ГЕНЕТИКА - Підручник - А.В. Сиволоб - 2008
РОЗДІЛ 6. Генетика багатоклітинних еукаріотів
ГЕНЕТИКА ІНДИВІДУАЛЬНОГО РОЗВИТКУ
Розвиток Caenorhabditis elegans
Розвиток нематоди C. elegans від зиготи до дорослої особини включає чотири личинкові стадії, які розділяються періодами линяння (L1, L2, L3 та L4). Протягом кожної личинкової стадії формуються окремі структури й диференціюються тканини тіла дорослої особини. Після останнього линяння утворюється доросла особина, яка має 959 соматичних клітин у гермафродитних форм або 1031 у самців. Доля кожної окремої клітини в ході розвитку C. elegans є відомою, тобто кожна клітина має свій описаний "родовід", який починається від першого дробленням зиготи й завершується утворенням функціональної диференційованої клітини.
У результаті першого поділу зиготи в передньо-задній площині зародку утворюються дві неоднакові за розміром клітини - АВ (велика, нащадки якої будуть формувати клітинні лінії передньої частини зародка) і Р1 (мала, її нащадки формуватимуть клітинні лінії задньої частини зародка). При цьому лише клітина Р1 має у своїй цитоплазмі Р-гранули - специфічні цитоплазматичні детермінанти, необхідні для утворення попередників статевих клітин. Подальший поділ клітини Р1 у передньо-задній площині дає дві клітини EMS(нащадки цієї клітини - MS та E - будуть визначати вентральну вісь зародка) і Р2. За рахунок нерівномірної сегрегації Р-гранул у цитоплазмі клітини Р1, ці гранули потрапляють тільки до клітини Р2 (усі клітини, які будуть мати Р-гранули після наступних поділів, і, відповідно, будуть детермінувати розвиток статевих клітин, позначаються літерою Р). При наступному поділі з Р2 утворяться дві клітини - С і Р3, поділ останньої зумовить появу ще двох клітин D і Р4 (рис. 6.20).
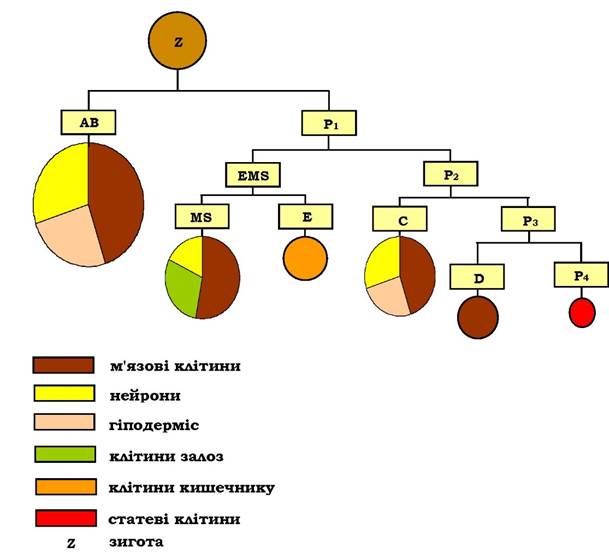
Рис. 6.20. Схема утворення клітин-попередників шести основних клітинних ліній C. Elegans
Отже, у перших поділах зиготи визначаються дорсовентральна та передньо-задня осі зародку, а шість клітин (АВ, MS, E, С, D і Р4), що утворюються при цьому, є попередниками шести основних клітинних
ліній личинки нематоди, які сформують основні тканини та органи дорослої особини. Слід відзначити, що первинна детермінація попередників статевих клітин відбувається вже в першому поділі зиготи.
Шляхи детермінації та диференціації клітин інваріантні в усіх особин даного виду. Вони регулюються продуктами певних генів, які умовно можна розділити на три групи, що контролюють: а) детермінацію клітин; б) час детермінації клітин та їхній поділ; в) селективну загибель певних клітин у ході розвитку. Крім того, у диференціації клітин C. elegans значну роль відіграють міжклітинні взаємодії.
Найдетальніше генетичний контроль розвитку C. elegans досліджено на прикладі формування вульви - жіночої статевої системи в гермафродитів. Вульва формується із семи клітин, одна з яких стає якірною клітиною, інші - клітинами-попередниками статевих шляхів, матки тощо. Вибір того, яка з клітин стане якірною, визначається щонайменше п'ятьма генами. При цьому важливою є не тільки сама наявність п'яти ключових генів, а також час їхньої експресії. Так, у мутантів за одним із цих генів - lin-14 - формування вульви відбувається на одну личинкову стадію раніше, ніж у нормі, що спричинює аномальний розвиток. Мутації, які змінюють час диференціації окремих клітин чи органів, називають гетерохронічними.
Після розподілу клітин на якірну та клітини-попередники, останні починають детермінуватися, утворюючи клітини трьох типів - первинні (1°), вторинні (2°) і третинні (3°). Цей етап перебуває під контролем щонайменше 15 генів, а сам процес детермінації має каскадний характер: якірна клітина стимулює розвиток первинної клітини, яка, у свою чергу, - вторинної і т. д. Особливе значення має місце взаєморозташування клітин: наприклад, при загибелі клітини 2°, найближча до неї клітина 3° стає вторинною. Тип регуляції детермінації, на який впливають оточуючі клітини, називається позиційним (рис. 6.21). Пізніше первинні та вторинні клітини- попередники будуть диференціюватися в тканини вульви, а третинні - у гіподерміс, який оточує вульву.
У процесі розвитку C. elegans шляхом послідовних мітотичних поділів утворюється 1090 соматичних клітин, дорослі особини - гермафродити при цьому мають 959 клітин, тобто 131 клітина елімінується протягом личинкових стадій. Універсальний для всіх еукаріотів механізм клітинної загибелі, при якому відбуваються точно запрограмовані біохімічні й морфологічні зміни окремих клітин, називається апоптозом.
Апоптоз контролюється великою кількістю внутрішньоклітинних і міжклітинних сигналів. Визначальну роль у розвитку запрограмованої клітинної загибелі відіграють специфічні протеази - каспази (розрізають молекули поліпептидів біля залишків аспарагінової кислоти). У клітині каспази знаходяться в неактивній формі - прокаспази - і не функціонують. Активація однієї з цих протеаз викликає каскадну активацію інших, що, у свою чергу, зумовлює активацію нуклеаз, деградацію ДНК і білків, які підтримують клітинні мембрани та цитоскелет, і, відповідно, загибель клітини. Сигналом для активації прокаспаз у процесі розвитку може бути нестача поживних речовин, факторів росту, міжклітинні сигнали різної природи тощо.
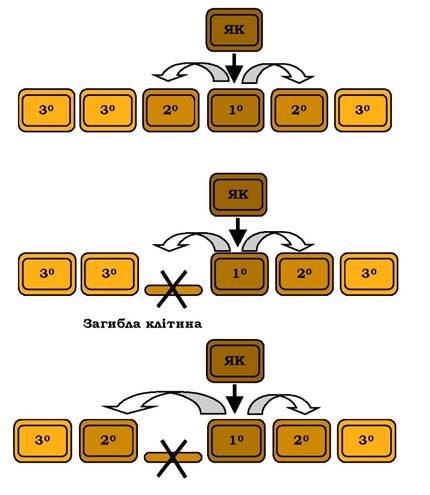
Рис. 6.21. Позиційний контроль розвитку клітин-попередників вульви у C. elegans.
Стрілками вказані позиційні сигнали детермінації розвитку. ЯК - якірна клітина
У C. elegans є три ключові гени, які контролюють селективну загибель клітин: ced 3, ced 4 і ced 9. Продукти генів ced 3 і ced 4 є індукторами апоптозу, мутації за цими генами зумовлюють повну відсутність елімінації 131 клітини. Продукт гена ced 9, навпаки, є інгібітором апоптозу: інактивація цього гена приводить до елімінації клітин, яка в нормі не відбувається. Білок Ced-3 є прокаспазою, що міститься у клітині в неактивній формі. Каспаза Ced-3 активується продуктом гена Ced-4, котрий індукує саморозрізання прокаспази й перетворення її в активу форму. У клітинах, в яких апоптоз не повинен відбуватися, білок Ced-4 блокований білком Ced-9. Апоптичний сигнал приводить до дисоціації Ced-9 від Ced-4 і запуску каспазного каскаду.

Рис. 6.22. Сигнальниі шляхи розвитку апоптозу у C. elegans і ссавців
Гени, які контролюють апоптоз, досить консервативні. У хребетних знайдені білки, гомологічні таким C. elegans. Зокрема, білок bcl-2, як і його гомолог Ced-9, блокує апоптоз у клітинах ссавців. Обидва білки мають трансмембранний домен і розміщені на зовнішній поверхні мітохондрій, ядра та мембранах ендоплазматичного ретикулуму, що дозволяє їм виступати сенсорами апоптичного сигналу. Білок Apaf-1 (гомолог Ced-4 у ссавців) активує перетворення прокаспази 9 в активну форму і таким чином запускає активаційний каскад протеаз. Білки Ced-4 та Apaf-1 є адапторами апоптичного сигналу, а білками- ефекторами виступають різноманітні каспази (рис. 6.22).