ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 2. ЕМБРІОЛОГІЯ
2.8 Ембріональний розвиток ссавців
Найбільш примітивні з сучасних ссавців — качконіс, єхидна, характеризуються овоцитами, які містять велику кількість жовтка, належать до телолецитального типу, зигота дробиться за меробластичним методом — дискоїдально. їх ембріогенез подібний до розвитку птахів.
Зигота качконоса розвивається поза материнським організмом. У єхидни зигота розвивається в складці шкіри на черевній поверхні тіла, але живлення від матері зародок не одержує.
У зиготі сумчастих жовтка мало, в процесі дроблення він навіть відторгується (сумчаста куниця, опосум).
У деяких сумчастих розвивається примітивна плацента, яка функціонує нетривалий час. Зародок цих представників ссавців народжується недорозвиненим, занадто маленьким (1-2 см) і доношується матір'ю в сумці, де встановлюється зв'язок стравоходу зародка з соском матері, її молоко час від часу впорскується в стравохід скороченням м’язів молочної залози.
Ссавці різняться між собою не лише будовою яйцеклітин та умовами ембріонального розвитку, а й іншими ознаками: дроблення зиготи, початком імплантації зародка, рівнем зрілості при народженні, тривалістю внутрішньоутробного розвитку тощо.
Вторинна поява в процесі еволюції оліголецитальних яйцеклітин у вищих ссавців зумовлена переходом їх зародка на живлення за рахунок організму матері. Зародкові оболонки і маткові тканини вступають в тісну взаємодію, утворюється плацента, яка безпосередньо постачає все необхідне для розвитку і виводить продукти обміну. З розвитком плаценти зникає необхідність у жовтку, однак у вищих ссавців жовтковий мішок утворюється, хоча і не виконує функції, пов'язаної з живленням.
Дроблення (рис. 34). В результаті запліднення яйцеклітина переходить із інгібірувального стану в стан надзвичайно інтенсивного метаболізму, що характеризується різким підвищенням дихальної і синтетичної активності. Дроблення оліголецитальних зигот ссавців повне, асинхронне відбувається відносно повільно. В той час, коли зигота рухається у матковій трубі, утворюється зародок, який складається із непарної кількості бластомерів, що мають вигляд купки клітин. Цю стадію називають морулою (від лат. morula - тута). В кінці дроблення зародок складається із купки темних великих клітин — зародкового вузлика (ембріобласта) і поверхневого шару світлих дрібних клітин — трофобладуа.
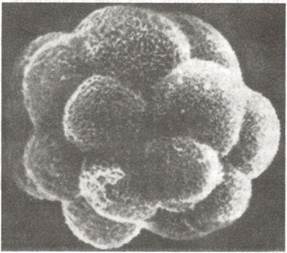
Рис. 34. Електронна мікрофотографія чотириденного зародка хом’яка х 700; на поверхні клітини трофобласта
За рахунок ембріобласта розвивається тіло зародка, а трофобласт виконує трофічну функцію, за його участю забезпечується зв’язок із стінкою матки. У трофобласті розвиваються численні вирости — первинні ворсинки, утворені тільки трофобластичними розростаннями, в які потім вростає позазародкова мезодерма (мезенхіма) і утворює там кровоносні судини. Ворсинки трофобласта з кровоносними судинами є вторинними, вони мають мезодермальну основу, а трофобласт з вторинними ворсинками називають хоріоном. Третинні ворсинки містять кровоносні судини. Якщо тварина багатоплідна (свині, собаки, кролі тощо), то у маткові труби надходять одночасно кілька яйцеклітин. Під час дроблення зародок переміщується у матковій трубі у напрямку матки (рис. 35).
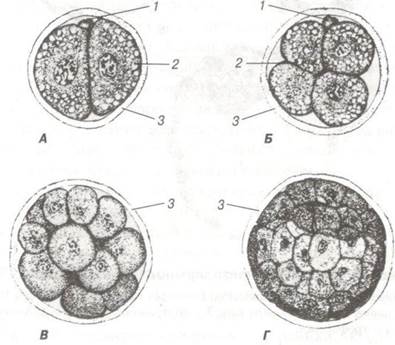
Рис. 35. Дроблення зиготи свині х 400:
А — стадія двох бластомерів; Б — стадія чотирьох бластомерів; В — морула;
Г — бластула, світліша центральна ділянка, в якій формується бластоцель шляхом перерозподілу клітин:
1- полярне тільце; 2— жирові вакуолі; 3 — прозора оболонка
У різних представників вищих ссавців період дроблення зиготи буває різної тривалості. Так, у кролематок через три доби після парування зародок знаходиться на стадії бластули.
Дроблення зигот у свині подібне з процесом у кролематок. В овець дроблення зиготи закінчується за 3-4 доби, у корів — за 6 діб, у приматів дроблення триває з 1 по 6 добу життя зародка. Спочатку трофобласт всмоктує секрет залоз матки «маточне молоко», зародок збільшується в об'ємі і перетворюється у бластодермічний міхурець, який називається бластоцистою. Під час дроблення і утворення бластоцисти прозора зона зиготи ще зберігається. В подальшому за допомогою ворсинок, що утворилися на поверхні трофобласта, бластоциста імплантується (від лат. іш — проникнення в, plantatis — посадка), тобто прикріплюється до слизової оболонки матки. Встановлення тісного взаємозв'язку із слизовою оболонкою матки — перший критичний період у розвитку зародка. У ссавців різних видів стінка трофобласта неодночасно вступає в тісний контакт з слизовою оболонкою матки. Так, у великої рогатої худоби імплантація настає на 17 добу, у кобили — на 63-70 добу, у макаки — на 9 день після запліднення, у людини — на 7 добу. В цей час під впливом ферментних систем трофобласта руйнується оболонка запліднення, яка до цього часу оточувала зародок і бластоциста вступає в контакт із слизовою оболонкою матки (рис. 36).
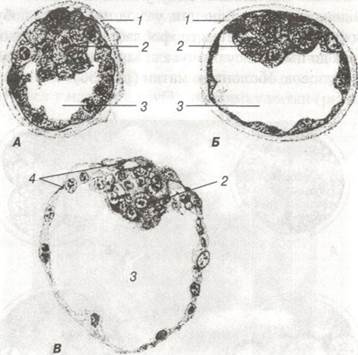
Рис. 36. Три стадії бластодермного міхурця свині х 375:
А — через 114 год. після парування; Б — через 145,75 год.; В — через 164 год. після парування:
1 — прозора зона; 2 — внутрішня клітинна маса; 3 — бластодель; 4 — трофобласт.
Клітини зародкового вузлика формують зародковий диск, аналогічний такому у птахів. У центрі зародкового диска утворюється зародковий щиток, із його матеріалу розвивається тіло зародка, а із останньої частини — провізорні органи. Процес гаструляції у сумчастих і плацентарних ссавців принципово такий, як у плазунів і птахів.
Шляхом деномінації зародкового диска утворюється ектодерма і ентодерма. Зародкові листки, що утворилися з матеріалу зародкового щитка, називаються зародковими, а ті, що виникли із позазародкової зони бластодиска — позазародковими.
Розвиток зародка ссавців пов'язаний з утворенням первинної смужки, яка подібна до первинної смужки птахів. Від переднього кінця первинної смужки, клітини переміщуються і утворюють головний відросток. Останній разом із первинною смужкою дає початок мезодермі, головний відросток і його мезодерма відповідають хордомезодермі нижчих тварин. Диференціювання зародкових листків відбувається так, як у інших тварин. Ці ознаки є найдавнішими і відображають еволюцію, якій піддавалися у своєму розвитку ссавці.
Необхідно зазначити, що однією з особливостей розвитку ссавців при ізолецитальному типі овоцитів і голобластичному дробленні зиготи вважають утворення тимчасових органів. Друга особливість розвитку ссавців — це раннє відокремлений зародкової частини від позазародкової. На початку дроблення формується трофобласт, за допомогою якого зародок одержує поживні речовини з материнського організму.
Після утворення зародкових листків трофобласт, що знаходиться над зародком, тимчасово розвивається і до утворення амніона знаходиться на поверхні. Внаслідок розростання ектодерми зародкового листка останній починає вигинатися, утворює амніотичні складки, які потім зростаються в загальний амніон. Зовнішні листки складок, які прямо переходять у трофобласт, також з'єднуються над зародком в одну оболонку, яка нагадує серозу птахів. До цієї оболонки зсередини підростає мезодерма паріетального листка спланхнотома, в якій розвиваються кровоносні судини. Так виникає хоріон ( від лат. chorion — шкіра). Слід зазначити, що у ссавців (як і у птахів) розвитку зародкових оболонок передує утворення тулубової та амніотичної складок. Тулубова складка піднімає зародок над жовтковим мішком і відокремлює його зародкову частину від незародкової, а зародкова ентодерма замикається в первинну кишку, проте остання залишається з'єднаною з жовтковим мішком вузьким жовтковим протоком.
Жовтковий мішок, як і у птахів, утворюється із позазародкової ентодерми та вісцерального листка мезодерми. На відміну від птахів, він містить не жовток, а білкову рідину. В стінці жовткового мішка утворюються кровоносні судини, він виконує кровотворну та трофічну функції.
Амніон (водна оболонка) розвивається з внутрішньої частини амніотичної складки, а хоріон — із зовнішньої. Амніон у ссавців виконує функцію, подібну до такої у птахів. Хоріон гомологічний серозній оболонці птахів. На його поверхні утворюються ворсинки, які вростають в стінку матки. Ця зона дуже потовщена, у ній багато кровоносних судин і її називають плацентою. Надходження поживних речовин у кров зародка та вихід із неї відбувається дифузним шляхом або за допомогою активного перенесення, тобто з витратами на цей процес енергії. Кров матері не змішується з кров’ю зародка.
Ентодерма задньої частини кишечника утворює вип'ячування — алантоїс. Так, як у ссавців сечовина із тіла зародка виділяється через плаценту в кров матері, але, на відміну від птахів, алантоїс у них не є сечовим мішком, однак його проксимальна частина пізніше бере участь в утворенні сечового міхура. Його мезодерма дає початок кровоносним судинам, які підростають до хоріона.
Поясна плацента характерна для хижаків. Ворсинки хоріона знаходяться в середній частині плодового міхура і утворюють на його поверхні ніби поясок (рис. 38).
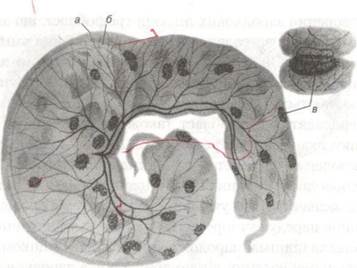
Рис. 37. Котиледонна плацента жуйних: а — хоріон, б — амніон, в — котиледон

Рис. 38. Поясковидна плацента хижаків
Оскільки зародок починає живитися за рахунок материнського організму, у багатьох ссавців жовтковий мішок не відіграє значної ролі, як у птахів, хоча жовтковий мішок утворюється, що характерно для ембріонів свиней, кролів, коней. В жовтковому мішку утворюються кровоносні судини, які забезпечують зародок поживними речовинами матки. Пізніше, коли алантоїс досягає значного розвитку і набуває функції жовткового мішка, останній поступово редукується.
Амніон виникає у вигляді шару соматоплеври, яка утворює навколо зародка складки таким чином, як це було описано для курячого зародка, проте у ссавців частина трофобласта, що знаходиться над зародковим диском, тимчасово розривається і зародок до утворення амніона залишається відкритим. Коли амніон замикається, він заповнюється рідиною,яку продукує. В амніотичній рідині зародок знаходиться у зваженому стані. При цьому зародок зазнає однакового тиску зовні і зсередини, що запобігає механічним пошкодженням. Крім того, амніотична оболонка забезпечує розвиток плода в оптимальному за складом електролітів та інших речовин водному середовищі. Амніон прикріплюється до тіла зародка у тому місці, де на черевній поверхні в ділянці жовткового стебельця стінка тіла відкрита, в подальшому тут утворюється пуповина.
Хоріоном є зовнішній шар бластоцисти ссавців. На різних стадіях розвитку його називають по-різному: на стадії внутрішньої клітинної маси — трофобластом, а після утворення ентодерми та мезодерми зовнішній шар бластоцисти продовжується в ектодерму зародка, й тому має трофоектодерму, оскільки частина цього шару нагромаджує поживні речовини матки. Після того, як мезодерма розшарується і її паріетальний шар з’єднується з ектодермою, цей шар, який став тепер позазародковою соматоплеврою, називають хоріоном. Про утворення алантоїса та його значення було описано вище.