Основи еволюції - Корж О.П. - 2006
Частина I. МІКРОЕВОЛЮЦІЯ
Розділ 7. Природний добір
7.3. Форми природного добору
Існують різні підходи до проблеми класифікації форм природного добору, яких нині налічують понад 30. Деякі вчені намагаються розрізняти в доборі процес і результат - як позитивне (творча дія активного формування ознаки), так і негативне (редагування шляхом простого усунення окремих форм). На думку М.В. Тимофєєва-Ресовського та ін., подібні підходи безперспективні, оскільки добір завжди є процесом (причому як з позитивним, так і негативним спрямуванням одночасно), а оцінювати його дію можна лише за результатами через певний проміжок часу.
Одну з класифікацій, на наш погляд, надмірно деталізовану, запропонували Ю.І. Рожков та О.В. Проняєв. Згідно з нею існуюче різноманіття типів мінливості організмів, умов середовища, популяційних та видових систем породжує різноманітність процесів добору, згідно з чим слід проводити й класифікацію останнього.
Запропонована цими авторами класифікація ґрунтується на виділенні трьох типів природного добору: індивідуального, позиційного та групового. Але, на їх думку, добір, що діє за кількісними та якісними ознаками, має певні відмінності (табл. 7.1).
Таблиця 7.1. Найважливіші типи й форми добору, які діють за кількісними ознаками (за Ю.І. Рожковим, О.В. Проняєвим, 1994)
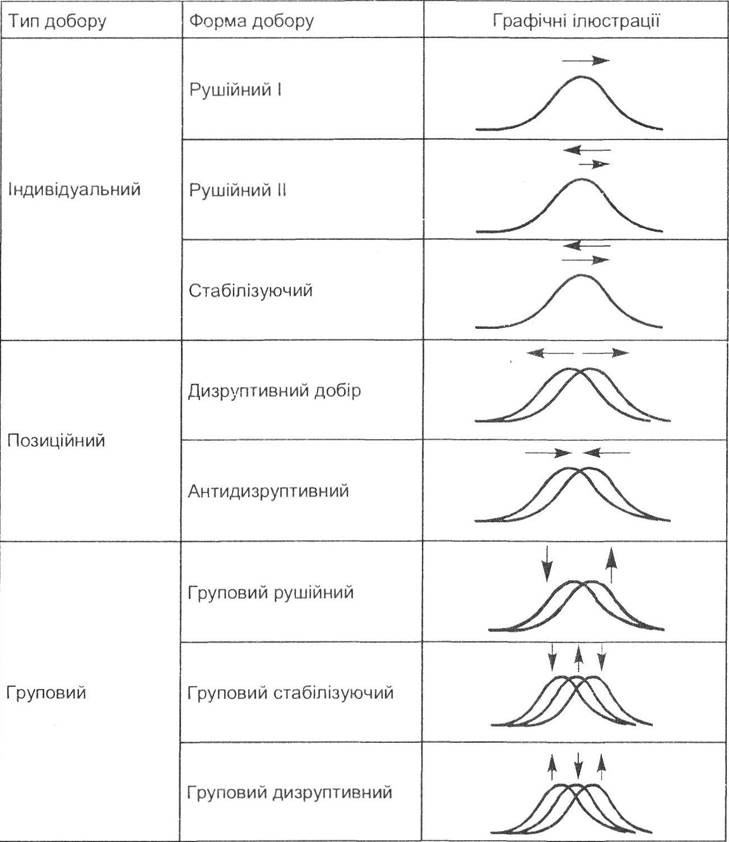
Таблиця 7.2. Найважливіші типи й форми добору, які діють за якісними ознаками (за Ю.І. Рожковим, О.В. Проняєвим, 1994)
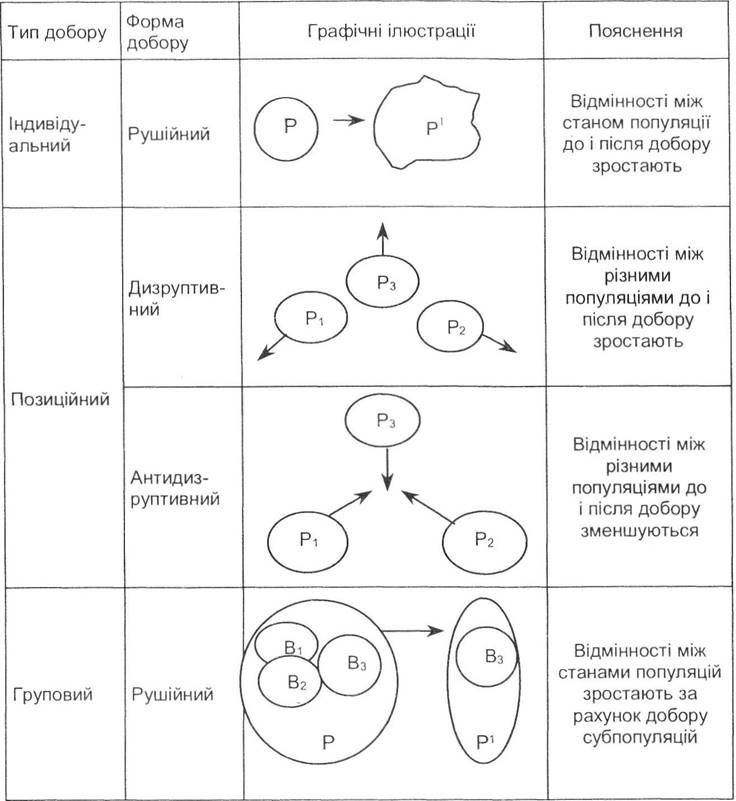
Індивідуальний добір - це відбирання окремих особин у популяціях. У випадку дії за кількісними ознаками розрізняють три форми цього добору - рушійну І, рушійну II та стабілізуючу. Якщо ж добір діє за якісними ознаками певних організмів, то зберігається лише рушійна форма.
Груповий добір стосується водночас цілих груп різного рангу — від сімейних до популяцій і навіть видів. Форми цього типу повністю збігаються з попередніми як у випадку дії за кількісними, так і якісними ознаками.
Позиційний добір є проміжним між попередніми типами і відбувається в тому разі, коли індивідуальні зміни всередині кількох груп зумовлюють їх взаємне віддалення (дизруптивна форма) або ж взаємне наближення (антидизруптивна форма). Форми цього типу, на відміну від попередніх, властиві дії добору як за кількісними, так і за якісними ознаками.
Визнаючи оригінальність наведеного підходу, зазначимо, що доцільніше було б виділити (зрозуміло, що умовно) монофакторні та поліфакторні форми добору. Якщо ж дотримуватись класифікації явищ добору на запропонованій нами основі, то до монофакторних можна було б віднести лише три класичні форми - рушійну, стабілізуючу та дизруптивну. Усі інші варіанти, як наведені вище, так і ті, про які не згадувалося (наприклад, частотнозалежний добір), є поліфакторними і можуть бути подані у вигляді комплексу попередніх елементарних форм.
Еволюційний процес завжди відносно тривалий і переважно полягає у збалансованій дії всіх зазначених елементарних форм природного добору. На кожному етапі формування ознаки, за якою досліджується стан певних популяцій, можна побачити дію різних форм добору. На початкових етапах завжди переважають дизруптивні процеси виокремлення нової (або нових) форм з материнської. Згодом вони переходять у рушійні, спрямовані на формування нових адаптивних комплексів новостворених форм. Завершує процес стабілізуюча форма добору, яка закріплює отримані комплекси ознак.
Оскільки в природі ми стикаємося переважно з результатами дії добору (вже адаптовані до певних умов організми), то спостерігаємо значне переважання стабілізуючої форми. Частотнозалежний добір може розглядатись як проміжний етап формування певної ознаки в популяції. Він полягає у почерговій зміні напрямку дії природного добору відповідно до змін умов, переважно незначних для життєздатності організму, наприклад, самиці дрозофіл спаровуються з білоокими або червоноокими самцями, коли вони становлять меншість у популяції. З часом популяція може втратити частину генетичної інформації, яка зумовлювала поліморфізм та періодичні зміни напрямку дії добору. Це може призвести до переважання стабілізуючої форми добору.
Первинні форми добору
До них ми можемо віднести три класичні форми, визнані більшістю фахівців, які слід вважати відносно монофактор- ними: рушійну, стабілізуючу та дизруптивну.
У природі жоден з екологічних чинників не спрацьовує окремо, але якщо інтегрований вплив довкілля на популяцію має єдине спрямування, його умовно можна вважати монофакторним. Інколи виділяють лише дві форми природного добору, об'єднуючи рушійну і дизруптивну.
Рушійна форма природного добору - найбільш типова форма, виділена Ч. Дарвіном, яка зумовлює утворення нового комплексу адаптаційних ознак. Зміст її полягає в зміщенні середньої ознаки в популяції згідно з загальним напрямком тиску довкілля (рис. 7.1). Рушійний добір зумовлює вироблення нових пристосувань шляхом спрямованої перебудови генофонду популяції, що, у свою чергу, супроводжується перебудовою генотипу особин. Ця форма переважає в умовах постійних змін навколишнього середовища, а наслідком її дії є або зміна норми реакції, або ж її розширення.
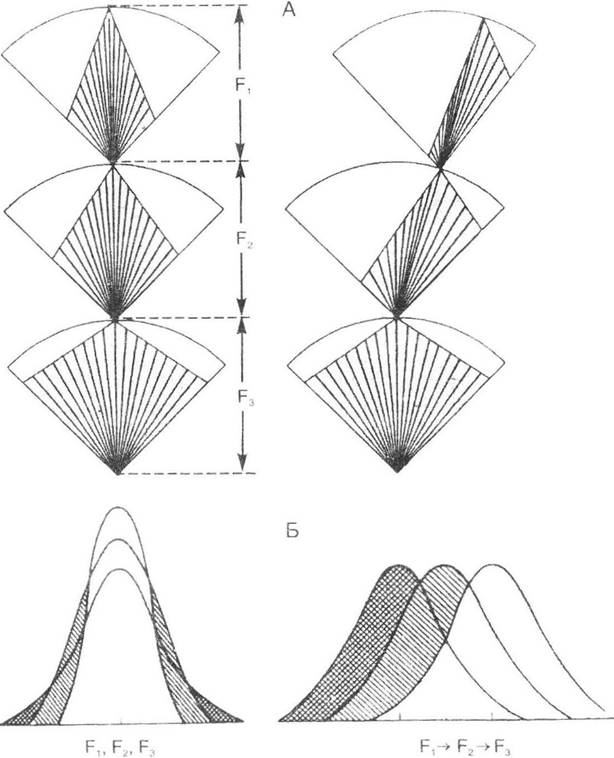
Рис. 7.1. Порівняння рушійної та стабілізуючої форм природного добору
(за М.В. Тимофєєвим-Ресовським, М.М. Воронцовим, О.В. Яблоковим, 1969):
А - дія добору всередині одного потомства; Б - тиск добору в популяції
Дію цієї форми природного добору можна проілюструвати явищем індустріального меланізму: через забруднення середовища відбувається витіснення світлих метеликів темнозабарвленими. Так, в Англії за останні 120 років із 700 видів метеликів близько 70 змінили своє світле забарвлення на темніше. Аналогічним прикладом можна вважати появу комах, стійких до дії інсектицидів, та мікроорганізмів, несприйнятливих до антибіотиків. Наведені факти демонструють пристосувальне значення дії природного добору, хоча ми ще не можемо говорити про створення нового виду (за своїм змістом воно ближче до елементарного еволюційного явища).
Стабілізуюча форма природного добору була докладно досліджена І.І. Шмальгаузеном. За теорією Ч. Дарвіна, наслідками дії добору є формування нових адаптивних ознак і усунення тих, які втратили своє адаптивне значення, а також збереження корисних пристосувань. Спочатку вважалося, що ознаки організмів у поколіннях не змінюються, оскільки відповідні гени зберігаються незмінними. Але наявність спадкової мінливості переконливо свідчить, що подібне явище стійкості ознак потребує додаткового пояснення, яке й надає введена І.І. Шмальгаузеном форма природного добору.
Зміст стабілізуючого добору полягає в тому, що в тривало стабільних умовах середовища селективну цінність набувають організми з найбільш оптимальними характеристиками (так звані середнячки). Якщо умови довкілля залишаються відносно стабільними (коливання факторів середовища мають випадковий або періодичний характер) і певний вид живих організмів пристосований до цих «звичних» умов, то фенотипи окремих особин також зазнають випадкових або ж періодичних змін. Доки середовище залишається без суттєвих змін, доти фенотип виду зберігається відносно сталим (рис. 7.1). Отже, дія цієї форми добору спрямована на збереження організмів із середніми ознаками й унеможливлення проявів фенотипічної мінливості (враховуючи й мутації, що відхиляються від середнього фенотипу). Результатом дії стабілізуючого добору є значна схожість особин як рослин, так і тварин у межах популяції. Причому подібність ця стосується лише фенотипу, а за генотипом, як правило, відбувається значне розширення мінливості внаслідок накопичення прихованих мутацій.
На думку І.І. Шмальгаузена, найважливіше значення стабілізуючого добору полягає в стабілізації взаємовідносин організму або популяції з навколишнім середовищем. Отже, ця форма добору не лише зберігає ознаки, але й стабілізує відносини організмів з довкіллям.
Ще одним наслідком дії стабілізуючої форми природного добору є вдосконалення процесів онтогенезу. Це стосується розвитку оптимального фенотипу в певних умовах, що відбувається на основі відбирання різних генотипів шляхом так званої каналізації морфогенезу, яка стабілізує розвиток організмів у певному напрямку внаслідок дії генів-модифікаторів. К. Уоддінгтон та Ф. Добжанський запропонували навіть поділяти стабілізуючий добір на нормалізуючий (зберігає сформовані адаптації) та каналізуючий, під впливом якого вдосконалюється онтогенез.
Зрозуміло, що ідеальної адаптації в природі не буває. Навіть у стабільних умовах подальше вдосконалення організму може відбуватися різними шляхами. У цьому полягає ще один прояв творчого характеру дії природного добору, який поступово розвиває певні ознаки в тривалій історії організмів.
Дію стабілізуючої форми природного добору можна зрозуміти на такому прикладі. Горобці з довшими та коротшими крильми під час бурі загинули: першим занадто довгі крила заважали боротися з вітром, а другим не вистачало сили. Ще один приклад - стабільність частин квітки порівняно з вегетативними органами рослин: пропорції квітки пристосовані до запилення комахами, тому повинні зберігати оптимальні розміри для вдалого запилення.
Щодо стабілізації онтогенезу (каналізуюча дія), то прикладами можуть бути організми з дуже широкою нормою реакції, завдяки якій під впливом певних чинників можуть формуватися різні морфи. Це стосується деяких видів жовтців, стрілолисту, у яких форма листків безпосередньо залежить від особливостей освітлення (глибина занурення рослин). Серед тварин найбільш наочні приклади дають суспільні комахи, яким властивий кастовий поліморфізм. Через особливості годування та вплив певних феромонів відбувається формування особини чітко визначеної касти (в еволюційно досконаліших форм визначення касти зміщується на все більш ранні етапи онтогенезу).
Дизруптивна форма природного добору не завжди визнається за самостійну і може розглядатись як варіант рушійної. І все ж до первинної її можна віднести тому, що дія добору в цьому випадку головним чином спрямовується проти середньої ознаки в популяції. Оскільки ця форма сприяє одночасному розвитку кількох напрямків мінливості (класи фенотипів), то в популяції формується поліморфізм.
У тому випадку, коли різні спрямування дії дизруптивної форми добору зумовлені відмінностями умов довкілля в окремих частинах ареалу (наприклад, клинальна мінливість), то популяції, що їх населяють, набувають стійких генотипних та фенотипічних відмінностей, що має пристосувальне значення. Якщо між такими популяціями припиняється вільне схрещування через певні ізоляційні механізми, то спостерігатиметься подальша їх дивергенція, яка призведе врешті-решт до формування незалежних видів.
Ф. Добжанський запропонував ввести «балансуючий добір», який має дві форми: добір на різноманіття, тобто дизруптивний добір, що формує поліморфізм, та добір на гетерозиготність (гетерозиготизуючий добір), який збільшує генофонд популяції. Урахування останнього дозволяє змінити уявлення про генетичний тягар: він набуває певного екологічного значення, збільшуючи життєздатність гетерозигот.
Прикладами дії дизруптивної форми можуть бути явища поліморфізму в популяціях живих організмів. Це і світла та темна форми двокрапкового сонечка, і різне забарвлення (під колір довкілля) равликів, богомолів, гусені, і навіть кастовий диморфізм, властивий суспільним комахам (морфологічні відмінності тварин різних каст). Але найбільш яскравим прикладом цього можна вважати формування комах з дуже довгими крилами або взагалі без крил на островах з постійними вітрами. В останньому випадку тварини з середньою ознакою (короткі крила) не можуть протистояти дії вітру і гинуть в морі.
Вторинні форми природного добору. До вторинних ми можемо віднести форми природного добору, що виникли в еволюції пізніше, або ж ті, що є складними і можуть бути поділені на кілька первинних. Подібних форм можна запропонувати безмежну кількість, але всі вони так чи інакше зводяться до комплексу первинних (рушійна форма II за Ю.І. Рожковим та О.В. Проняєвим, частотнозалежний добір тощо).
Статевий добір виник в еволюції пізніше, принаймні після формування в тварин різностатевості. Певні положення про цю форму були сформульовані ще Дарвіном, але теорію статевого добору найбільш активно розвивав Л.Ш. Давіташвілі.
У тому випадку, коли відбираються ознаки лише однієї статі, добір називають статевим. Результатом дії такої форми добору є формування статевого диморфізму (рис. 7.2). Еволюційне значення цього явища полягає в тому, що індивід одразу розпізнає стать іншого представника свого виду, завдяки чому може відповідно себе поводити. У тих видів тварин, у яких статевий диморфізм відсутній або виражений порівняно слабо, статевий добір концентрує свій вплив на поведінці, сприяючи формуванню відповідних ритуалів (останні властиві тваринам незалежно від наявності статевого диморфізму). Статевий добір в цілому сприяє прогресивному розвитку виду, хоча в багатьох випадках він може викликати розвиток властивостей, несприятливих для виживання окремої особини.

Рис. 7.2. Приклади статевого диморфізму в птахів (за О.В. Михєєвим, 1995):
а - гага звичайна (Somateria mollissima); б - турухтан (Philomachus pugnax) в парувальному вбранні
Раніше в науці точилися суперечки щодо доцільності виділення статевого добору як самостійної форми (навіть Уолес піддавав сумніву доречність останнього). Але ще Дарвін підкреслював особливості наслідків дії статевого добору для організмів, які не збігаються з іншими формами. Ознаки, закріплені внаслідок дії цієї форми природного добору, є корисними лише протягом незначного періоду - реалізації репродукційного потенціалу. Увесь інший час вони можуть бути байдужими або навіть шкідливими для певного організму, через що зростає ймовірність його загибелі від ворогів чи з інших причин. У той же час, якщо загибель одиниць (навіть багатьох) сприяє ефективному розмноженню популяції в цілому, то це явище СЛІД розцінювати як адаптивне.
Не слід вважати, що статевий добір може діяти незалежно від інших форм природного добору. Більше того, орган, що спочатку сформувався під безпосередньою дією статевого добору, може в подальшому набути нової функції, а орган, який жодним чини не стосувався процесів розмноження, може видозмінитися і набути певного статевого значення. Отже, у формуванні статевих ознак можна виокремити рушійну, дизруптивну та стабілізуючу форми природного добору.
На думку Л.Ш. Давіташвілі, статевий добір відсутній у рослин, протистів і низькоорганізованих безхребетних тварин. У той же час навіть у трилобітів (відомі з раннього кембрію) можна припускати можливість нижчих проявів цієї форми природного добору. Розвиток статевого добору повинен був безпосередньо залежати від удосконалення органів чуття, у першу чергу зору й слуху, а також загального розвитку нервової системи. Це пояснюється тим, що для формування шлюбної поведінки необхідна наявність певних передумов як в отриманні необхідної інформації з довкілля, так і в її переробці та здійсненні адекватної відповіді.
Шляхи, якими статевий добір спрямовує еволюцію великих груп, можуть суттєво відрізнятися, що залежить від загальних особливостей будови, популяційної структури та поведінки відповідних груп тварин. Якщо в багатьох риб і птахів формується яскраве забарвлення, надмірно розвиваються певні анатомо-мор- фологічні структури, то ссавці звичайно мають переважно бліде забарвлення.
В історичному минулому статеві ознаки значно відрізнялися від сучасних форм. У кайнозойських та мезозойських рептилій ці утворення були надзвичайно великими (гребені, вітрила і різноманітні вирости - рис. 7.3), що потребувало значних витрат матеріалу й енергії для їх розвитку. До того ж ці структури були нерухомі або ж зі значно обмеженою рухливістю, через що не здійснювали активних демонстрацій і навряд чи виконували будь-які інші функції. Більш сучасні пристосування (забарвлення пір'я в птахів тощо) виявляються прогресивнішими завдяки легкості, здатності до періодичних змін, значній лабільності, зручності використання та поліфункціональності. Особливого значення набуває поведінка тварин, яка в період розмноження має вигляд спеціальних ритуалів (шлюбна поведінка - рис. 6.3, 7.2).
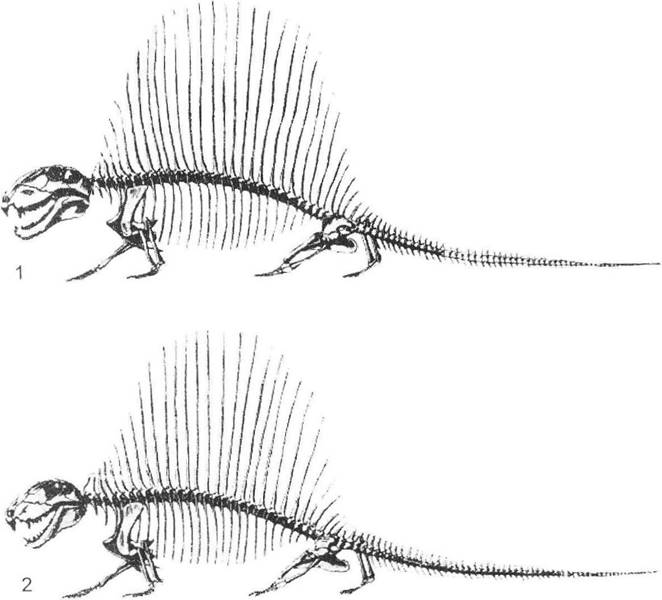
Рис. 7.3. Статевий добір у вітрильних пелікозаврів на прикладі Dimetrodon limbatus (за Ромером та Прайсом (Л.Ш. Давіташвілі, 1961)):
1 - самець (довжина тіла близько 283 см); 2 - самиця (довжина тіла близько 256 см)
Вторинні статеві ознаки є виключно мінливими, що пов'язане з їх прискореним еволюційним розвитком. Пояснював це явище ще Дарвін: формування вторинних статевих ознак багато в чому залежить від надання переваги особині протилежної статі (прояв безпосередніх уподобань конкретних організмів). Зрозуміло, що воно не безмежне й має свою норму реакції. Тому мінливість статевих ознак стосується переважно незначних відхилень при збереженні повноти загальної картини.
Ще більших відмінностей, порівняно з морфологією, набуває шлюбна поведінка різних видів тварин, яка може відбуватися значно швидше за розвиток морфологічних структур. Згідно з Н. Тінбергеном, шлюбна поведінка виконує принаймні чотири додаткові функції: синхронізації, просторової орієнтації, залицяння та репродуктивної ізоляції.
Синхронізація необхідна для одночасного достигання статевих продуктів; залицяння спрямоване на чітке позначення ролі партнерів. Орієнтація в просторі допомагає партнерам знайти одне одного (переважно є функцією приваблення), а репродуктивна ізоляція протидіє схрещуванню з представниками інших видів. Тому шлюбну поведінку не можна зводити лише до турнірів самців за право володіти самицею (у деяких видів змагання відбуваються як між самцями, так і між самицями, а інколи - лише між останніми).
Особливістю шлюбних змагань є те, що вони вкрай рідко набувають жорсткого характеру з загибеллю одного з партнерів. Більшість цих змагань зводиться до демонстрації сили з подальшою втечею слабшого суперника. Подібна тенденція є еволюційно виправданою, оскільки переможений організм не гине, молода особина отримує можливість взяти участь у розмноженні наступного року. Тому статевий добір не стільки знищує, скільки усуває слабші організми від розмноження.
Одним з утворень, які виникають саме під дією статевого добору, можна вважати роги копитних, еволюцію яких докладно розглянуто С.О. Сєверцовим. Сьогодні досить складно визначити напевне, для чого призначалися роги в копитних на початку їх еволюції — боротьби за самицю чи захисту від хижаків. Невеликий зріст та сила, недостатні для нанесення смертельних ударів під час боїв за самицю, що були властиві предковим формам жуйних копитних, свідчать про те, що поява рогів могла бути наслідком дії саме статевого добору. Зі збільшенням розмірів тіла тварини гострі короткі роги (рис. 7.4) стають небезпечними, унаслідок чого їх еволюція була спрямована на перетворення останніх на турнірну зброю.

Рис. 7.4. Форми рогів примітивних оленів (за С.О. Сєверцовим, 1951):
1 - голова Elaphodus; 2 - голова Pudu; 3 - голова Mazama rufa
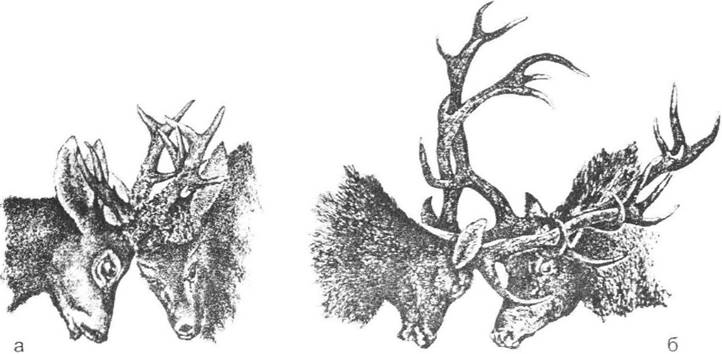
Рис. 7.5. Турніри самців:
а - козулі (Capreolus capreolus); б - оленя благородного (Cervus elapus) (за С.О. Сєверцовим, 1951)
Такі сучасні форми, як лось та олені, захищаються від хижаків копитами, а їх розгалужені роги дозволяють лише боротися за самицю з іншим самцем (рис. 7.5). В оленів, яким властиві часті сутички за самиць, зчеплення суперників розгалуженими рогами замінює укол на штовхання з усією силою корпусу і ніг (крім випадків нерозгалужених рогів - так звані олені- вбивці). До того ж роги щороку скидаються навесні (безгодівний період), а потім знову відростають до нового періоду розмноження. Гострі очні відростки, які спрямовані гачками вперед, але участі в шлюбному турнірі не беруть, можуть використовуватись оленями для захисту від хижаків.
Зовсім інша тенденція розвитку спостерігається для порожнисторогих, у яких роги вкриті роговим покривом і нездатні розгалужуватись. Самці використовують їх не для нанесення удару, а як «шпаги», тому їх вага та розміри мають неабияке значення (рис. 7.6).
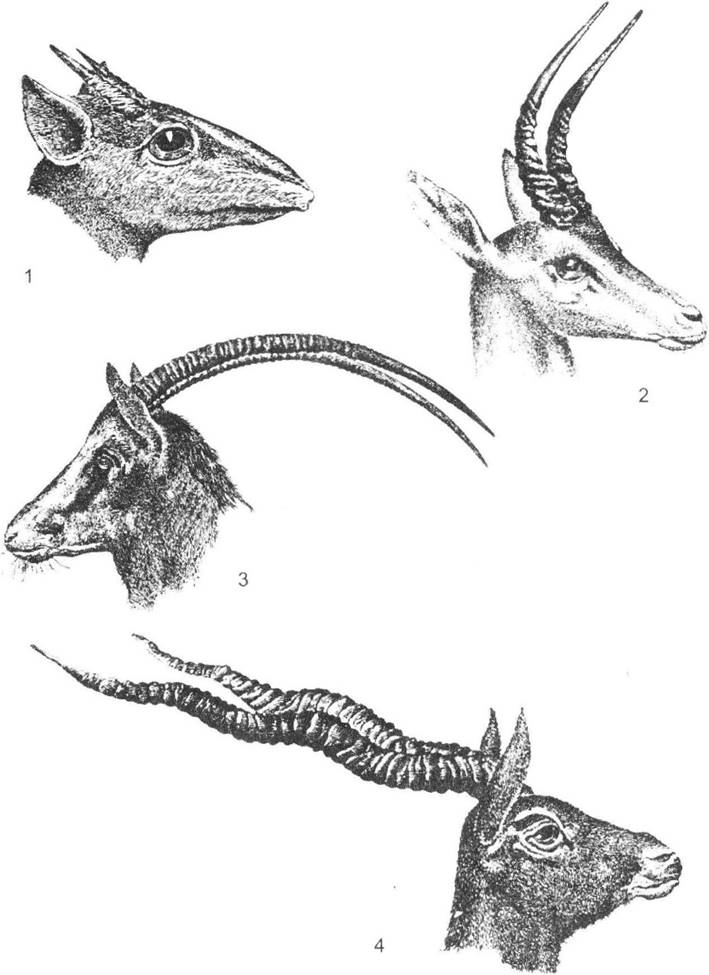
Рис. 7.6. Голови різних видів антилоп (за С.О. Сєверцовим, 1951):
1 - Madoqua·, 2 - Ammodorcas klarkei; 3 - Oryx leucoryx; 4 — Antilopa cervicapra
Великі представники порожнисторогих (наприклад, бики) можуть ефективно використовувати свої роги для захисту від хижаків, у чому виявляється активна дія міжвидової боротьби за існування. У деяких форм (гну) під впливом статевого добору основна частина рогу перетворюється на шолом, який захищає череп тварини від ударів суперників лобом, а кінці залишаються загостреними й зігнутими гачком, завдяки чому підвищується ефективність у захисті від хижаків. У випадку зникнення значного тиску хижаків (особливим прикладом є свійські тварини) серед копитних переважає статевий добір, що зумовлює зміну форми і розташування на черепі рогів (не виходять за площину лоба) і робить укол суперника неможливим.
Груповий добір часто протиставляється індивідуальному як форма природного добору, при дії якого перевагу в розмноженні набувають представники певної групи. Ця форма так само повинна вважатись вторинною, оскільки виникла в еволюції відносно пізно. Через те що груповий добір може відбуватися лише на тлі певних соціальних відносин організмів, то виник він не раніше, ніж субсоціальні спільноти тварин.
Деякі вчені вважають, що ця форма добору є не більше, ніж припущення, оскільки багато в чому вона ґрунтується на альтруїзмі, що до того ж може суперечити загальному уявленню про природний добір. Дійсно, часто внаслідок дії групового добору відбувається закріплення рис, шкідливих для окремого організму, але корисних для групи й виду в цілому.
Одним із прикладів дії є суспільні комахи, у яких здатність до розмноження мають лише одиниці, а інші члени колонії тільки забезпечують нормальну реалізацію їх репродуктивного потенціалу. Вважається, що суспільні комахи виникли через необхідність кращого захисту гнізда, передавання кишкових симбіонтів тощо, тобто в цьому випадку розподіл функцій між членами колонії, який є негативним для окремої особини, має бе- посереднє позитивне значення для еволюції виду в цілому. Несуспільні форми, особливо за умови високої конкуренції, значно поступаються суспільним за всіма показниками.
Еволюція людини ще більш наочно підкреслює наявність групового добору, оскільки постійно відбувалась під його безпосереднім впливом. Усі форми, починаючи з людиноподібних мавп, які не йшли шляхом розвитку суспільства, виявилися приреченими.
І.І. Шмальгаузен, який одним із перших виокремив цю форму добору, розумів її як переважання представників певних угруповань (у першу чергу в розмноженні) — від сімейних груп до надпопуляційних утворень. Незалежно від того, визнаємо ми існування групового добору чи ні, саме він залишається поки що єдиним більш-менш реалістичним поясненням еволюції суспільства як біологічного явища.
Думки вголос
Ця глава завершує виклад загальних положень про механізми макроеволюції. Далі розглядатимуться наслідки, тобто формування адаптацій і, як узагальнений результат усього еволюційного процесу, видоутворення. Незважаючи на те що мікроеволюція є найбільш розробленою частиною синтетичної теорії еволюції, і тут існує необхідність нових узагальнень. У першу чергу це стосується особини. Поки що механізми мікроеволюції ґрунтуються або на доорганізменому (мутації тощо), або ж на надорганізменому рівнях (популяція, хвилі життя тощо). При цьому найбільш важлива одиниця життя - особина - залишається поза увагою вчених. Чи можемо ми примиритися з таким обмеженим підходом до вивчення біологічних явищ?