Основи еволюції - Корж О.П. - 2006
Частина ІІ. МАКРОЕВОЛЮЦІЯ
Розділ 13. Особливості здійснення філогенезу
13.1. Форми філогенезу різних груп
Напрямки еволюції організмів визначаються, з одного боку, тим середовищем, у якому вони розвиваються, а з іншого - особливостями будови самого організму (його попереднім філогенезом).
Філогенез - це історичний розвиток певних груп організмів, еволюція органічного світу в цілому, окремих його систематичних груп тощо.
Філогенез може розвиватись у вигляді первинних та вторинних форм: до первинних відносять філетичну еволюцію та дивергенцію, які виступають основою для інших еволюційних перетворень, а серед вторинних розрізняють конвергенцію та паралелізм (рис. 13.1).
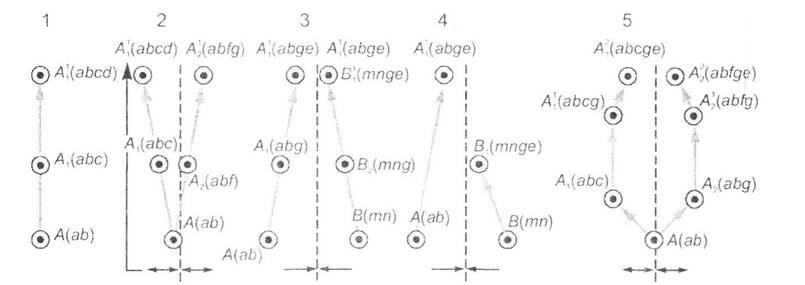
Рис. 13.1. Схеми різних форм філогенезу (за О.В. Яблоковим, А.Г. Юсуфовим, 1989):
1 - філетична еволюція; 2 - дивергенція; 3,4 - синхронна та асинхронна конвергенції; 5 - паралелізм
Філетична еволюція - це зміни одного виду на інший (в одному філогенетичному напрямку) без урахування можливих відгалужень, тобто зсув середніх ознак, властивих певній групі. Подібні перетворення стають основою для всіх інших варіантів еволюції груп і тому можуть бути виділеними майже для кожного виду.
Зауважимо, що не всі дослідники дотримуються думки щодо необхідності виділення цієї форми філогенезу. З одного боку, він розглядається як варіант видоутворення (анагенез), а з іншого - не може тривати для певної групи нескінченно довго й на певному етапі передбачає подальший перехід до дивергенції або інших форм філогенезу. На це слід зазначити, що еволюція ніколи не стоїть на місці, підтвердженням чому і є філетична еволюція. Навіть у відносно стабільних груп (наприклад, філогенетичні релікти) спостерігається поступова зміна форм у часі. Якщо розвиток без поділу групи на кілька нових триває недовго, то немає підстав відмовлятись від пропонованого терміна, який дозволяє краще зрозуміти загальну спрямованість еволюційних перетворень певних видів або більших систематичних груп у ході їх пристосувань до певних умов існування.
Як приклад можна навести палеонтологічні ряди добре досліджених груп організмів. Найвідомішим є філогенетичний ряд коней — найбільш спеціалізованих сучасних представників непарнокопитних тварин. Розвивалася ця гілка переважно в Північній Америці, звідки мали місце постійні переселення її представників до Європи. Сучасний кінь після переселення повністю вимер в Америці. Вихідною формою, що дала початок всій гілці коней, була невеличка лісова форма (фенакодус), яка живилась м'яким листям дерев. Третинні похолодання та зменшення кількості лісів з подальшим формуванням відкритих просторів зумовили поступовий перехід предків коней до степових умов існування.
Такий перехід не міг відбутися без значних перебудов організації, що торкалося відразу кількох структур. У першу чергу існування на відкритих просторах супроводжувалося необхідністю втечі від природних ворогів, що призвело до поступового збільшення розмірів та перетворення початкової п'ятипалої кінцівки спочатку на трипалу (еогіпус, мезогіпус, міогіпус та інші форми), а в подальшому й на однопалу (пліогіпус, сучасний кінь - рис. 13.2).
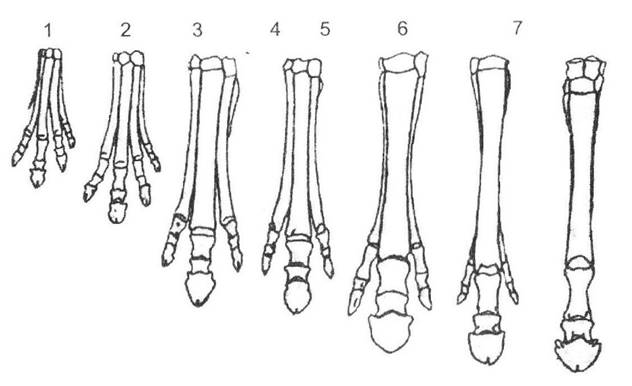
Рис. 13.2. Еволюція кінцівки коней (за Г.Ф. Осборном із І.І. Шмальгаузена, 1935):
1 - Eohippus (н. еоцен); 2 - Orohippus (сер. еоцен); 3 - Mesohippus (н. олігоцен); 4 - Miohippus (в. олігоцен); 5 - Hypohippus (сер. міоцен);
6 - Нірраііоn (в. міоцен - н. пліоцен); 7 - Pliohippus (в. міоцен - н. пліоцен)
Водночас відбувається редукція малої гомілкової кістки та її приростання до великої, а також редукція нижньої частини ліктьової кістки та зростання її верхньої частини з променевою (процеси олігомеризації, пов'язані з посиленням основної функції відповідних органів).
Значні перетворення торкнулися також будови зубної системи цих тварин (рис. 13.3). Перехід до живлення травою (значно жорсткіша їжа порівняно з листям дерев, особливо сіно та насіння злаків) зумовив необхідність зменшення стирання зубів (формується гіпсодонтний тип корінних зубів). Наявність високих бугрів та гребенів, укритих цементом, дозволяє при стиранні коронки на будь-якому рівні залишати не лише крихкий цемент, але й кілька прошарків твердої емалі, що значно подовжує час слугування зубів.
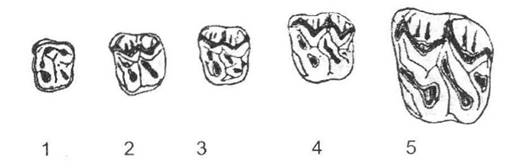
Рис. 13.3. Перетворення верхніх корінних зубів коней (за Г.Ф. Осборном із І.І. Шмальгаузена, 1945):
1 - Hyracotherium; 2 - Pachynolophus; 3 - Anchilophus; 4 - Mesohippus; 5 - Anchitherium
Іншим добре відомим прикладом філетичної еволюції можна вважати історію розвитку слонів, які розвивалися як вузько- спеціалізовані форми. Найцікавішими змінами слід вважати перетворення зубної системи, у якій друга пара різців набуває гіпертрофічного розвитку і формує врешті-решт бивні. Поступове скорочення щелепи та ускладнення структури жуйних зубів до лофодонтного типу (рис. 13.4) - великі за розмірами жуйної поверхні з наявністю багатьох поперечних складок в азіатського слона (Elephas maximus) - до 27, призвело до так званої горизонтальної зміни зубів. Вона полягає в тому, що задні зуби прорізаються лише поступово, відповідно до зношування попередніх, функціонально та морфологічно замінюючи їх. У сучасних слонів водночас нормально функціонує лише один корінний зуб з кожного боку щелепи.
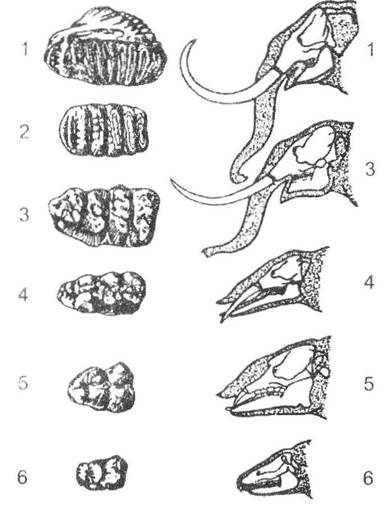
Рис. 13.4. Еволюція корінних зубів та голови в ряду хоботних
(за І.І. Шмальгаузеном, 1945): 1 - Elephas (сучасний вид); 2 - Stegodon (пліоцен); 3 - Mastodon (дилювій);
4 - Trilophodon (міоцен); 5 - Palaeomastodon (н. олігоцен); 6 - Moeritherium (в. еоцен)
Вихідною формою для слонів вважається еоценовий меритерій (рис. 13.5), який мав майже повну зубну систему та нагадував невеликого за розмірами (до 1 м) тапіра. У палеомастодонта спереду залишаються лише бивні, які у верхній щелепі більші, а нижня щелепа сама є дуже видовженою; очевидно, цим формам був властивий уже досить великий хобот. У подальшому сталося розділення гілки слонів принаймні на дві групи: у динотерій почали переважати нижні бивні (верхні поступово редукувалися), а в мастодонтів та інших слонів, навпаки, розвинулися верхні. Лише в гілки, яка призвела до формування справжніх слонів, сформувався лофодонтний тип кутніх зубів. Відомі й інші приклади подібних філогенетичних рядів.
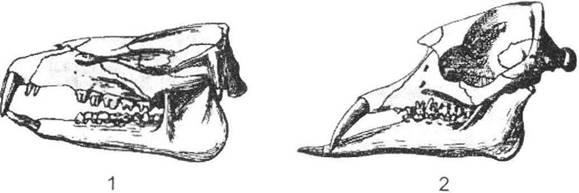
Рис. 13.5. Черепи примітивних хоботних (за Ендрю із І.І. Шмальгаузена, 1945):
1 - Moeritherium (в. еоцен); 2 - Palaeomastodon (н. олігоцен)
Дивергенція - розходження форм (або ознак) з формуванням гомологічних структур через відмінності в умовах існування спочатку єдиної групи (або споріднених груп). Ще Ч. Дарвін передбачав здійснення еволюції саме шляхом поступового розходження ознак з вимиранням проміжних, менш пристосованих форм. Чим більші відмінності середовища існування цих дочірніх груп і чим довше триває процес розходження, тим глибше виражена відповідна дивергенція.
Наочним результатом дивергентного розвитку організмів можна вважати появу гомологічних органів (рис. 11.1), які мають спільне походження, але є наслідком пристосування організмів до різних умов існування. І.І. Шмальгаузен наводить такий приклад дивергенції: якщо відмінності між річковою та озерною фореллю або різними видами жаб незначні, то розходження ознак значно зростає в таких споріднених груп, як видра та куниця, білка та ховрашок (різні умови існування), і ще більше відмінностей в їжака та кажана, слона та сирени, птаха й крокодила.
Прикладом дивергенції можна вважати також розвиток копитних (кондилартри) та хижих форм (креодонти), які на початкових стадіях еволюції ледь розрізнялися між собою. Існує думка, що копитні походять від певної предкової групи, але в їх розвитку мала місце значна кількість паралелізмів. Тому сучасна корова має приблизно однакову спорідненість як з конем, так і з тигром або будь-яким іншим хижаком. Вважається, що саме дивергенція надає необхідну базу для розвитку основних систематичних груп.
Конвергенція - це набуття схожих рис організації систематично далекими формами, які потрапили в наближені умови існування. Під неспорідненими формами в цьому випадку слід розуміти організми, пов'язані віддаленою формою спорідненості (певного мірою між собою пов'язані родинними зв'язками всі організми). Особливістю конвергентного розвитку є формування аналогічних органів (функціонально схожі, але мають принципово різну організацію). Чим специфічніші вимоги до організму збоку середовища його існування і чим триваліші та глибші пристосування до цього середовища, тим яскравіше виражена конвергенція (рис. 13.6). Особливості конвергенції полягають у тому, що схожість торкається лише зовнішніх ознак, не поширюючись на структуру, і є вторинною, тоді як дивергенція базується на глибокій схожості організації і вторинних відмінностях, зумовлених специфікою середовища існування.
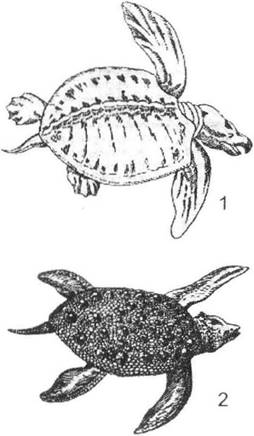
Рис. 13.6. Конвергентна схожість морських черепах (ряд Chelonia) з викопними плоскозубими рептиліями (за І.І. Шмальгаузеном, 1945): 1 - Archelon (в. крейда); 2 - Placochelys(тріас)
Так, прикладами схожості, яка спирається на конвергенцію, є редукція очей у печерних тварин, розвиток ластів у водних рептилій та ссавців. Конвергентну схожість цілих організмів вбачаємо в обтічній формі тіла акули, костистої риби, рептилії (іхтіозавр) та ссавця (дельфін). Конвергентними формами слід вважати й більш споріднених організмів: розвиток довгих кінцівок у журавлів та лелек, схожість між сумчастим кротом та звичайним кротом, опосумом та видрою, сумчастим літуном, білкою-летягою та шерстокрилом. У цьому разі також маємо справу з неспорідненими формами, оскільки їх схожі ознаки надбані організмами незалежно ще на початкових фазах їх розвитку. Так, у карети (Eretmochelys) та шкірястої черепахи (Dermochelys) колись були спільні предки, але ці суто наземні черепахи розвивалися дивергентно ще на суходолі, а їх подальший перехід до водного середовища та розвиток спільних ознак слід вважати наслідком конвергентного розвитку.
Паралелізм - явище паралельного набуття схожих структур у близькородинних видів тварин. І.І. Шмальгаузен визначає паралелізм також як конвергенцію споріднених форм. Проблемним залишається при цьому відносність визначення ступеня такої спорідненості. У споріднених форм провести чітку межу між паралелізмом та конвергенцією, а як наслідок - між аналогічними та гомологічними органами, майже неможливо. У той же час використання самого терміна «паралелізм» визнається доцільним, оскільки він позначає незалежний розвиток схожих пристосувань на базі гомологічних структур. Таким чином, паралелізм начебто складається з двох процесів: попередньої дивергенції та подальшого паралельного розвитку спільних ознак у схожих умовах існування відповідних організмів.
Якщо дві споріднені групи організмів розвиваються паралельно за багатьма ознаками, то очевидна паралельна еволюція цих форм.
Прикладами паралельного розвитку можна вважати дві найважливіші гілки динозаврів - ящеротазових та птахотазових, у яких паралельно відбувалося збільшення розмірів тіла, повернення до ходіння на чотирьох кінцівках, а також формування великої кількості інших спільних ознак. Паралельно розвивалися також обидві гілки птерозаврів - птеродактилі та рамфоринхи, які перейшли від плануючого до справжнього польоту, паралельно набуваючи таких ознак, як пневматизація скелету, розвиток крил, грудини тощо. З іншого боку, при розгляді птерозаврів та птахів слід говорити лише про конвергенцію, оскільки ці пристосування розвивалися в них абсолютно незалежно і дещо відмінними шляхами (їх крила слід вважати аналогічними органами).
Паралельним можна вважати розвиток зубатих та беззубих китів; тюленів, моржів і котиків; однопалих форм коней (Північна Америка) та літоптерн (Південна Америка) тощо. На прикладі паралелізму пристосувань копитних (рис. 13.7) простежується потреба в мірі перебудови організації. Так, літоптерни швидше розвинули однопалі кінцівки, але при цьому розміри середнього пальця в них майже не змінилися. Коні еволюціонували повільніше, але адаптивні надбання супроводжувались глибокими перебудовами всієї організації цих тварин - збільшення розмірів пальця, скоординоване зростання розмірів тіла тощо. Вважається, що саме через глибшу модифікацію всієї організації коні після об'єднання Америк швидко витіснили з еволюційної арени літоптерн як більш прогресивні форми.
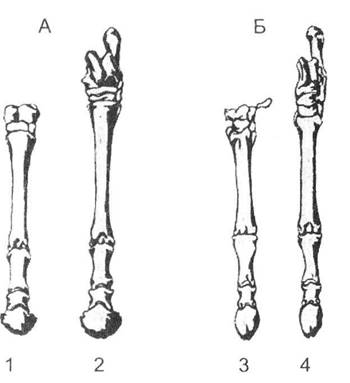
Рис. 13.7. Порівняння будови передніх (1) та задніх (2) кінцівок різних копитних (за Дж. Сімпсоном, 1983):
А - сучасний кінь (Equus); Б - ранньо-міоценовий літоптерн (Thoatherium)
Інколи розрізняють явище синхронного та асинхронного паралелізму. Прикладом першого з них вважається розвиток однопалих форм серед копитних (відбувався майже одночасно). Асинхронний паралелізм передбачає розвиток подібних структур У різних форм, які відокремлені в часі (вважається, що шаблезубість виникала в котячих принаймні чотири рази в різний геологічний час - рис. 13.8). Але при використанні подібного підходу можна говорити й про синхронну та асинхронну конвергенцію, оскільки розвиток обтічної форми тіла в дельфінів ніяк не міг відбуватися синхронно з іхтіозаврами та іншими попередниками.
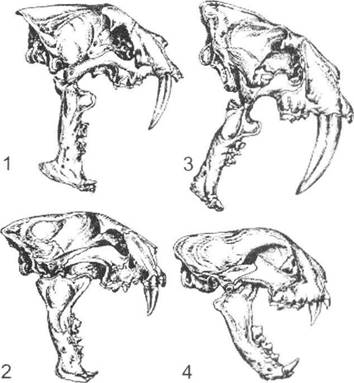
Рис. 13.8. Асинхронний паралелізм на прикладі розвитку в котячих шаблезубості (за А. Ромером з О.В. Яблокова, А.Г. Юсуфова, 1989): 1 - Hoplophoneus (олігоцен); 2 - Dinictls(олігоцен); 3 - Smilodon (пліоцен); 4 — Pseudoaelurus (плейстоцен)
У цілому можна вважати, що паралельний розвиток спочатку був властивий усім спорідненим формам, які потім дивергували. Це майже загальне правило за винятком подальшого переходу цих груп до існування в різко відмінних умовах. У той же час на практиці без докладного історичного вивчення відповідних форм досить складно розрізнити явище паралелізму від інших форм еволюції груп.