имеет место вымывание извести, которая переотлагается в более глубоких горизонтах. В аридных полупустынных и пустынных областях доля гумуса становится все меньше. Здесь образуются, например, окрашенные в каштановый или серый цвет почвы (каштаноземы, аридизоли). Во впадинах таких областей, где скудная влага может скапливаться и просачиваться, это приводит благодаря сильному испарению к восходящему транспорту растворимых солей (например, Nа2СO3, Na2SO4, NaСI, MgSO4 и др.), которые накапливаются и образуют выцветы на поверхности почвы. В таких большей частью сильнощелочных почвах (солончаки) pH может достигать 10. Во влажных тропиках подстилка разлагается очень быстро и формируются бедные питательными веществами латеритные почвы. Из глубоко выветренных минеральных почв вымываются щелочные и щелочноземельные элементы, а также кремневые кислоты, тогда как окислы железа и алюминия наряду с каолинитом накапливаются. Эти почвы едва ли содержат большие количества выветриваемых силикатов (бокс 12.1).
Бокс 12.1. Классификация почв
В связи с большим числом различных типов почв возникает необходимость их группирования в классы почв. Такая классификация облегчает общение между специалистами и позволяет осуществлять картирование почв по сходным особенностям. До сих пор отсутствует признанная международная единая система классификации почв. В настоящее время классификация проводится тремя различными методами, причем основой для них служат или почвообразующие факторы, или почвообразовательные процессы, или свойства самих почв. Классификация, основанная на факторах почвообразования, приводит к делению почв в соответствии с климатическими и растительными зонами (зональные почвы) или с типом исходной породы и топографией (азональные почвы). Типичные зональные почвы — бореальные подзолы, буроземы умеренной зоны, как и тропические латеритные почвы. Типичные азональные почвы — почвы наносных участков, переувлажненные или примитивные почвы.
Классификация по свойствам почв основана на точно определяемых признаках диагностических горизонтов. Эта система развивалась в США в 1960-х годах («Soil Taxonomy») и сейчас относится к наиболее употребляемым методам классификации, хотя требует значительного количества полевых и лабораторных данных. В большинстве европейских стран, напротив, почвы классифицируются с морфогенетической точки зрения, причем одновременно учитываются почвообразовательные процессы и факторы местообитания. Эта система лучше всего подходит для педогенетической интерпретации отдельных почв, она была несколько модифицирована в отдельных странах соответственно их конкретным условиям и потребностям.
Очень широко употребляемая классификационная система — система ФАО-ЮНЕСКО, разработанная в целях картирования почвенного покрова мира. В этой системе классификация проводится по диагностическим признакам почв, равно как и по почвообразовательным процессам и факторам местообитания.
Из-за очень сильно различающихся классификационных подходов строго логичное сопоставление единиц классификации различных систем невозможно и не всегда имеет смысл. Тем не менее можно попытаться показать, как наиболее распространенные типы почв представлены в трех классификационных системах.
«Soil Taxonomy» [таксономия почв] США высшими категориями признает 10 порядков (orders); названия почв составлены в основном из греческих и латинских слогов (табл. А).
Таблица А. Таксономия почв США
Порядок (order) |
Признаки |
Происхождение названия |
Энтисоли |
Неразвитые почвы без выраженных горизонтов |
Англ.«recent» — молодые |
Вертисоли |
Плотные, темные почвы из набухающих глин с сильно выраженной способностью к набуханию и сжатию |
Лат. «venere» — поворачиваться, изменяться |
Инсептисоли |
Слаборазвитые почвы с выраженными горизонтами |
Лат. «inceptum» — начало |
Аридисоли |
Почвы, свойственные сухому климату |
Лат. «aridys» — сухой |
Моллисоли |
Почвы с мощным, темно-окрашенным, рыхлым, богатым гумусом типа мулль горизонтом А |
Лат. «mollis» мягкий, рыхлый |
Сподосоли |
Почвы с альфегумусовым (АI, Fе) горизонтом обогащения, возникшим вследствие иллювиирования и с выраженным горизонтом отбеливания |
гpeч.«spodos» — зола |
Альфисоли |
Почвы с глинистым горизонтом обогащения, но с умеренным силикатным выветриванием |
«Pedalfer» — староамериканское название почв, лишенных карбонатов |
Ультисоли |
Почвы с глинистым горизонтом обогащения, слабой насыщенностью основаниями; свойственны областям с годовой температурой > 8 °С |
Лат. «ultimus» — последний |
Оксисоли |
Почвы внутренних тропических районов, богатые полуторными окислами, сильно выветренные |
«Oxis» |
Гистосоли |
Болотные и другие почвы с мощным слоем органического вещества |
гpeч. «histos» — ткань |
Таблица В. Почвенная классификация ФАО
Группа |
Признаки |
Происхождение названия |
Флювисоли |
Почвы пойм и побережий со слабо выраженной дифференциацией профиля |
Лат.«fluvius» — река |
Глейсоли |
Почвы с сильно выраженными гидроморфными признаками |
Gley — влажные, тяжелые почвы |
Регосоли |
Примитивные почвы из рыхлых горных пород на твердых кристаллических породах |
Греч. «regos» — покров |
Литосоли |
Слаборазвитые поверхностные почвы преимущественно из твердых кристаллических пород |
Греч. «lithos» — камень |
Андосоли |
Темные почвы из вулканических пеплов |
Яп. «аn do» — черные почвы |
Вертисоли |
Почвы с сильно выраженными явлениями набухания и сжатия из-за высокого содержания глин |
Лат. «vertere» — поворачиваться, изменяться |
Камбисоли |
Почвы с изменяющимися в результате выветривания окраской, структурой и текстурой |
Лат. «cambiare» — меняться |
Кальцисоли |
Почвы, обогащенные известью на глубине менее 1,25 м |
От «calcium» |
Солончаки |
Почвы, обогащенные свободными солями (NaСI, гипс и др.) |
Русское название засоленных почв |
Солонцы |
Почвы с высокой сорбцией Na |
Русское название щелочных почв |
Черноземы |
Черные земли степей |
Русск. «черный» |
Лювисоли |
Почвы с иллювиированием глин и высокой насыщенностью основаниями |
Греч.«Iоuо» — вымывать |
Подзолы |
Почвы с сильноотбеленным горизонтом вымывания |
Русск. «под золой» |
Акрисоли |
Кислые почвы с низкой насыщенностью основаниями |
Лат. «acris» — кислый |
Нитисоли |
Почвы с иллювиированием глин и хорошо различимыми глинистыми куланами |
Лат. «nitidus» — блестящий |
Феррасоли |
Почвы с высоким содержанием полуторных окислов |
Лат. «ferrum» — железо, «al» — алюминий |
Гистосоли |
Органические почвы, болотные почвы |
Греч. «histos» — ткань |
Антросоли |
Почвы, возникшие под воздействием человека и (или) существенно им преобразованные |
Греч. «anthropos» — человек |
Почвенная классификация ФАО содержит 28 основных групп почв (табл. В), в ней перечислены группы, имеющие особенно важное значение для Центральной Европы.
В табл. С представлен фрагмент классификационной системы, применяемой в Германии, как пример систем, используемых в большинстве стран Европы.
Таблица С. Система классификации почв Германии (фрагмент)
Тип почвы |
Свойства |
Наземные почвы |
|
Примитивные наземные почвы |
См. регосоли и литосоли ФАО |
Почвы типа А — С: |
Почвы без суглинистой подпочвы |
ранкеры |
На твердых горных породах, некарбонатных или бедных карбонатами |
регосоли |
На рыхлых горных породах, некарбонатных или бедных карбонатами |
рендзины |
На карбонатных или гипсовых твердых горных породах |
парарендзины |
На мергелях |
степные почвы |
См. черноземы ФАО |
Буроземы |
Типичные буроземы без иллювиирования глин или парабуроземы с иллювиированием глин |
Подзолы |
См. классификацию ФАО |
«Кальциевые земли» |
Пластичные почвы из карбонатных пород сухих и теплых местообитаний; террафуска и терра росса |
Переувлажненные почвы |
Стагносоли: псевдоглеевые и стагноглеевые почвы |
Антропогенные почвы |
Коллювий или коллювисоли, хортисоли, ригосоли |
Полуназемные почвы (с грунтовыми водами) |
|
Пойменные почвы |
См. флювисоли ФАО |
Глеевые почвы |
См. глейсоли ФАО (типичные глеевые, заболоченные глеевые, болотные глеевые) |
Марши |
Илистые почвы |
Болота |
Почвы с торфяным слоем свыше 3 дм |