БОТАНІКА З ОСНОВАМИ ГІДРОБОТАНІКИ (ВОДНІ РОСЛИНИ УКРАЇНИ) - Б.Є. Якубенко - 2011
VI. СИСТЕМА РОСЛИННОГО СВІТУ
НИЖЧІ СПОРОВІ РОСЛИНИ
ЗАГАЛЬНА ХАРАКТЕРИСТИКА ТА ОСОБЛИВОСТІ
Відділ зелені водорості - Chlorophyta
Еукаріотичні, вільно плаваючі чи прикріплені, одноклітинні, колоніальні та багатоклітинні, мікро- та макроскопічні (понад 8 м завдовжки) прісноводні, і гіпергалінні, морські та грунтові і аерофітні, кріофільні та термофільні, монадного, кокоїдного, сарциноїдного, нитчастого й гетеротрихального, інколи з паренхіматозно-платівчатим таломом із листо-, стебло- й ризоїдоподібними утворами, а також неклітинного сифонального та сифонокладального типу структури слані.
Одна з найчисленних груп водоростей (понад 20 тис. видів), що нараховує у флорі України понад 1300 видів, представлених близько 1500 таксонами видового та внутрішньовидового рангу.
Фотосинтетичний апарат аналогічний за складом судинним рослинам й містить хлорофіли а та b (як вийняток прообраз хлорофілу с), каротини ( α-, β-, γ- та ε-каротин), ксантофіли лютеїнового ряду (лютеїн, зеаксантин, неоксантин, віолаксантин, антераксантин). Водночас, для окремих груп (класів) властиві специфічні ксантофіли - празиноксантин, сифонеїн, сифоноксантин, лороксантин.
Зелене забарвлення представників є типовим, завдяки переважанню хлорофілів. Проте, за певних умов (вік, стадія росту, життєвий стан - спочиваючі клітини, гіпергалінні умови) спостерігається накопичення каротину, олії, забарвленої каротинами та ксантофілами (гематохром) і формування жовто-оранжевого чи червоного забарвлення.
Хлоропласти різноманітної форми, від одного до багаточисельних, пристінні чи центральні осьові, первинно-симбіотичні, мають типово рослинну будову, як у судинних рослин. Вони покриті двомебранною оболонкою, а тилакоїди згруповані у ламели по 2-6 або утворюють грани, оперізуючі тилакоїди відсутні.
Нерідко хлоропласт містить піреноїд з крохмальною обгорткою чи без неї - «голий», інколи він відсутній. Продукт асиміляції - крохмаль, що відкладається в стромі хлоропласта та навколо піреноїда, а за межами пластид - олія, лейкозин, зрідка інулін. Для клітин багатьох представників характерна наявність вакуолі з клітинним соком (рис. 24).
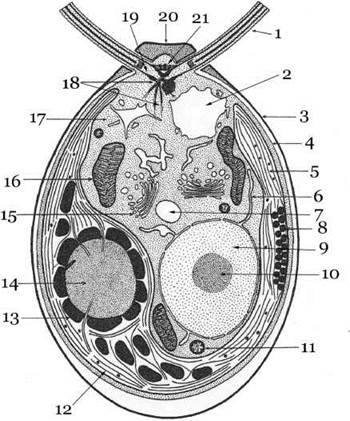
Рис. 24. Схематичний план будови клітин зелених водоростей:
1 - джгутик, 2 - пульсуюча вакуоля, 3 - клітинна оболонка, 4 - плазмалема,
5 - хлоропласт, 6 - ендоплазматичний ретикулум, 7 - вакуоля, 8 - вічко (стигма), 9 - ядро, 10 - ядерце,
11 - краплина олії, 12 - тилакоїди, що зібраніу грани, 13 - крохмальна обгортка (зерна крохмалю) піреноїда, 14 - піреноїд, 15 - апарат Гольджі, 16 - мітохондрія, 17 - цитоплазма, 18 - мікротубулярні корені джгутиків, 19 - базальне тіло джгутиків,
20 - папіла, 21 - фібрилярне сполучне волокно базальних тіл (за ЕМ, 1976, 1983)
Більшість монадних водоростей мають фоторецепторний комплекс із стигми - затінюючого утвору фоторецептора, регулятора освітлення, яка сформована каротиновмісними глобулами та ущільненим шаром плазмалеми - фоторецептора, що знаходиться безпосередньо під оболонкою хлоропласта.
Монадні форми мають 2-4 джгутики, інколи один чи численні (до 120), рівні за довжиною - ізоконтні та однотипні за будовою - ізоморфні, найчастіше - гладенькі, зрідка з мастигонемами, інколи з органічними субмікроскопічними лусочками. У ділянці виходу джгутиків із клітини (так звана «перехідна зона») знаходиться специфічна зірчаста структура, яка властива цим водоростям і судинним рослинам. Характер розміщення «джгутикових коренів», особливості організації кореневої системи джгутиків є різнотипними та використовуються як діагностичні ознаки на рівні класів. Джгутики зорієнтовані один до одного супротивно на одній лінії (конфігурація «12/6»), за годинниковою стрілкою (конфігурація «1/7»), а також проти годинникової стрілки (конфігурація «11/5») (рис. 23). Клітини більшості зелених водоростей мають целюлозно-пектинову оболонку (рис. 24), але деякі позбавлені її та оснащені лише ущільненим шаром плазмалеми, інколи покритої субмікроскопічними органічними лусочками.
Оболонка клітин у більшості видів - суцільна, в окремих представників - сформована з окремих частин (2-9), гладенька або орнаментована різноманітними утворами - бородавками, ребрами, зубчиками, шипиками, шипами, щетинками, волосками тощо. Клітини містять справжні вакуолі з клітинним соком, а деякі - 2, інколи декілька скоротливих вакуолей.
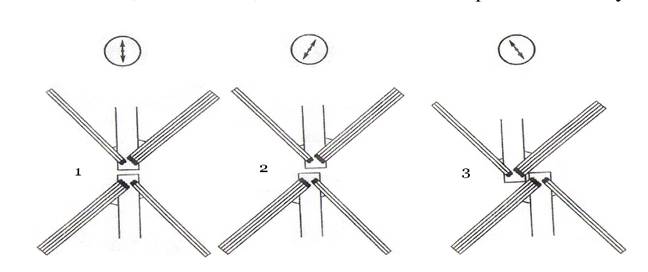
Рис. 25. Схематичний план розміщення мікротубулярних коренів джгутиків зелених водоростей:
1 - хрестоподібне, симетричне, в одній площині (конфігурація «12/6»), 2 - 3 - асиметричне розміщення:
2 - конфігурація «1/7» (за годинниковою стрілкою) та 3 - конфігурація «11/5» (проти годинниковоїстрілки)
Мітохондрії з пластинчастими кристами. Ядерний комплекс має типово еукаріотичну будову. Оболонка ядра в одних представників зберігається протягом мітотичного поділу, в інших вона руйнується надзвичайно рано, а у требуксієвих - зберігається протягом поділу, проте руйнується на полюсах. Ядерце зникає в період мітотичного поділу. Мітоз різного типу - закритий, напівзакритий або відкритий, характерний для представників конкретного класу. Центріолі присутні у нерухомих форм, та відсутні у клітинах монадних представників.
Цитокінез більшості представників відділу зелених водоростей здійснюється завдяки кільцевій борозні. Водночас, у водоростей із розвинутою клітинною оболонкою відзначене також формування клітинної платівки (цитотомія за участю кільцевої борозни та клітинної платівки, яка завжди обумовлює наростання талому), що є особливістю цього відділу.
Для зелених водоростей характені різні типи розмноження - вегетативне, нестатеве та статеве. Вегетативне розмноження одноклітинних відбувається поділом клітини на дві (схізотомія), а у ценобіальних, колоніальних і багатоклітинних - фрагментацією талому (схізогонія, наслідком якої є споруляція чи утворення та ріст багатоклітинних сарциноподібних агрегатів або ж спричинює ріст таломів у нитчастих та гетеротрихальних форм) і формування акінет. їм властиві основні способи нестатевого розмноження (завдяки зооспорам - з оболонкою чи без неї, аплано-, авто-, гіпно-, гемізоо- та геміавто- спорам) і статевого процесу (холо-, ізо-, гетеро-, оогамія), що відомі у еукаріо- тичних водоростей, з поступовою редукцією рухливих репродуктивних стадій. Все більшого значення набуває вегетативний і статевий тип розмноження.
Життєві цикли зелених водоростей різнотипні. Більшість представників (еугамних) характеризуються гаплофазними, гаплодиплофазними та диплофазними циклами зі зиготичною, гаметичною, споричною і соматичною редукцією, з ізо- та гетероморфною зміною поколінь, а серед агамних або апогамних форм відзначені життєві цикли за типом цикломорфозу.
Класифікаційна система цього відділу знаходиться в стадії доопрацювання. У 70-80 х роках XX ст. на заміну класично- морфологічним підходам (за типом морфологічної структури слані та особливостям статевого процесу) використано ультраструктурні (характер будови джгутикового апарату, симетрія розміщення джгутиків і тип організації й будови джгутикових коренів, особливості поділу клітини та ядра, тонка будова клітинних покривів тощо), а за їхнею значущістю проведено новий розподіл відділу на класи.
Згодом молекулярно-філогенетичні результати досліджень (за аналізом нуклеотидних послідовностей ядерного гену, що кодує 5S, 18S і 26S субодиниці рибосомальної РНК, та інших хлоропластних і мітохондріальних генів, наприклад, генів фактору подовження тубуліну, цитохрому с, актину, RUBISCO тощо, підтвердили доцільність і обгрунтованість класифікаційних змін серед відділу і сприяли встановленню нових класів та перерозподілу таксономічних груп зелених водоростей, уточнення яких триває і нині.
Найвизначним і достатньо аргументованим за сукупністю класичних та нових молекулярно-біологічних результатів дослідження є поділ відділу Chlorophyta на класи: Prasinophyceae, Chlorophyceae, Oedogoniophyceae, Trebouxiophyceae, Ulvophyceae та Siphonophyceae (Bryopsidophyceae). Основні характеристики та представництво деяких із цих класів (які передбачені типовою програмою) ми розглянемо.
Зелені водорості є дуже поширеною групою рослин, що заселили різноманітні біотопи від ефемерних водойм до морських акваторій, ґрунту, льоду, термальних джерел та аерофітону. Певні їхні групи (класи) приурочені до різних екологічних умов. Найрізноманітніше представлені Chlorophyceae та Trebouxiophyceae (частково) у прісних водоймах, а Ulvophyceae та Siphonophyceae - у морях, Trebouxiophyceae, Chlorophyceae (частково) - в аерофітних та ґрунтових умовах.
Масовий розвиток окремих представників викликає «цвітіння» водойм та накопичення біомаси до декількох кілограмів у кубічному метрі (Dunaliella salina), а в ґрунтах чи у разі «цвітіння» ґрунту їхня маса досягає до 50 кг/га, інколи 200 кг/га. Нерідко зелені водорості є збудниками зеленого або червоного «цвітіння» снігу (вольвоксові та хлорококові), кори дерев та поверхні різних матеріалів, як натуральних, так і промислових (хлорелові, требуксієві). Відомі також симбіотичні представники, які формують відносини з іншими рослинами (водоростями та судинними формами), грибами та грибоподібними організмами (лишайниками), а також тваринами (кишковопорожнинні, інфузорії). Водночас, серед них відзначені паразитичні представники водоростей-макрофітів, судинних рослин і хребетних тварин.
Визнана їхня роль як об’єктів біотехнології, промислових виробництв і аквакультури, біоіндикаторів стану довкілля та водного середовища, тест- об’єктів забруднення повітря, продуцентів ^-каротину (Dunaliella salina, Haematococcus pluvialis), ліпідних комплексів як основи біодизелю (Botryococcus braunii), гліцерину та вуглеводневих сполук (Dunaliella), а також як біодобавки до корму худобі, птахам та в їжу людини (Chlorella vulgaris, Scenedesmus sp., Ulva sp., Enteromorpha sp., Monostroma sp.). Відзначена також їхня роль як ґрунтозахисного та водоутримуючого фактору в ґрунті (Chlamydomonas).