ГЕНЕТИКА - Підручник - А.В. Сиволоб - 2008
РОЗДІЛ 1. Природа генетичного матеріалу
РЕПЛІКАЦІЯ ДНК
Комплементарне спарювання нуклеотидів у складі подвійної спіралі ДНК негайно вказує на механізм копіювання генетичної інформації шляхом реплікації. Синтез ДНК відбувається при реплікації з використанням обох полінуклеотидних ланцюгів як матриць - за так званим напівконсервативним механізмом: дві дочірні моле- кули-копії містять один материнський ланцюг (що служив матрицею) і один ланцюг, синтезований de novo. Включення нуклеотидів до ланцюга, що синтезується, детермінується матрицею за принципом комплементарності. Базові молекулярні механізми реплікації є спільними для всіх організмів.
Зростання ланцюга ДНК відбувається в напрямку від 5'- до З'-кінця. Субстратами реакції є 3'-кінцева ОН-група дезоксирибози зростаючого ланцюга та дезоксирибонуклеозидтрифосфати (рис. 1.16). Фермент, що каталізує цю реакцію, - ДНК-залежна ДНК-полімераза.

Рис. 1.16. Схема ДНК-полімеразної реакції
Реплікація ДНК починається з невеликої ділянки - ориджину (origin), де здійснюється ініціація процесу, головним моментом якої є розходження ланцюгів ДНК. Далі з ходом реплікації такий реплікативний міхур (рис. 1.17) розростається у двох протилежних напрямках. На кожному боці міхура існує так звана реплікативна вилка, в основі якої й відбувається синтез ДНК.
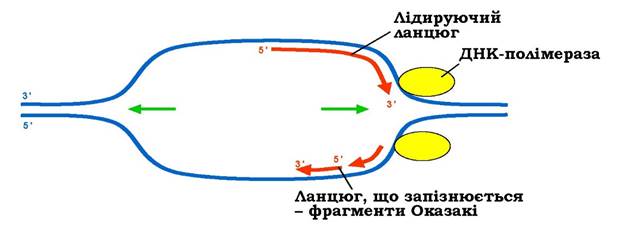
Рис. 1.17. Реплікативний міхур - дві реплікативні вилки, які переміщуються у протилежних напрямках (ланцюги, що синтезуються, показано тільки для однієї з них)
У кожній реплікативній вилці працюють дві молекули ДНК-полімерази, що здійснюють синтез двох полінуклеотидних ланцюгів. Оскільки два ланцюги є антипаралельними, а синтез здійснюється тільки в напрямку від 5'- до З'-кінця, то синтез тільки одного з ланцюгів може відбуватися (і відбувається) безперервно, починаючись від ориджину (рис. 1.17). Цей ланцюг називають лідируючим, його З'-кінець розташований поблизу від основи реплікативної вилки. Синтез іншого ланцюга розпочинається від реплікативної вилки: синтезуються окремі фрагменти - так звані фрагменти Оказакі, які пізніше з'єднуються між собою. Для синтезу кожного з фрагментів треба спочатку звільнити певний простір на матричному ланцюзі - пересунути реплікативну вилку вперед (рис. 1.17); відповідно, фрагментарний ланцюг називають ланцюгом, що запізнюється. Середня швидкість реплікації на одну реплікативну вилку становить ~750 нуклеотидів за секунду в бактерій, 60-90 нуклеотидів за секунду в еукаріотів. Синтез бактеріальної хромосоми відбувається за ~50 хв, повна реплікація ДНК еукаріотичної клітини - за кілька годин.
Ділянку ДНК, де здійснюється реплікація, яка розпочинається з однієї точки, називають репліконом. Бактеріальна хромосома часто містить тільки один ориджин (зокрема, в E. coli) - являє собою єдиний реплікон. У деяких бактерій може бути два реплікони на хромосому. Еукаріотична хромосома є полірепліконом - містить велику кількість точок ініціації. Загалом геном, наприклад ссавців, містить близько 40 тис. ориджинів. Розмір еукаріотичного реплікона варіює від 50 до 200 тис. пар основ, що збігається з розмірами петельних доменів хроматину. Отже, хроматинова петля - це один реплікон, а ориджин збігається з ділянкою, асоційованою з ядерним матриксом. Сусідні реп- лікони еукаріотичної хромосоми врешті-решт "зустрічаються", унаслідок чого утворюються дві копії ДНК хромосоми.
Більшість ДНК-полімераз мають дві ферментативні активності: власне полімеразну, за рахунок якої до З'-кінця ланцюга, що синтезується, приєднуються нуклеотиди, і 3’-екзонуклеазну, яка використовується для редагування помилок - відщеплення помилкових нуклеотидів, щойно приєднаних до 3'-кінця. ДНК-полімераза є прецизійним молекулярним пристроєм: її полімеразний активний центр забезпечує впізнання комплементарного нуклеотиду в складі матриці нуклеозидтрифосфатом, приєднує цей черговий нуклеотид до зростаючого ланцюга (рис. 1.16) і пересувається на один нуклеотид уперед уздовж матриці, знову повторюючи вказані операції з наступним нуклеотидом. При цьому частота помилкового включення нуклеотидів забезпечується на рівні ~10-5. Але оскільки ДНК синтезується "раз і назавжди" перед її передачею нащадкам, такий рівень помилок не може вважатися задовільним. Якщо внаслідок приєднання помилкового нуклеотиду утворилася некомплементарна (тобто нестабільна) пара основ, спрацьовує нуклеазний активний центр, помилковий нуклеотид відщеплюється, і ДНК-полімераза здійснює нову спробу подовження ланцюга. У результаті такої осциляції полі- мерази з перемиканням активності між двома центрами рівень помилок знижується до ~10-8. Остаточна частота помилок становить ~10-10 за рахунок активності систем репарації (див. нижче), які спрацьовують під час і відразу після реплікації.
Дві ДНК-полімерази, що працюють у реплікативній вилці, об'єднані в складний мультибілковий комплекс - реплісому, компонентами якої є також інші важливі структурні та функціональні модулі: ДНК-геліказа - АТР-залежна молекулярна машина, що руйнує подвійну спіраль попереду від реплікативної вилки; праймаза, яка забезпечує синтез праймера - короткої ділянки РНК на початку кожного фрагмента Оказакі, після чого праймер подовжується ДНК-полімеразою (сама ДНК-полімераза не здатна ініціювати синтез нуклеїнової кислоти, а може тільки продовжувати синтез праймера); компоненти, що сприяють утриманню ДНК-полімераз у реплікативній вилці тощо.
РНК-праймер на початку кожного фрагмента Оказакі має бути замінений на відповідну послідовність ДНК. Ця робота виконується за рахунок 5'-екзонуклеазної активності певних ферментів, після чого ДНК-полімераза заповнює прогалину між сусідніми фрагментами Оказакі. У результаті між двома фрагментами Оказакі залишається одноланцюговий розрив, який зшивається ще одним важливим ферментом - ДНК-лігазою.
У клітині Escherichia coli працюють ДНК-полімерази трьох типів (позначаються римськими цифрами). Дві з них (І та ІІІ) належать до класу полімераз високої точності синтезу, ДНК-полімераза ІІ - поліме- раза низької точності, яка використовується в певних репараційних процесах. Основна реплікативна полімераза - ДНК-полімераза ІІІ. ДНК-полімераза І (або полімераза Корнберга), на відміну від інших ДНК-полімераз, має також додаткову 5'-екзонуклеазну активність - саме ця полімераза й використовується при з'єднанні фрагментів
Оказакі під час реплікації (видаляє праймер і заповнює прогалину), а також при репараційних процесах синтезу ДНК.
П'ять типів еукаріотичних ДНК-полімераз високої точності прийнято позначати грецькими літерами. Основними ДНК-синтезуючими (під час реплікації та репарації) є ДНК-полімерази б і є. Вони ж заповнюють прогалину між фрагментами Оказакі, що утворюється після видалення праймера певною нуклеазою. Полімераза а використовується як праймаза при ініціації синтезу лідируючого ланцюга й кожного фрагмента Оказакі (синтезує РНК-праймер і трохи подовжує його як ДНК). Полімераза в використовується при ексцизійній репарації основ Полімераза γ - реплікативна ДНК-полімераза мітохондрій.
В еукаріотичних клітинах працює ще досить велика кількість ДНК-полімераз низької точності (ζ, η, ι, κ), функція яких полягає в забезпеченні синтезу ДНК у випадку пошкодження матриці.
Характерною особливістю еукаріотичної системи реплікації є те, що подвоюється не циркулярна, як у прокаріотів, а лінійна молекула ДНК - така, що має два кінці. Унаслідок цієї простої обставини на З'-кінцях матричних ланцюгів ДНК залишаються одноланцюгові хвости (рис. 1.18): два РНК-праймери на 5'-кінцях ланцюгів, що синтезовані, видаляються, а прогалина не може бути заповненою, оскільки немає 3'-кінця, який міг би бути використаним як праймер. Одноланцюгові хвости піддаються швидкій нуклеазній деградації, і після кожної реплікації ДНК повинна вкоротитися.

Рис. 1.18. Дві дочірні лінійні молекули ДНК після реплікації
Кінцеві ділянки молекул ДНК, що містяться у клітинному ядрі, - теломери - складаються з невеликих елементів послідовності (шість, рідше вісім нуклеотидів), які тандемно повторюються - теломерних повторів. У хребетних і більшості вищих рослин теломерний повтор є однаковим - TTAGGG. Подовження теломер після реплікації здійснюється за допомогою спеціального ферменту - теломерази, яка є РНК-залежною ДНК-полімеразою. РНК-матриця входить до складу самого ферменту й містить ділянку, комплементарну теломерному повтору. Використовуючи її як матрицю і 3'-кінець як праймер, теломераза покроково добудовує до 3'-кінця кілька копій теломеразного повтору. Теломераза є активною у проліферуючих недиференційованих клітинах і в злоякісно трансформованих клітинах і неактивною - у диференційованих соматичних клітинах вищих еукаріотів. Певне критичне скорочення теломер, яке відбувається у таких клітинах після кількох десятків клітинних поділів, є одним із ключових механізмів активації програми їхньої загибелі (див. розділ 6).
Подовжений теломеразою одноланцюговий хвіст використовується як матриця для синтезу іншого ланцюга за звичайним реплікативним механізмом. Після видалення РНК-праймера на кінцях подовженої хромосоми (у складі G-ланцюгів, збагачених на гуанін) залишаються одноланцюгові 3'-вирости (як на рис. 1.18). За рахунок взаємодії зі специфічними білками одноланцюговий виріст "втягується" у дволанцюгову ДНК, порушуючи при цьому водневі зв'язки дуплекса: утворюється закрита форма теломери, що називається t-петлею (рис. 1.19).
Основна функція t-петлі полягає в захисті кінців лінійної молекули ДНК від деградації екзонуклеазами та в тому, щоб зробити кінець хромосоми непомітним для репараційних систем: відкрита форма те- ломери буде сприйматися репараційними системами як розрив, що може призвести до об'єднання кінців двох різних хромосом.

Рис. 1.19. Схема організації t-петлі в теломерній ділянці хромосоми