ЦИТОЛОГІЯ, ГІСТОЛОГІЯ, ЕМБРІОЛОГІЯ - 2008
Розділ 3. ЗАГАЛЬНА ГІСТОЛОГІЯ
3.3 Сполучна тканина
3.3.2 Кровотворення (гемоцитопоез)
Гемоцитпопоез (від гр. haima — кров, kythos — клітина, poesis — творення) — складний послідовний процес утворення клітин, у результаті якого досягається постійність кількісного та якісного складу формених елементів крові. Розрізняють кровотворення після народження протягом усього життя тварини та кровотворення у період ембріонального розвитку.
Після народження тварин кровотворення відбувається у мієлоїдній (гр. mielos — мозок) тканині червоного кісткового мозку плоских та епіфізів трубчастих кісток, де відбувається розвиток стовбурових клітин і усіх формених елементів крові — еритроцитів, лейкоцитів, кров'яних пластинок. Утворення еритроцитів, лейкоцитів і кров'яних пластинок називають терміном мієлопоез (від гр. myelos — мозок).
Кровотворна тканина тимуса, селезінки, лімфатичних вузлів, слизових оболонок внутрішніх порожнинних органів називається лімфоїдною. У ній утворюються лімфоцити і плазмоцити, а також здійснюється диференціація і розмноження Т- та В-лімфоцитів. Цей процес називають лімфоцитопоезом.
У птахів лімфоїдна тканина знаходиться у фабрицієвій бурсі, слизових оболонках органів травлення, печінці, легенях, лімфоїдному утворенні ока тощо. Між органами мієлоїдної та лімфоїдної тканин існує тісна взаємодія, що відбувається шляхом міграції і рециркуляції клітин крові, нейрогуморальної регуляції кровотворення і розвитку крові.
Відповідно до сучасної унітарної теорії кровотворення, усі зрілі клітини крові походять з однієї загальної родоначальної клітини, яку називають стовбуровою кровотворною клітиною (СКК). Її диференціювання у різні види клітин крові зумовлюється мікрооточенням ретикулярної тканини органів мієлоїдного та лімфоїдного кровотворення і дією специфічних гемопоетинів. Основи унітарної теорії кровотворення були закладені відомим російським гістологом О.О.Максимовим у 1927р. Такою клітиною є малий лімфоцит, який зберігає здатність перетворюватися у клітини крові. Ця теорія значно поглиблена подальшими різноманітними новими методами дослідження — клональними (групи клітин, що утворюються в селезінці з однієї клітини), трансплантації, радіобіологічними, генетичними, імунологічними, електронномікроскопічними. Було встановлено, що стовбурові клітини крові (СКК) виникають в ембріональному періоді у жовтковому
мішку 7-8 денних ембріонів мишей і потім розселяються у всій кровотворній системі.
У дорослому організмі найбільша кількість СКК міститься у червоному кістковому мозку (на 100000 клітин кісткового мозку припадає 50 СКК). За допомогою світлового та електронного мікроскопів встановлено, що за своєю будовою СКК подібні до малого лімфоцита.
За даними І.Л.Черткова і А.І.Воробйова (1981), у сучасній схемі кровотворення розрізняють шість класів клітин:
I. — Поліпотентні клітини — попередники СКК;
II. — Частково детерміновані клітини — попередники, потенції цих клітин частково обмежені, вони здатні до утворення мієлопоезу і лімфопоезу;
III. — Уніпотентні клітини — попередники, здатні розвиватися в одному напрямі, під впливом гемопоетинів;
IV. — Бласти, морфологічно розпізнавані проліферуючі клітини — попередники, здатні до мітозу;
V. — Дозріваючі клітини;
VI. — Зрілі клітини, здатні до виходу в кров.
Еритроцитопоез — це розвиток еритроцитів, який відбувається у червоному кістковому мозку із стовбурової кровотворної клітини. Ця клітина ділиться, диференціюється у клітину — попередника мієлопоезу. Її ще позначають як колонієутворюючу одиницю гранулоцитів, еритроцитів, моноцитів, мегакаріоцитів (КУО-ГЕММ), або напівстов- бурову клітину (НСК). З цієї клітини утворюються більш детерміновані попередники двох видів: КУО-НГЕ (колонієутворююча одиниця нейтрофільних гранулоцитів та еритроцитів) і КУО-МГЦЕ (колонієутворююча одиниця мегакаріоцитів та еритроцитів). Таким чином, наступна стадія розвитку еритроцитів — їх уніпотентний попередник КУО-Е розвиваються лише в напрямі еритроцитів. Може утворитися двома шляхами — з КУО-ГНЕ або КУО-МГЦЕ. Уніпотентну клітину еритроцитопоезу називають ще еритроцитопоетинчутливою (ЕЧК), оскільки її подальша диференціація індукується гормоном еритропоетином, який виробляється у нирках і посилює проліферацію ЕЧК та їх перетворення в проеритробласти. Цей гормон також стимулює розвиток і розмноження еритроїдних клітин подальших стадій.
Проеритробласти — перші морфологічно розпізнавані клітини еритроїдного ряду, круглої форми, діаметром 15-25 мкм. Ядро велике кругле, розміщене центрально, дрібносітчасто-зернистої структури, містить одне-два ядерця; цитоплазма забарвлюється базофільно, навколо ядра знаходиться світла перинуклеарна зона. Клітини містять багато рибосом, центросому з двома центріолями, характерною є наявність зерен феритину і виростів цитоплазми. Проеритробласти поділяються і перетворюються на базофільні еритробласти.
Базофільні еритробласти мають трохи менші розміри, порівняно з проеритробластами (10-18 мкм), хроматин у них розміщений грудочками променеподібно, як спиці в колесі, цитоплазма базофільна за рахунок великої кількості РНК. Базофільні еритробласти діляться мітозом, починається синтез гемоглобіну і перетворення їх у поліхроматофільні еритробласти.
Поліхроматофільні еритробласти величиною 10-14 мкм, ядро містить щільний хроматин, чіткої колесоподібної структури, ядерця не визначаються. Цитоплазма поліхромна, тобто забарвлюється кислими та основними барвниками. Оксифілія зумовлена наявністю гемоглобіну, а базофілія — РНК. Кількість рибосом у цих клітинах зменшується, феритин розміщується агрегатами. Поліхроматофільні еритробласти діляться мітозом, їхні генерації називають поліхроматофільними нормобластами.
Поліхроматофільні нормобласти розміром до 10 мкм, ядро ущільнюється, втрачає колесоподібне розміщення хроматину, стає пікнотичним. Клітини не здатні до поділу, більшість їх втрачає ядро і перетворюється спочатку в ретикулоцити, а потім в еритроцити. У ретикулоцитах, що проникають у кровоносні судини, ще деякий час зберігається РНК у вигляді сіточки.
Гранулоцитопоез. Джерелом для розвитку зернистих лейкоцитів t також стовбурові та напівстовбурові кров'яні клітини, які одночасно починають диференціюватися через ряд проміжних стадій у трьох різних напрямках, вони продукують клітини трьох видів: нейтрофіли, еозинофіли та базофіли. При цьому утворюються більш детерміновані клітини — попередники і моноцити.
У міру дозрівання гранулоцити зменшуються у розмірах, змінюється форма їх ядер від круглої до сегментованої, у цитоплазмі нагромаджується специфічна зернистість. Диференціація та розмноження клітин цього ряду відбувається під впливом гормона гранулоцитопоетину.
Першою морфологічно розпізнаваною клітиною в напрямі грану- юцитопоезу є мієлобласт. Експериментально доведено, що серед мієлобластів є нейтрофільні, еозинофільні та базофільні мієлобласти. Клітини досягають розміру до 20 мкм, ядро у них кругле або овальне, ніжно-сітчастої будови, містить 2-5 ядерець. У цитоплазмі багато мітохондрій, рибосом, комплекс Гольджі, лізосоми, що характеризуються позитивною реакцією на кислу фосфатазу.
Мієлобласти діляться мітозом і перетворюються у промієлоцити. Останні мають загрубілу будову ядра, їх цитоплазма характеризується появою специфічної зернистості, залежно від якої розрізняють нейтрофільні, еозинофільні та базофільні клітини. Промієлоцити діляться і перетворюються у мієлоцити.
Нейтрофільні мієлоцити розміром 12-16 мкм інтенсивно діляться мітозом. їх цитоплазма дифузно ацидофільна, містить первинні та специфічні гранули, у них є усі органели. В міру дозрівання овальне ядро стає бобоподібним, кількість гетерохроматину збільшується, ядерця не виявляють. Ці клітини належать до юних форм. Вони перетворюються в метамієлоцити. Цитоплазма останніх містить збільшену кількість специфічних гранул. При подальшому дозріванні їх ядро набуває вигляду зігнутої палички. Подібні форми називаються паличкоядерними лейкоцитами. Потім ядро сегментується, і клітина перетворюється у сегментоядерний нейтрофіл. Повний період розвитку нейтрофільного гранулоцита становить приблизно 14 діб.
Еозинофільні мієлоцити. За характером будови ядра мало чим відрізняються від нейтрофільних мієлоцитів. Цитоплазма їх містить оксифільну зернистість. В міру дозрівання еозинофільні мієлоцити діляться, а їх ядро набуває підковоподібної форми. Такі клітини називаються еозинофільними метамієлоцитами. Поступово їх ядро в середній частині набуває ниткоподібної форми, потім стає двочастковим, а у цитоплазмі збільшується кількість специфічних гранул. Клітина не здатна до поділу. Базофільні мієлоцити зустрічаються у меншій кількості, ніж нейтрофільні або еозинофільні. Цитоплазма базофільних мієлоцитів містить у широко варіюючій кількості специфічні базофільні гранули різних розмірів, які проявляють метахромазію при фарбуванні азуром і легко розчиняються у воді. Найменше сегментується ядро у базофілів.
Моноцитопоез. Утворення моноцитів відбувається із стовбурових клітинчервоного кісткового мозку за схемою: стовбурова кровотворна клітина, напівстовбурова кровотворна клітина, із неї утворюється уні- потентний попередник моноцитів. Наступною клітиною, яка морфологічно розпізнавана є монобласт. Це значного розміру клітина, у якої базофільна цитоплазма має вигляд вузенької смужки, що оточує велике ядро з двома ядерцями, монобласт ділиться мітозом, диференціюється у промоноцит. У останнього збільшується кількість цитоплазми, ядро набуває бобоподібної форми, втрачає здатність до мітотичного поділу, перетворюється в моноцит. Моноцити надходять у кровотік, потім проникають у тканини, де й перетворюються у незрілі, а потім зрілі макрофаги.
Лімфоцитопоез. Розвиток лімфоцитів починається із стовбурової кров'яної клітини червоного кісткового мозку, з якої утворюється клі- тина-попередник лімфоцитопоезу.
Важлива особливість цього процесу полягає у тому, що розвиваються морфологічно подібні, але різнорідні за функціональним значенням клітини, тобто розвиток лімфоцитів відбувається у двох напрямах - Т і В-лімфоцитів. В обох рядах виникають уніпотентні попередники, які через лімфобласти (Т і В) перетворюються у лімфоцити (Т і В). Розвиток Т-лімфоцитів продовжується у тимусі, тут вони перетворюються в бласти, які діляться і диференціюються, перетворюючись у ефекторні клітини. У ссавців у червоному кістковому мозку утворюються В-лім- фоцити (у птахів в бурсі Фабриціуса). Тут вони диференціюються, у них з'являються імуноглобулінові рецептори. В подальшому диференційовані В-лімфоцити заселяють селезінку, лімфатичні вузли, лімфатичні вузлики шлунково-кишкового тракту. У цих органах під впливом антигенів відбувається проліферація і подальша спеціалізація В- лімфоцитів з утворенням ефекторних клітин.
Розвиток кров'яних пластинок (тромбоцитів) відбувається в червоному кістковому мозку і пов'язаний з розвитком у ньому гігантських клітин — мегакаріоцитів. У міру дозрівання, у результаті своєрідної багаторазової ендометатичної репродукції, утворюються великі клітини (40-50 мкм), які містять у багатолопастевому ядрі 32-64 хромосомних набори. У цитоплазмі мегакаріоцитів утворюється система мікрофіламентів і мікротрубочок, а також специфічні гранули. У подальшому відбувається фрагментація цитоплазми мегакаріоцитів на відоремленні ділянки — кров'яні пластинки, які через стінки синусоїдів червоного кісткового мозку потрапляють у кровоток.
Ембріональне кровотворення (розвиток крові як тканини) відбувається у стінці жовткового міхура, потім у печінці, червоному кістковому мозку і лімфоїдних органах (тимусі, селезінці, лімфатичних вузлах; у птахів — у фабрицієвій сумці). Порівняно із ссавцями, у птахів добре розвинений жовтковий міхур. У мезенхімі стінки жовткового міхура відмежовуються і ущільнюються скупчення — кров’яні острівці. На їх периферії мезенхімні клітини стають плоскими, з’єднуються між собою і утворюють ендотеліальну стінку судини. Центральні клітини острівця округлюються, втрачають відростки: перетворюються у стовбурові клітини крові (СКК). Частина СКК диференціюється у первинні клітини крові — бласти. Це великі клітини з базофільною цитоплазмою і ядром з добре помітними ядерцями. Частина первинних кров'яних клітин після мітотичного ділення перетворюється у первинні еритробласти, що мають великий розмір і називаються мегалобластами. У їх базофільній цитоплазмі нагромаджується гемоглобін і вони перетворюються в поліхроматофільні, а потім в оксифільні еритробласти з великою кількістю гемоглобіну.
Деякі первинні еритробласти втрачають ядро, в інших воно зберігається. Внаслідок цього утворюються без'ядерні та ядерні первинні еритроцити, які відрізняються від нормоцитів великими розмірами і тому їх називають мегалоцитами. Такий тип кровотворення називається мегалобластичним. Він характерний для ембріонального періоду, але може з’являтися і після народження при деяких захворюваннях (злоякісне недокрів'я). Поряд з мегалобластичним у стінці жовткового міхура починається нормобластичне кровотворення, у результаті якого із бластів утворюються вторинні еритробласти; спочатку вони перетворюються у поліхроматофільні еритробласти, потім в нормо- бласти, із яких утворюються вторинні еритроцити (нормоцити). Розміри яких відповідають еритроцитам (нормоцитам) дорослої тварини. Розвиток еритроцитів у стінці жовткового міхура відбувається інтраваскулярно; крім того, в стінці жовткового міхура екстраваскулярно з частини первинних клітин-бластів утворюється невелика кількість нейтрофілів та еозинофілів.
Частина СКК розноситься з кров'ю до різних органів зародка, у яких починається процес кровотворення. На п'ятому тижні ембріогенезу великої рогатої худоби кровотворення починається у печінці, вона стає універсальним органом, де екстраваскулярно утворюються еритроцити, зернисті лейкоцити, мегакаріоцити. В кінці ембріонального періоду розвитку кровотворення в печінці закінчується. На 7-8-му
тижні у великої рогатої худоби диференціюються лімфоцити тимуса і, мігруючи із нього, Т-лімфоцити заселяють Т-зони селезінки, лімфатичних вузлів. Селезінка також орган, у якому утворюються усі різновиди формених елементів крові.
На четвертому місяці ембріонального розвитку починається кровотворення у червоному кістковому мозку. У період ембріонального еритропоезу замінюється генерація утворення еритроцитів, які відрізняються морфологією і типом утворення гемоглобіну. Ембріональний (фетальний) тип гемоглобіну утворюється первинними еритроцитами; фетальний тип гемоглобіну — еритроцитами в печінці та селезінці. Дефінітивний тип гемоглобіну утворюється еритроцитами червоного кісткового мозку.
Власне сполучні тканини
Власне сполучну тканину поділяють на: волокнисту і тканину зі спеціальними властивостями, волокниста сполучна, тканина подібно до інших різновидів утворена з клітин та міжклітинної речовини, обсяг якої перевищує об’єм клітин. Міжклітинна речовина утворена основною речовиною та волокнами. Залежно від цього співвідношення цю тканину поділяють на пухку і щільну.
Для сполучної тканини зі спеціальними властивостями діритаманний переважний розвиток окремих клітинних елементів та деякі особливості міжклітинної речовини. В залежності від цього її поділяють на: жирову, пігментну, ретикулярну і слизову.
Ці тканини становлять велику групу сполучних тканин зі спеціальними властивостями. У тілі вищих хребетних вони значно поширені і виконують різноманітні механічні та формоутворюючі функції, їм належить важливе місце у забезпеченні реакції імунного захисту — фагоцитозі, утворенні імуноглобулінів та інших біологічно активних речовин. Сполучні тканини виконують трофічну і пластичну функції тощо.
Пухка сполучна тканина.
Цій тканині належить важливе місце в життєдіяльності організму. Вона є у складі майже всіх органів — заповнює проміжки між ними, утворює їх оболонки та прошарки. Крім опорно-механічної функції, пухка сполучна тканина виконує захиснул трофічну функції, а також бере участь у пластичних процесах при загоюванні ран, утворенні капсули навколо стороннього тіла тощо.
До складу пухкої сполучної тканини входять численні клітини та міжклітинна речовина. Усі її клітини розмежовані міжклітинною речовиною. Більшість з них здатні до розмноження і міграції, знаходяться в єдиному функціональному зв'язку з клітинами судинної крові та лімфоїдної системи організму.
Адвентиційні або періцити (від лат. adventicus - прийдешній) клітини. Вони мають видовжену або зірчасту форму, овальне ядро, базо- фільну цитоплазму і незначну кількість органел. Ці клітини розташовуються на зовнішніх стінках гемокапілярів, здатні до мітозу і диференціації в інші клітини (фібробласти, міофібробласти і адіпоцити). Деякі автори заперечують існування цих клітин.
Фібробласти (від лат. fibra — волокно і гр. blastos — зародок ), виникають з мезенхімних клітин. У пухкій сполучній тканині вони зустрічаються частіше від інших клітин, продукують різноманітні компоненти міжклітинної речовини^ синтезують і виділяють високомолекулярні речовини для утворення волокон основної речовини (рис. 50, 51).
В постембріональному періоді і при регенерації попередниками фібробластів є адвентиційні клітини. Залежно від їх зрілості, будови та функціонального значення фібробласти поділяють на три різновиди. Малодиференційовані фібробласти (20-25 мкм) веретеноподібної форми з незначною кількістю коротких відростків. Цитоплазма базофільна, під електронним мікроскопом в ній виявляють невелику кількість вільних рибосом, вузькі канальці гранулярної ендоплазматичної сітки, комплекс Гольджі, мітохондрії. Ядро овальне, має ядерце. Малодиференційовані фібробласти здатні до активного мітозу. Вважається, що ці клітини продукують глікозаміноглікани. Зрілі фібробласти пластинчастої форми, з відростками досягають 50 мкм; на поперечному розрізі веретеноподібні за рахунок потовщеного ядра. В цитоплазмі фібробласта розрізняють дві зони — внутрішню ендоплазму (більш базофільну), зовнішню ектоплазму (її забарвлення слабше). Ядро овальне, світле, багате на еухроматин і містить 1-2 ядерця. Фібробласти здатні рухатися за рахунок скоротливих білків актину і міозину, які розміщені в мікрофіламентах ектоплазми.
Цитоплазма зрілих фібробластів містить добре розвинену гранулярну ендоплазматичну сітку, комплекс Гольджі у вигляді цистерн і міхурців, тут синтезуються глікозаміноглікани, різної форми і розміру мітохондрії, лізосоми. Фібробласти, крім глікозаміногліканів синтезують білки, проколаген, проеластин, ферментні білки, а також фібрилярний глікопротеїд фібронектин, який впливає на рух клітин і зв'язування їх з мікрооточенням.

Рис. 50. Пухка сполучна тканина:
1-фібробласт; 2-макрофаг; 3-тканинний базофіл; 4-колагенові волокна; 5-еластичні волокна
Фіброцити — веретеноподібні клітини з видовженим ядром, знаходяться між волокнами сполучної тканини. Фіброцити містять незначну кількість органел, нездатні до розмноження, характеризуються зниженими синтетичними процесами.
Міофібробласти. Функціональну дію проявляють у грануляційній тканині; містять розвинену ендоплазматичну сітку та скоротливі фібрили. Ці клітини є похідними фібробластів.
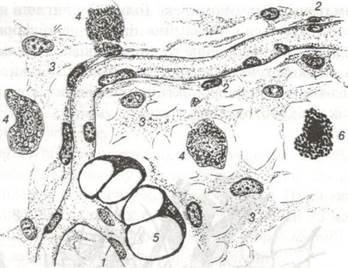
Рис. 51. Пухка сполучна тканина із підшкірної клітковини:
1-ендотелій капіляра; 2-камбіальна клітина; 3-фібробласт; 4-макрофаг; 5-жи- рова клітина; 6-тканинний базофіл.
Гістіоцити - макрофаги пухкої сполучної тканини, котрі диференціюються з моноцитів крові. Вони розміщуюються поодиноко або групами. Розмір їх 12-18 мкм, здатні до міграції, форма кругла, витягнута. Цитоплазма базофільно-плямиста, містить помірні мітохондрії і комплекс Гольджі, гранулярна ендоплазматична сітка майже не розвинена. Багато в них лізосом та фагосом, за їх допомогою перетравлюються фагоцитовані речовини. Ядро невелике, овальне, містить багато гетерохроматину, інтенсивно забарвлюється. Під електронним мікроскопом в гістіоцитах виявлені псевдоподії, пальцеподібні вирости, мікроворсинки, що відходять від їх поверхні. За їх допомогою макрофаги захоплюють макромолекулярні частинки, мікроорганізми, віруси, продукти клітинного розпаду. На поверхні плазмолеми гістіоцита містяться рецептори для пухлинних клітин, еритроцитів, Т- і В-лімфоцитів, імуноглобулінів тощо. Наявність рецепторів макрофагів забезпечує їх участь в імунних реакціях. Попередниками гістіоцитів є моноцити периферійної крові. Моноцити, які мігрують із судин, у відповідних середовищах адаптуються і перетворюються у органо- і тканинноспецифічні макрофаги, утворюють макрофагічну систему.
Макрофаги — багатофункціональні клітини. їм належить важливе місце у формуванні природного і набутого імунітету. У природному імунітеті участь макрофагів проявляється в здатності до фагоцитозу і піноцитозу. Фагоцитозний матеріал всередині макрофага піддається ферментативному розщепленню за допомогою лізосом. Так ліквідуються шкідливі для організму речовини. Макрофаги синтезують активні речовини — лізоцим, інтерферон, пірогени тощо. В набутому імунітеті їх значення проявляється в пізнанні антигена, його перетворенні з корпускулярної форми в молекулярну, передачі імунокомпетентним клітинам і виконання ефекторної функції при взаємодії з Т-і В-лімфоцитами.
Окрім гістіоцитів до макрофагів належать зірчасті клітини Куафера (печінки), вільні та фіксовані макрофаги лімфовузлів, селезінки, червоного кісткового мозку, альвеолярні макрофаги легень, плевральні та перитонеальні макрофаги, остеокласти, клітини Лангерганса (шкіра), клітини мікроглії (нервова тканина), макрофаги синовіальних оболонок суглобів та інших органів.
Тканинні базофіли (мастоцити, лаброцити, тучні клітини) походять від стовбурової кровотворної клітини (СКК).
Термін «тучні клітини» вперше запропонував Ерліх в 1877 р. для позначення клітин, які були насичені гранулами. Кількість тканинних базофілів і базофілів крові деяких тварин має зворотню залежність. Тканинні базофіли є у дрібних кровоносних судинах сполучної тканини органів травлення, молочної залози, шкіри, тимуса, матки, яйцепроводу птахів. За формою вони округлі, овальні, веретеноподібні тощо. Розмір тканинних базофілів 10-25 мкм і більше. У птахів вони значно менших розмірів, ніж у ссавців. Ядро цих клітин у більшості випадків оточене гранулами, кругле, займає центральне положення, має звичайну будову. Серед органел тканинні базофіли містять добре розвинений комплекс Гольджі, велику кількість мітохондрій, небагато гранулярної та агранулярної ендоплазматичної сітки.
Головною складовою частиною тканинних базофілів є велика кількість базофільних гранул (0,3-1 мкм), рівномірно розміщених в цитоплазмі. Вони характеризуються метахромазією, зумовленою сульфатованим глікозаміногліканом. Гранули тканинних базофілів містять ряд біологічно активних речовин, серед яких гепарин становить 30% вмісту гранул, а також гістамін, хондроітинсульфат, дофамін,
серотонін. Гепарин знижує проникність міжклітинної речовини, запобігає зсіданню крові. Гістамін викликає скорочення міоцигів, сприяє виходу плазми з вєнул і капілярів за рахунок розширення і підвищення проникності їх стінки. Гістамін стимулює міграцію еозинофілів, активізує макрофаги. Тканинні базофіли одні із перших реагують на проникнення антигенів клітин із крові. На плазмолемі знаходиться значна кількість імуноглобулінів, що належать до класу JgC. Зв'язування антигенів і утворення комплексу антиген — антитіло супроводжується дегрануляцією і виділенням із тканинних базофілів судинноактивних речовин, які зумовлюють появу місцевих і загальних реакцій.
Плазматичні клітини (плазмоцити). Займають фундаментальне місце в імунних реакціях. Плазматичні клітини забезпечують утворення антитіл, що циркулюють в крові. Ці антитіла називаються гуморальними (від лат. humor — рідина). Плазматичні клітини високоспеціалі- зовані. Вони синтезують і виділяють основну масу імуноглобулінів. Походять плазматичні клітини з В-лімфоцитів під впливом антигенів. Антиген (від гр. anti — проти, genon — утворювати) означає речовину, проти якої утворюється інша речовина.
Значна частина плазматичних клітин знаходиться під епітелієм органів дихання, кишечника, у лімфатичних вузлах, селезінці, мигдалику, сполучній тканині слізної залози. За формою вони більш-менш округлі, розміром до 12 мкм.
Цитоплазма, порівняно з об’ємом ядра, займає значно більшу площу. Ядро округле, міститься ексцентрично. Значна частина хроматину знаходиться у конденсованому стані; хроматин часто розміщується подібно цифрам на циферблаті годинника. Цитоплазма інтенсивно базофільна, на її фоні біля ядра помітне «світле подвір'я», де знаходиться комплекс Гольджі. Базофілія цитоплазми зумовлена великою кількістю РНК. Цитоплазма плазмоцитів спеціалізована для біосинтезу білкового секрету, який знаходиться в добре розвинених цистернах гранулярної ендоплазматичної сітки. Синтезовані молекули імуноглобулінів надходять у просвіт цистерни, а потім в комплекс Гольджі, звідки після приєднання вуглеводного компонента виводяться на поверхню клітини. Інші деталі будови плазматичної клітини звичайні, за винятком того, що плазматична мембрана цих клітин часто утворює пальцеподібні відростки. Плазматичні клітини не здатні до поділу, вони старіють, гинуть і поглинаються макрофагами.
Адіпоцити (від лат. adeps — жир) - це клітини здатні накопичувати в своїй цитоплазмі резервний жир і мають кулясту форму. їх розміри залежать від кількості жиру і досягають 120 мкм. Жир з’являється в них спочатку у вигляді дрібних капель, котрі зливаються в одну велику краплю, яка заповнює всю клітину. Внаслідок цього ядро розміщується під гілазмолемою. Такі клітини на поперечному розрізі набувають форму «каблучки».
Значне накопичення жиру інколи призводить до розриву плазмо- леми клітини і жир виходить в міжклітинну речовину. При електронній мікроскопії в цих клітинах спостерігається майже повна відсутність ендоплазматичної сітки, слабо розвинений комплекс Гольджі. Проте в клітинах бурого жиру ці органели розвинені значно краще.
Пігментоцити - ці клітини в нормальній сполучній тканині ссавців трапляються рідко. Вони знаходяться переважно в стінці очного яблука (в склері, судинній та райдужній оболонках).
В окремих ділянках шкіри (вим’я, мошонка) кількість пігментоцитів значна. Ці клітини мають зірчасту форму і зерна пігменту - меланіну. Одні з цих клітин здатні синтезувати і накопичувати пігмент, інше - лише накопичувати.
У нижчих хребетних пігментні клітини (хроматофори) спостерігаються постійно. Мають відросчасту, неправильну форму. Під регулюючим впливом нервової системи хроматофори можуть активно скорочуватися і внаслідок чого змінюється колір шкіряного покрову, що робить тварину малопомітною.
Міжклітинна речовина пухкої сполучної тканини
До складу міжклітинної речовини входить основна речовина (аморфна) а також колагенові, еластичні та ретикулінові волокна.
Основна речовина - це желеподібне гідрофільне середовище. Воно бере участь в транспорті метаболітів поміж клітинами і кров’ю, в механічній, опорній та захисній функціях.
До її складу входять сульфатовані глікозамінглікани, такі як хондроітинсульфат, дерматансульфат, кератинсульфат та гепаринсульфат, які зв’язані з білками і утворюють протеоглікани. Ці речовини обумовлюють консистенцію і функціональні особливості основної речовини. Крім вказаних компонентів до її складу входять: ліпіди, альбуміни та глобуліни крові, мінеральні речовини (солі Na, К, Са, та інші.)
Кількість основної речовини в різних ділянках сполучної тканини неоднакова. Навколо капілярів і дрібних судин в ділянках, що містять жирові прошарки, основної речовини мало, а на межі з тканинами іншого походження, наприклад з епітелієм її багато.
Аморфний компонент цієї речовини бере участь в метаболізмі води, регулюванні іонного складу в з’єднанні клітин і волокон, адгезії клітин та інші.
Фізико-хімічний стан міжклітинної речовини в значній мірі визначає функціональні особливості сполучної тканини. Чим щільніша міжклітинна речовина, тим більше виражена механічна і опорна функції. Трофічна функція навпаки краще забезпечується напіврідкою за консистенцією міжклітинною речовиною. Під дією ендогенного та екзогенного гістаміна, гіалуронідази відбувається підвищення проникливості аморфного компонента міжклітинної речовини. Підвищення концентрації глікозамінгліканів, зниження активності гіалуронідази навпаки знижує його проникливість.
Колагенові (від гр. colla — клей) волокна дуже міцні. При варінні вони утворюють клейку речовину. У сполучній тканині колагенові волокна мають різноспрямоване положення. Це хвилеподібні, спіралеподібно скручені округлі тяжі, завтовшки 20-30 мкм. Вони утворені з фібрилярного білка колагену, який синтезується рибосомами гранулярної ендоплазматичної сітки фібробластів. Розрізняють певні рівні організації колагенових волокон.
Молекулярний рівень відбувається в цитоплазмі фібробластів. Молекула колагена має довжину до 280 нм, побудована з трьох полі- пептидних ланцюгів (а-ланцюги). Попередник колагену - прокола- ген утворює спіраль. Кожен ланцюг утворений з трьох різних амінокислот, які багаторазово повторюються протягом її довжини. Перша амінокислота може бути будь-якою, друга пролін або лізин, третя - гліцин. Пролін і лізин окислюються у гідроксилізин - маркерні амінокислоти зрілого колагену. Проколаген надходить у міжклітинну речовину.
Наступний етап - надмолекулярний, проходить шляхом агрегування в довжину поперечно пов’язаних водневими зв’язками молекул колагену. Потім утворюються протофібрили з 5 - 6 мікрофібрил.
Третій період - за участю глікозамінгліканів і глікопротеїнів, які також продукуються фібробластами, утворюються пучки фібрил поперечно-посмугованої структури товщиною 50-100 нм. Період повторення темних і світлих ділянок через кожні 64 нм.
В четвертому періоді — утворюються пучки волокон товщиною 150 мкм. Розрізняють декілька типів колагену. Вони різняться за молекулярною організацією та локалізацією в тканинах. Колаген першого типу знаходиться у сполучній тканині шкіри, кісток, рогівці ока, та склері; другого типу - в гіаліновому та волокнистому хрящах та скловидному тілі ока; третього типу - в шкірі плода, стінці великих кровоносних судин та ретикулярних волокнах; четвертого типу — в базальних мембранах і капсулі кришталика; п’ятий тип - навколо фібробластів, ендотеліоцитів та міоцитів у вигляді екзоскелету. Колагени від шостого до дванадцятого типу вивчені недостатньо.
Еластичні волокна. Наявність їх у пухкій сполучній тканині визначається еластичністю і розтягуваністю. По товщині і міцності вони поступаються колагеновим волокнам. На поперечному розрізі мають плескату форму, розгалужуються і анастомозують поміж собою.
Основним хімічним компонентом цих волокон є глобулярний білок, що синтезується фібробластами. В порівнянні з колагеном в цьому білку міститься значно менше полярних амінокислот. Для елас- тина характерна відсутність цистіна і наявність двох похідних амінокислот - десмозина та ізодесмозина, які надають еластину здатності до розтягування і еластичності.
Електронна мікроскопія дозволила встановити, що еластичні волокна в центрі містять аморфний компонент, а по периферії - мікро- фібрилярний.
Ретикулярні волокна (від лат. reticulum — сітка) можна спостерігати на препаратах іпрегнованих азотнокислим сріблом. Тому їх ще називають аргірофільними. Розрізняють два види волокон: власне ретикулярні або дефінітивні утворення з колагену третього типу, та преколагенові - початкова стадія утворення в період ембріогенезу та при регенерації.
Ретикулярні волокна дуже близькі до колагенових за своїм складом, але відрізняються від них меншою товщиною, розгалуженістю та наявністю анастомозів. Разом з ретикулярними клітинами, котрі їх утворюють вони, формують сітчасті структури. При електронній мікроскопії в цих волокнах виявлені протофібрили товщиною 40 нм. склеєні аморфною речовиною. Ретикулярні волокна мають високу концентрацію ліпідів, вуглеводів та сірки. Вони стійкі до дії кислот,
лугів і трипсину. За здатністю до розтягання вони займають проміжне положення між колагеновими і еластичними волокнами.
Сполучна тканина зі спеціальними властивостями
До неї входять: жирова, ретикулярна та слизова тканини.
Жирова тканина утворена адіпоцитами та пухкою сполучною тканиною. Групи адіпоцитів утворюють частки. Значні нагромадження часток жиру називають жировою тканиною. Між жировими частками знаходяться прошарки пухкої сполучної тканини, у яких проходять кровоносні судині і нервові волокна. Поміж жировими клітинами в частках містяться окремі фіброцити, лімфоцити, тканинні базофіли, аргирофільні волокна, кровоносні капіляри. Жирова тканина в організмі тварини може становити до 30% від живої маси. Її кількість тісно пов’язана з годівлею, нагулом, станом ендокринної системи тощо. У ссавців існує біла та бура жирова тканина. Біла жирова тканина міститься у жирових депо — підшкірній жировій тканині та у брижі, що особливо характерно для свиней; у деяких порід овець — біля кореня хвоста (курдюк), у прошарках сполучної тканини між м'язовими пучками тощо. Це висококалорійна речовина, при окисленні якої звільняється велика кількість енергії, а також вода. Жирова тканина виконує амортизаційну функцію, зберігає тепло тощо. Структурною одиницею білої жирової тканини є кулясті клітини, які досягають 120 мкм.
Бура жирова тканина побудована із дрібних клітин, щільно розміщених між собою і оточених капілярами та нервовими волокнами. Бура жирова тканина в значній кількості міститься у тілі гризунів і тварин, що впадають в зимову сплячку. Для її клітин характерне центральне розміщення ядра та дрібних жирових крапель, їх злиття в більш велику краплю не відбувається. У проміжках між жировими краплями знаходяться численні мітохондрії, а також інші органели. Бурий колір жирових клітин утворюється за рахунок цитохромів мітохондрії. При зниженні температури зовнішнього середовища активність окислювальних процесів у бурій жировій тканині підвищується. При цьому виділяється теплова енергія, яка зігріває кров у капілярах. Бура жирова тканина є важливою для регулювання температури у новонароджених і зігріванні тварин після пробудження від зимової сплячки.
Nретикулярна тканина утворює строму кровотворних органів, в яких вона добре розвинена. В комплексі з макрофагами забезпечує специфічне мікрооточення для проліферації, диференціації та міграції клітинних елементів.
Ретикулярна тканина також входить до складу слизових оболонок,Ів меншій мірі — печінки. Вона утворена із ретикулярних клітин зірчастої форми і волокон, які формують сітку (від лат. reticulum - сітка ). В комірках сітки міститься тканинна рідина, лімфоцити, макрофаги, плазмоциди та інші клітини, ^а походженням вона є похідним мезенхімид. Ядро ретикулярних клітин округле, характеризується помірним вмістом хроматину;^цитоплазма містить малочисельні мітохондрії та комплекс Гольджі на різних рівнях розвитку, інші органели. Ретикулінові волокна знаходяться у глибоких інвагінаціях плазмолеми ретикулярних клітин.
Слизова сполучна тканина спостерігається лише в пупковому канатику плода. Клітинні елементи представлені тут у вигляді мукоцитів - клітин типу фібробластів. Головна особливість цієї тканини - це відсутність волокнистих структур.
Основна міжклітинна речовина містить високомолекулярні біополімери, які забезпечують тургор (пружність) тканин пухкого канатика і попереджають можливість перетискання кровоносних судин, що живлять зародок.
Ретикулярна тканина утворює строму кровотворних органів, в яких вона добре розвинена. В комплексі з макрофагами забезпечує специфічне мікрооточення для проліферації, диференціації та міграції клітинних, елементів.
Ретикулярна тканина також входить до складу слизових оболонок, в меншій мірі — печінки. Вона утворена із ретикулярних клітин зірчастої форми і волокон>які формують сітку (від лат. reticulum -сітка ). В комірках сітки міститься тканинна рідина, лімфоцити, макрофаги, плазмоцити та інші клітини. За походженням вона є похідним мезенхіми. Ядро ретикулярних клітин округле, характеризується помірним вмістом хроматину; цитоплазма містить малочисленні мітохондрії та комплекс Гольджі на різних рівнях розвитку, інші органели. Ретикулінові волокна знаходяться у глибоких інвагінаціях плазмолеми ретикулярних клітин.
Щільна волокниста сполучна тканина
Відмінною особливістю цієї тканини є переважний розвиток міжклітинної речовини, головним чином колагенових волокон, дуже міцних і майже нездатних до розтягування. У складі сухожилля ці волокна витримують навантаження до 6 кг на 1 мм2 площі розрізу. Виходячи з цього, утвореним таким колагеном структурам характерна значна міцність на розрив. Щільна волокниста сполучна тканина залежно від способу орієнтації колагенових волокон у просторі поділяється на сформовану і несформовану. В першому випадку всі колагенові волокна розміщені в одній площині і мають один напрямок.
Сформована щільна волокниста сполучна тканина утворює сухожилля, фіброзні мембрани, зв'язки (рис. 52), за допомогою яких м'язи з'єднуються з кістками, де тяга прикладається в одному напрямку. Між окремими пучками колагенових волокон, що складаються з численних фібрил, розміщені високодиференційовані клітини — фіброцити, що забезпечують фізіологічну регенерацію цієї тканини. Пучок колагенових волокон, оточений шаром фіброцитів, називають сухожильним пучком першого порядку. На його повздовжньому розрізі фіброцити розміщені у вигляді ланцюжка. Кілька сухожильних пучків першого порядку в сукупності утворюють сухожильні пучки другого порядку. Вони розмежовуються між собою прошарками пухкої сполучної тканини, що має назву ендотенонію. Пучки другого порядку, в свою чергу, об'єднуючись утворюють сухожильні пучки третього і, навіть, четвертого порядків. Вони оточені пухкою сполучною тканиною, яку називають перитенонієм. Пухка сполучна тканина, що утворює ендотеноній та перитеноній, містить основну міжклітинну речовину, фібробласти, макрофаги, судинно-нервові пучки.

Рис. 52. Щільна сформована сполучна тканина сухожилля на поздовжньому зрізі:
1-ядра фіброцитів; 2-ендотеноній; 3-сухожильні пучки другого порядку.
В несформованій щільній сполучній тканині колагенові волокна лежать у різних напрямках. Вона входить до складу сітчастого шару шкіри, апоневрозів, капсул лімфовузлів, селезінки та інших органів, колагенові пучки ідуть в різних напрямках, що забезпечує їх міцність при різноманітних напрямках дії механічних факторів. Поряд з пучками колагенових волокон несформована щільна волокниста сполучна тканина вміщує прошарки пухкої сполучної тканини (рис. 53).
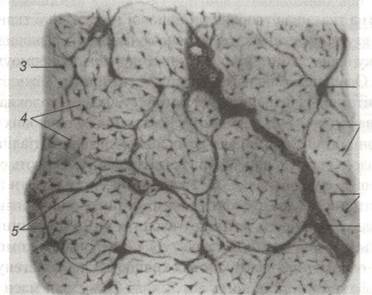
Рис. 53. Щільна сформована сполучна тканина сухожилля теляти на поперечному розрізі:
1-пучки першого порядку; 2-фіброцити: 3-ядро фіброцита; 4-відростки ядра;
5-прошарки пухкої сполучної тканин між пучками другого порядку; 6-прошарки пухкої сполучної тканини.
Щільна сформована сполучна еластична тканина входить до складу потиличної (надостистої) зв'язки, що добре розвинена у коней та великої рогатої худоби. Складається вона з сітки еластичних волокон, які поздовжньо витягнуті в одному напрямку; мають у нативному вигляді жовтий колір, розгалужуються і анастомозуючись між собою, зумовлюють значну еластичність.
На відміну від щільної колагенової волокнистої сполучної тканини вона не утворює пучків різних порядків, оскільки пухка сполучна тканина не розподіляється у ній по всій еластичній сітці. В щілиноподібних просторах між еластичними волокнами знаходяться фіброцити та тонкі колагенові фібрили, що переплітаються між собою. Місцями у широких прошарках пухкої сполучної тканини знаходяться судини мікроциркуляторного русла.
Щільна несформована еластична тканина у вигляді циркулярно розміщених пластинчатих мембран і еластичних сіток входить до складу середньої оболонки судин еластичного типу — аорти і легеневої артерії, а також в сітчастому шарі шкірі, окісті та охрясті.