Основи еволюції - Корж О.П. - 2006
Частина III.ШЛЯХИ РОЗВИТКУ ЖИТТЯ
Розділ 19. Основні напрямки розвитку живих організмів
19.2.Вихід рослин на суходіл
Однією з імовірних причин виходу на суходіл могла бути зростаюча конкуренція за певні фактори (наприклад - за світло). Нижчі рослини, які першими вийшли з водного середовища, не мають стабілізованого водного обміну, унаслідок чого в засушливі періоди впадають у стан анабіозу.
Наземні умови існування значно відрізняються від умов життя у воді: на суходолі вищі рослини одночасно існують у двох принципово відмінних середовищах: наземні частини - в атмосфері, а підземні - у ґрунті.
Повітряне середовище характеризується значно більшим вмістом кисню, ніж водне, а в ґрунті умови мінерального живлення та водний режим (постійний водообмін між рослинами і навколишнім середовищем) є іншими. Тому перехід предків вищих рослин в ці нові умови існування міг відбутися лише після набуття ними спеціальних пристосувань.
Одним із перших, а може, й головним пристосуванням вищих рослин до суходільних умов існування слід вважати появу епідерми (епідермісу) — особливого захисного шару, який складається зі щільно зімкнених клітин покривної тканини (рис. 19.2). Вона запобігає висиханню рослин, негативному впливу температурних коливань, проникненню всередину різних хвороботворних організмів тощо. Виникла епідерма шляхом відповідного перетворення та спеціалізації зовнішніх клітин тіла рослини. їх поверхня в більшості вищих рослин вкрита особливою захисною плівкою - кутикулою, яка стійка до хімічного впливу і має водо- відштовхуючі властивості. Якби шар кутикули був безперервним, це позбавило б рослини можливості газообміну з середовищем і викликало б їх загибель. Тому в процесі еволюції виникають спеціальні дрібні утворення - продихи, через які відбувається інтенсивна дифузія водяної пари, кисню та вуглекислого газу. Наявність продихів є однією з основних рис вищих рослин, починаючи з риніофітів. Ні продихів, ні епідерми майже немає в покривних тканинах коренів та у вторинноводних рослин, що слід розглядати як результат редукції останніх.
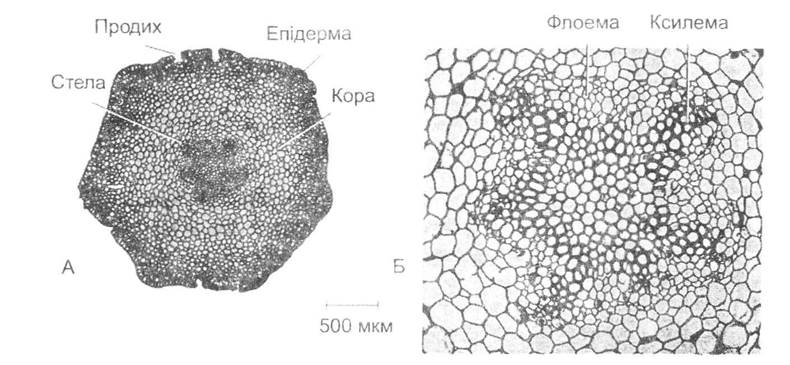
Рис. 19.2. Стебло Psilotum nudum (з П. Рейвна, Р. Еверта та С. Айкхорна, 1990):
А - поперечний зріз стебла; Б — протостела
Виникнення покривної тканини було не єдиною умовою переходу рослин до наземного існування. Організація епідерми відповідає двом протилежним вимогам: з одного боку, запобігати висушуванню тіла рослини, а з іншого - дозволяти повітрю з вуглекислим газом проникати до зелених клітин з хлоропластами. У той же час продихи не лише пропускають вуглекислий газ, а й дозволяють вільно випаровуватися воді в процесі транспірації. Тому вже на початку еволюції вищих рослин виникає необхідність у підтриманні їх водного режиму на відповідному рівні.
Вищі рослини стали відносно незалежними від коливань рівня вологості в ґрунті й атмосфері після набуття здатності стабілізувати вміст води у своєму тілі. Вирішення цієї проблеми викликало утворення ксилеми, яка спеціально пристосована до виконання функцій проведення води. З'являються і флоемні елементи, але аналогічні структури були ще у водоростей, тому флоема вважається значно старшою в еволюційному плані, ніж ксилема.
Із збільшенням розмірів організму змінюється співвідношення поверхні та об'єму, для підтримання якого на оптимальному біологічному рівні відбувається розчленування рослинного організму. В. Ціммерман запропонував теломну теорію походження органів сучасних вищих рослин, яку нині підтримують майже всі ботаніки. Згідно з цією теорією, органи більшості рослин сформувались із сукупності теломів риніофітів шляхом сплющення, переверши- нювання, зростання та редукції теломів (рис. 19.3).
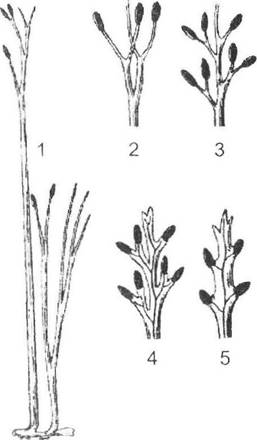
Рис. 19.3. Ринія (Rhynia mayor) (1) та схема виникнення спорофіла папороті з телому типа ринії (2-5) (І.М. Григора та ін., 2004)
Ці процеси викликали диференціацію спорофіта (нестатеве диплоїдне покоління) та розподіл функцій між його частинами, а також утворення особливого органу, де містяться спори, - спорангія, що розвинувся шляхом видозміни кінцівок гілок. Його стінка стає стерильною, захищає внутрішній вміст від висихання.
На суходолі розмноження спорами набуває особливого значення, оскільки переважна більшість їх потрапляє в умови, які не підходять для проростання.
На відміну від водного середовища оболонка спор стає жорсткою, малопроникною для води й виконує захисну функцію. Як наслідок, у рослин в суходільних умовах виникає необхідність утворення великої кількості спор, що потребує достатнього накопичення органічних речовин і відповідного збільшення розмірів спорофіта.
Водночас із формуванням захисної стінки спорангію виникає проблема вивільнення спор та їх подальшого поширення в довкіллі. У риніофітів розсіювання спор спочатку відбувалося після розтріскування стінок спорангію, але пізніше з'являють ся спеціальні механізми, що поступово вирішує проблему нестатевого розмноження (рис. 19.4).

Рис. 19.4. Спорангії та спори (з І.В. Гончаренко, 2004):
А - риніофіти; Б - різноспорові плауноподібні; В - хвощеподібні; Г - рів носпорові папоротеподібні; 1 - термінальний спорангій; 2 - стінка спорангію; 3 - спорогенна тканина; 4 - мікроспорангій; 5 - мега-спорангій; 6 - мікроспора; 7 - мегаспора; 8 - спорангії на спорангіофорах; 9 - спора з флатерами; 10 - маргінальні спорангії; 11 - спорангій
Еволюційні процеси зачепили внутрішні структури рослин: відбувався розвиток тканин, що були в риніофітів, і формування нових. Удосконалювалися провідні системи, найрозвинутішим елементом яких вважаються судини (судинні рослини). Через необхідність підтримання вертикального положення тіла в рослин розвиваються склеренхімні волокна стебел, формуються деревинні та луб'яні волокна тощо. Поступово з'являється камбій, що забезпечує вторинний ріст та потовщення стебла, гілок і коріння.
У водоростей, які живуть у водному середовищі, не існує проблеми з заплідненням жіночих статевих клітин (яйцеклітин), оскільки чоловічі гамети здатні вільно рухатись. З виходом рослин на суходіл виникає потреба в захисті статевих органів від висихання та забезпеченні статевого процесу в нових умовах. Гаметангій у цих умовах зазнає певних видозмін: зовнішній шар стінки стерилізується і, як у спорангії, виконує захисну функцію (відбувається формування антеридіїв - чоловічих та архегоніїв - жіночих репродуктивних органів — рис. 19.5).
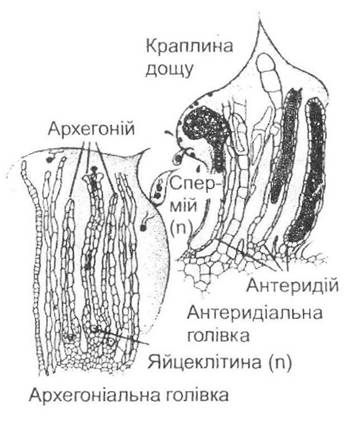
Рис. 19.5. Будова антеридіїв та архегоніїв, а також процес запліднення на прикладі бриївого моху (з П. Рейвна, Р. Еверта та С. Айкхорна, 1990)
І все ж формування зовнішнього захисного шару гаментангіїв не вирішує всіх проблем статевого розмноження, оскільки рух сперматозоїдів можливий лише за умови наявності води. Таким чином, постає проблема поєднання нерухливого способу життя та процесу запліднення статевих клітин, що суттєво стримує поширення вищих рослин суходолом. Наслідком цього стає зменшення ролі гаметофіту та його поступова редукція в переважної більшості вищих рослин.
Вважається, що всі вищі рослини виникли з якихось водоростей (рис. 19.6). Доказом цього є схожість риніофітів - найбільш давніх, давно вимерлих представників вищих рослин - з водоростями, особливо в типах їх розгалуження. Близькими є будова й функції хлоропластів, чергування поколінь вищих рослин та багатоклітинних водоростей, наявність джгутиків та здатність до самостійного плавання чоловічих статевих клітин багатьох вищих рослин.
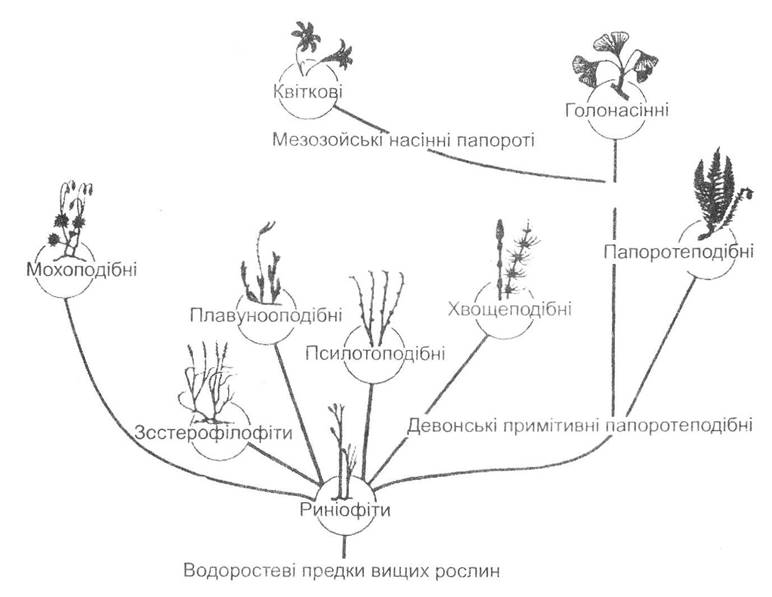
Рис. 19.6. Схема еволюційних зв'язків відділів вищих рослин (за І.В. Грушвицьким та ін., 1978)
Припускається, що вищі рослини походять, скоріш за все, від прісноводних або солонуватоводних зелених водоростей. Докази на користь цієї гіпотези такі: головним фотосинтезую- чим пігментом в обох групах є хлорофіл А, а допоміжним — хлорофіл В та каротиноїди; целюлоза є найважливішим компонентом клітинної оболонки; крохмаль відкладається в хлоропластах, а не в цитоплазмі, як в інших водоростей тощо. У водоростей спостерігаються найрізноманітніші співвідношення диплоїдної та галоїдної фаз життєвого циклу. Але у водоростевого предка вищих рослин, можливо, диплоїдна фаза була розвинена краще, ніж гаплоїдна.
Найбільш давнім та примітивним відділом вищих рослин є риніофіти, які виникли в силурі й проіснували до пізнього девону. Завдяки надзвичайній простоті своєї організації деякі риніофіти зовнішньо були більше схожі на водорості, ніж на вищі рослини. Вони характеризувалися розташуванням спорангіїв на верхівках пагонів, відсутністю кореневої системи та листя, дихотомічним або несправжньомоноподіальним розгалуженням спорофіта, примітивною анатомічною будовою та рівноспоровістю (утворення в рослин спор, однакових за розміром та фізіологічними особливостями).
їх провідна система була слабо розвинена і являла суцільну масу провідної тканини з центральним ядром ксилеми, яке повністю оточене флоемою. Це найбільш примітивний тип стели - центрального циліндру. Ксилема складається з трахеїд; на відміну від більшості судинних рослин у риніофітів механічні (опорні) тканини відсутні. Уже для цих рослин характерною була наявність невеликої кількості продихів простої будови: продих складався лише з двох замикаючих клітин та продихової щілини. Оскільки меристема розташовувалася лише на верхівках пагонів, риніофіти ще не мали здатності до вторинного росту.
Спорангії в риніофітів були товстостінними, різної форми та розмірів. У них відбувається розвиток численних спор, типових за своєю будовою для вищих рослин, але ще не було спеціальних пристосувань для розкриття спорангіїв або вони були дуже примітивними. У ринії спорангії ззовні ще практично не відрізнялися від верхівок звичайних пагонів.
Відсутність у палеонтологічних відкладеннях гаметофітів цих рослин є причиною недостатньо чітких уявлень про їх організацію. Виходячи з даних порівняльної анатомії гаметофітів сучасних рослин учені припускають, що гаметофіт риніофітів був добре розвинений, переважно підземний, мікоризний (симбіоз вищих рослин з грибами), дихотомічно розгалужений, з провідною системою, але питання про його будову залишається поки що відкритим.