Основи еволюції - Корж О.П. - 2006
Частина ІІ. МАКРОЕВОЛЮЦІЯ
Розділ 12. Еволюція онтогенезу
12.5. Співвідношення онтогенезу та філогенезу
Згідно з Ф. Мюллером, потомство набуває нового стану, відхиляючись від батьківської форми, у два способи: змінюючи самий цей хід або ж подовжуючи його. У другому випадку нащадки проходять увесь розвиток предків, унаслідок чого історичний розвиток виду відображається в його індивідуальному розвитку. Історичні свідчення, які зберігаються в онтогенезі, поступово стираються, оскільки розвиток все більше набуває вигляду прямого шляху - від яйця до кінцевої форми тварини. Ці зміни можуть значно посилюватись через боротьбу за існування, яку ведуть вільноіснуючі личинки. Швидкість еволюційних перетворень організмів може певним чином відрізнятись, і в цьому випадку в онтогенезі добре зберігатимуться лише ті ознаки, які неодноразово повторювалися в онтогенетичному розвитку предкових форм під час відносної еволюційної незмінності відповідних видів. Початкова історія виду зберігалася в онтогенезі тим повніше, чим менше відхилявся спосіб життя молодих особин від дорослих і чим менше сталося зсувів окремих стадій від пізніших до більш ранніх життєвих періодів та якщо не відбувалось самостійних надбань.
Факти, які відображують повторення в онтогенезі філогенезу, поділяють на три основні групи:
1) відтворення в онтогенезі загального шляху історичного розвитку від простого до складного. Це може полягати в тому, що розвиток в онтогенезі, як і в філогенезі в цілому, починається однією клітиною і завершується складнодиференційованим багатоклітинним організмом;
2) повторення в онтогенезі загальних особливостей будови дорослих предків;
3) повторення в онтогенезі нащадків певних особливостей індивідуального розвитку предків.
Згідно з біогенетичним законом Е. Геккеля, онтогенез є коротким та швидким повторенням (рекапітуляцією) філогенезу, зумовленим фізіологічними функціями спадковості та пристосованості. Підтвердженням цього закону є проходження ембріонами вищих тварин у своєму розвитку стадій, властивих предковим формам. У той же час далеко не всі стадії мають місце, зокрема зародок майже ніколи не набуває вигляду дорослих предкових форм, а лише схожий на їх ембріони. До проблем цього закону слід віднести його односпрямованість: простежується лише залежність онтогенезу від філогенезу тощо.
Карлом Ернстом фон Бером був сформульований закон зародкової схожості, який тривалий час становив теоретичну базу ембріологічних досліджень. У цьому законі Карл фон Бер встановив два важливих явища ембріонального розвитку організмів:
1) характерні ознаки дорослих тварин закладаються в зародка в певній послідовності одна за іншою;
2) послідовність формування ознак (органів) в онтогенезі відповідає послідовності систематичних категорій згідно зі зменшенням їх спільності, тобто філогетичним віком цих ознак (спочатку закладаються найбільш загальні ознаки, а лише потім спеціалізовані). Іншими словами, ознаки великих систематичних груп закладаються раніше, ніж дрібних, філогенетично молодих (рис. 12.12).
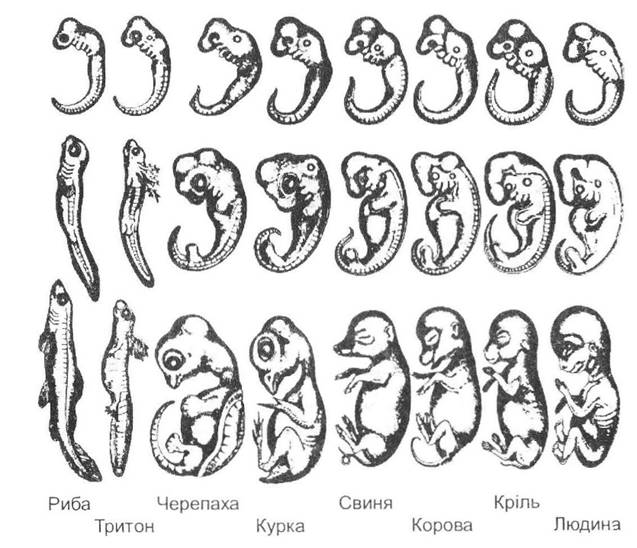
Рис. 12.12. Ілюстрація до закона Бера (за С. Гілбертом, 1993)
О.М. Сєверцов запропонував теорію філембріогенезу - еволюційних змін онтогенезу, які призводять до подальших змін філогенетичного розвитку відповідної групи. Учений виділяє три шляхи подібного перетворення онтогенезу, які можуть викликати відповідні філогенетичні зміни - анаболія, девіація та архалаксис.
Анаболія - це еволюційні зміни розвитку організму на пізніх стадіях (інколи її розглядають як своєрідну надбудову). На думку О.М. Сєверцова, велика кількість органів дорослих організмів розвивається саме шляхом додавання певних стадій на кінцевих етапах морфогенезу відповідного органу. Часто ці процеси супроводжуються певним подовженням онтогенезу, оскільки орган спочатку розвивається нормальним шляхом, а лише
після його формування відбувається додаткове перетворення. Кінцеві стадії морфогенезу предків закріплюються в онтогенезі нащадків, завдяки чому ми можемо спостерігати повторення ознак дорослих предків. Цей тип еволюції органів вважається досить поширеним, а найбільш наочними є приклади розвитку щелеп у саргана (рис. 12.13), плавців у морського півня та морського чорта.
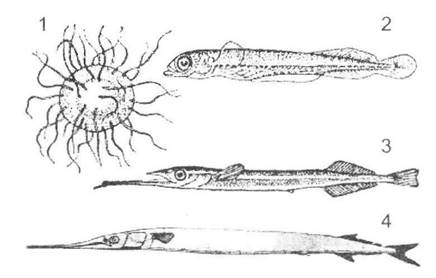
Рис. 12.13. Стадії розвитку саргана (Belone sp.) (за різними авторами): 1 - ікра; 2, 3 - мальок; 4 - доросла риба
Девіація може розглядатись як процес перетворення певних органів на середніх етапах їх формування. Орган, як правило, закладається, як і раніше, але на середніх етапах відбувається відхилення в його розвитку від предкових форм. Як приклад можна навести специфіку розвитку луски в акулових риб та в рептилій (закладання здійснюється аналогічно, а в подальшому відбуваються суттєві зміни - у риб луска костеніє, а в рептилій - роговіє).
Архалаксис вважається процесом зміни розвитку органа на ранніх етапах або в період його закладання. Зміна самих зачатків органів майже завжди супроводжується змінами і всього їх морфогенезу (рис. 12.14). При подібному типі еволюції тривалість розвитку не збільшується, а рекапітуляцій в подальшому онтогенезі нащадків не спостерігається. За типом архалаксису утворюються переважно нові, раніше відсутні ознаки або функціональні зв'язки між органами. Цей процес також вважається досить поширеним: формування видовженого тіла в змій (збільшується кількість хребців з 80 до 250-300 переважно за рахунок тулубових при збереженні майже незмінної кількості хвостових хребців), закладення лише трьох пальців у сучасного коня (два з них перетворюються на грифельні кістки), зміна кількості зубів тощо.
Згідно з доказами О.М. Сєвєрцова, повторення в онтогенезі предкових форм можливе лише у випадку анаболії, в інших варіантах перетворення онтогенезу повернення до попереднього стану неможливе. Дослідник пропонує виділяти позитивну та негативну анаболію, девіацію та архалаксис (у цілому ним запропоновано 12 модусів філембріогенезів), що не набуло загального визнання.
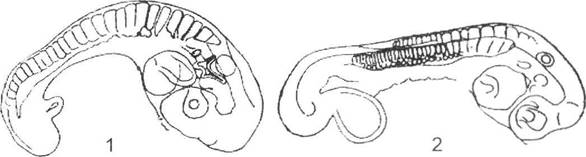
Рис. 12.14. Приклад архалаксисе (за О.М. Сєверцовим, 1945):
1 - зародок гекона з 28-тулубовими сомітами; 2 - зародок вужа з 34-тулубовими сомітами
На сьогодні значна кількість учених не лише не погоджується з теорією філембріогенезу, а взагалі не визнає біогенетичного закону через значну кількість невідповідностей у розвитку представників певних груп (наявність багатьох винятків позбавляють закон сенсу). У той же час вони згодні з фактом взаємозв'язку між онтогенезом та філогенезом, часто зосереджуючи свою увагу лише на явищі мутацій та їх подальшому впливі на результати онтогенетичного розвитку і, як наслідок, філогенетичних змін відповідних груп (уявлення про тератологічні зміни - формування багатообіцяючих виродків, педоморфоз, гормональні зсуви, онтомутації, макромутації тощо).
Слід зазначити, що ще Ч. Дарвін та Ф. Мюллер показали: еволюційні зміни в онтогенезі можуть відбуватися на будь-яких стадіях, що й викликає більші або менші відхилення від властивого предковим формам розвитку.
Виникають вони внаслідок формування нових особливостей або пересування наявних структур у просторі чи в часі. При цьому не завжди більш ранні зміни викликають більше відхилень у подальшому розвитку групи.
Думки вголос
Явище онтогенезу настільки багатогранне й різноманітне, що в такому короткому огляді не можна охопити всі найважливіші його аспекти. Незважаючи на відсутність загальновизнаної теорії онтогенезу, слід погодитись, що саме індивідуальний розвиток є одним із найунікальніших біологічних явищ, який виступає підґрунтям для здійснення багатьох інших, у тому числі й еволюційних процесів. Неодностайність думок ще раз підкреслює необхідність подальших пошуків, але й застерігає від догматичного наслідування будь-яких наукових авторитетів.