ТЕОРЕТИЧНІ ОСНОВИ РИБНИЦТВА - І.М. Шерман - 2011
3. ТЕОРЕТИЧНІ ОСНОВИ ВЕДЕННЯ РИБНИЧИХ ТЕХНОЛОГІЧНИХ ПРОЦЕСІВ
3.4. Особливості оптимізації зимівлі риб
Риби є пойкілотермними тваринами і це відповідним чином висвітлене вище, що суттєво зменшує необхідність поглиблюватися в це явище, яке характерне не тільки для риб, воно типове для всієї фауни, в якої відсутня система терморегуляції, і температура особин фактично повністю залежить від температури оточуючого середовища.
Помірні і високі широти демонструють достатньо чітку зміну пор року, що закономірно супроводжується зміною температур повітря і, як наслідок, термічного режиму відповідних акваторій. Мешканці помірних і високих широт в процесі тривалого філогенезу напрацювали відповідні механізми адаптації до таких явищ і це стало нормою для виду в ареалі. Сформувалися відповідні іхтіоценози природних акваторій у складі яких, відповідно до термічного режиму визначалися стенобіонти і еврибіонти. При цьому мешканці морів і океанів, завдячуючи великим об’ємам води, представлені переважно стенобіонтами, а мешканці озерно - річкових систем, з малими об’ємами води, де зміни термічного режиму можуть бути достатньо стрімкими, еврибіонтами.
Людство в процесі доместикації риб досягло певних позитивних результатів і сьогодні має місце ефективне культивування морських і прісноводних видів риб, у складі яких є еврибіонти і стенобіонти.
Виходячи з викладеного, зрозуміло, що особливості оптимізації зимівлі риб тісно пов’язані з термічним режимом і фізіолого - біохімічними реакціями організму на це явище, значення якого переоцінити важко.
У сучасному рибництві зимівля є однією з важких ланок технологічного процесу в тепловодному і холодноводному рибництві. Сучасні рибничі господарства, залежно від культи об’єктів культивування, представлені відповідно тепловодними і холодноводними підприємствами, що формує відповідну специфіку технологічних процесів у часі та просторі. Холодноводні рибничі господарства практично не пов’язані з паузами у годівлі і це обумовлює практично безперервну годівлю протягом року, що робить достатньо проблематичним розглядати зимівлю, як конкретний технологічний процес, відокремлений від безпосереднього виробництва рибної продукції, вирощування ремонту і утримання плідників. Для холоднолюбивих видів риб, а це переважно хижаки, які живляться протягом року, залежно від динаміки термічного режиму раціон корегується. При цьому холодолюбиві види риб є оксифілами, які на тлі відповідного термічного режиму постійно вимагають високого вмісту розчиненого у воді кисню у поєднанні з інтенсивним водообміном з метою винесення з середовища продуктів метаболізму.
На відміну від холодноводного рибничого господарства в тепловодному рибничому господарстві культивують переважно мирних видів риб. Серед них переважають еврибіонти по відношенню до термічного режиму у весняно - літній період у переднерестовий період, але у осінньо - зимовий період вони практично припиняють живлення, зменшують рухливість і фактично до мінімуму скорочують витрати енергії, основою якої в цей час є енергетичні складові, накопичені організмом протягом попередньої вегетації, тобто нагульного періоду.
Сучасне ставове тепловодне рибне господарство ґрунтується на полікультурі коропа і рослиноїдних риб, інтенсивність живлення яких, темп росту за інших однакових умов щонайтісніше пов'язані з температурою води. Зі зниженням температури води до 8 - 9 °С ці риби припиняють інтенсивне живлення, різко знижують рухову активність, концентруються у поглибленнях дна водойми. У цей період уповільнюється обмін речовин, а їх енергетичні потреби задовольняються за рахунок раніше накопичених жирових запасів, що супроводжується втратою маси і зниженням коефіцієнту вгодованості. Поряд з цим для коропа, рівно як і для інших видів риб для дихання потрібен кисень, кількість якого зменшується у міру ослаблення фотосинтетичної діяльності водяних рослин, а з утворенням льоду різко зменшується дифузія кисню з повітря, що об'єктивно негативно впливає на рибу.
Виходячи з викладеного, ще восени, потрібно організувати зимове утримання риби - зимівлю - один зі складних технологічних процесів у ставовому рибництві, коли на одиниці площі зимувальних ставів створюються досить високі концентрації як цьоголітків, так і ремонтного і маточного поголів'я, а за трирічного обороту - і дволітків.
Від успішності зимівлі часто залежить результат усієї господарської діяльності рибницького підприємства, оскільки понаднормативний відхід риби у період зимівлі створює різкий дефіцит рибопосадкового матеріалу. Практика засвідчує, що відходи цьоголітків за зиму часто становлять близько половини усієї посадженої на зимівлю риби, а в деяких господарствах за зиму іноді гине практично весь рибопосадковий матеріал.
Залежно від наявних у господарстві можливостей зимівлю проводять у спеціальних зимувальних ставах, пристосованих для зимівлі, вирощувальних і нагульних ставах, у саджалках і басейнах, що обумовлено прийнятою технологією. Головна і загальна вимога, яку ставлять до зимувальних ставів, — створення відповідних умов для зимуючої риби, а це перш за все достатня глибина, проточність води, задовільні фізико-хімічні параметри середовища.
Результати зимівлі залежать не тільки від абіотичних параметрів середовища, а і від фізіологічного стану риби, хімічного складу тіла, розмірів, маси, коефіцієнта вгодованості. Протягом зими маса цьоголітків зменшується в середньому на 10 - 12 %, рідше — на 16 - 17, довжина, висота, ширина тіла — на 7,5 %. Дволітні риби втрачають значно менше — 6, рідше — 7 - 10 % маси тіла, а довжина, ширина і висота їх тіла зменшуються на 3,5 — 4,0 %; втрати жиру у цьоголітків становлять майже 50 % (836 — 1254 Дж), втрати білка — 17—30 (418-627 Дж), а загальні енергетичні витрати дорівнюють 32 - 40 %.
Отже, зрозуміло, що цьоголітки мають бути добре підготовленими до тривалого обміну речовин в умовах фактичного голодування, що
досягається достатнім накопиченням в організмі резервних речовин протягом вегетаційного періоду, серед яких основну роль відіграють жирові залози. У цьоголітків, вирощених на природних кормах, вміст жиру має становити 1,8 — 3,0 %, на штучних — не менше 4, білка — відповідно 12 і 11 %.
Проте у практиці ставового рибництва більш широко використовуються такі критерії оцінки зимостійкості як: розмір, маса, вгодованість. Ці показники взаємозв'язані між собою і певною мірою характеризують загальний стан риби, що дозволяє прогнозувати результати зимівлі.
В умовах суворих зим, які типові для зони Полісся, найбільш стійкішими є цьоголітки лускатих коропів. Це певною мірою пояснюють розміщенням жиру під лускою, який захищає особин від дії низьких температур у період зимівлі. Враховуючи наявність проблеми зимостійкості для її підвищення, виконано спеціальні роботи, орієнтовані на створення гібридних форм коропа й амурського сазана, помісних гібридів між українським, рамчастим і ропшинським коропом. У результаті отримано зимостійкі породи коропа, а саме: ропшинського, українського, нивківського, сарбоянського. У промислових господарствах, де важко створити умови, наближені до оптимальних для зимівлі, доцільно використовувати гібриди першого покоління, які мають високу виживаність при вирощуванні та зимівлі. На виживаність молоді у період зимівлі суттєвий вплив має якість плідників, їх вік і походження. Неприпустимі близькоспоріднені парування, використання старих і молодих самців та самиць, перевагу доцільно віддавати середньовіковим групам в процесі штучного відтворення. Результати зимівлі, як наголошено вище, залежать від підготовки зимувальних ставів, створення відповідних умов середовища для зимуючої риби. При цьому доцільно керуватися відповідними критеріями - глибина зимувальних ставів має бути такою, щоб непромерзаючий шар води становив 1,2, у північних районах - 1,5 - 1,75м. Оптимальні розміри ставів - 0,2 - 1,0 га за співвідношення довжини і ширини 2:1. Ложе зимувальних ставів має бути сплановане з таким розрахунком, щоб нахил був у бік водоспуску. По дну ставу влаштовують рибозбірні канали, а біля водоспуску - рибозбірні ями (приймальники) або рибовловлювачі. Це дає змогу прискорити процес облову зимувальних ставів, зменшити травмування риби, суттєво скоротити проведення її антипаразитарної обробки. Водоспуски зимувальних ставів обладнують двома рядами спеціальних щитів, або іншими словами шандор. Водопостачання їх може здійснюватись із різних водних джерел: головного ставу, джерел, річки, артезіанської свердловини. Вода, що надходить у став, має містити розчиненого заліза не більш як 0,8 мг/л, мати загальну твердість 5-8 мг-екв/л, окиснюваність — не вище як 10 мг О2/л, рН — 7,2 — 8,6, мінімальний вміст сульфатів і хлоридів. Наявність у воді розчинених шкідливих газів метану і сірководню неприпустима.
Вміст кисню у водному середовищі повинен бути у межах 8 - 9 мг/л. Надходження у зимувальні стави стічних вод чи шкідливих речовин (фенолів, гербіцидів, нафти) не допускається.
Зимувальну систему ставів обладнують аераційними установками різних конструкцій, які за потреби дають змогу збільшити вміст розчиненого кисню у воді. Якщо реакція середовища у джерелі водопостачання кисла, на водоподаючому каналі або лотку встановлюють вапняні фільтри. За подачі води на відповідних спорудах обов'язково влаштовують рибозагороджувальні решітки, фільтри, сміттєвловлювачі для захисту ставів від потрапляння риби. Лотки, водоподавальні канали, труби утеплюють, вкриваючи їх дошками, а зверху - землею з гноєм.
Вживають також заходів щодо захисту зимувальних ставів від надходження талих вод шляхом відведення їх обвідним каналом. Особливу увагу приділяють своєчасному ремонту гідротехнічних споруд, які забезпечують безперебійну водоподачу взимку у достатній кількості та доброї якості.
Зимувальні стави влітку утримують сухими, що сприяє їх знезаражуванню і мінералізації органічних речовин. Після весняного розвантажування канави по ложу ставу, водоподаючі та водоскидні канали розчищають від мулу і рослинності, а в літній період зимувальні стави 2 - 3 рази обкошують. Наприкінці вересня - в жовтні стави дезинфікують з розрахунку 2,0 - 2,5 т негашеного вапна на 1 га, промивають і заливають водою за 10 - 15 днів до посадки риби.
Час пересаджування цьоголітків на зимівлю визначають за температурними умовами (часом припинення живлення і росту), але обов’язково до настання морозів, які можуть призвести до підвищеного травмування й обмороження риби. Проте, якщо температура води ще досить висока (понад 6-8 °С) й у вирощувальних ставах є природний корм, пересаджування у зимувальні стави може призвести до подовження часу голодування цьоголітків і підвищення відходу їх за зиму. Водночас не можна затягувати час пересаджування риби до заморозків, оскільки за мінусових температур можливе обмороження зябер, що здатне спричинити загибель риби. Якщо у господарстві є кілька зимувальних ставів, то рибу з окремих вирощувальних ставів краще пересаджувати у певний зимувальний, але хронічний дефіцит зимувальних площ робить це побажання достатньо проблематичним.
Під час пересаджування не можна перевантажувати чани, живорибні автомобілі, ноші, оскільки цьоголітки можуть отримати травми. Для розвантаження спеціальних ємкостей застосовують брезентові рукави, лотоки, по яких цьоголітків разом з водою зливають безпосередньо у став. Щільність посадки цьоголітків у зимувальні стави залежить від певної грунтово - кліматичної зони. За більшої тривалості зимівлі щільність посадки менша і навпаки, що забезпечить успішну зимівлю при урахуванні цього положення, табл. 3.16.
Таблиця 3.16. Нормативи утримання риби в спеціальних зимувальних ставах
Показник |
Норматив |
Щільність посадки цьоголітків у зимувальні стави |
|
за роздільного утримання, тис. екз/га |
|
коропа |
500 - 800 |
рослиноїдних риб |
450 - 550 |
Вихід однорічок із зимувальних ставів від посадки |
|
цьоголітків, % |
|
коропа |
70-85 |
рослиноїдних риб |
70-85 |
Вихід однорічок із пристосованих водойм від |
|
посадки цьоголітків, % |
|
коропа |
60-75 |
рослиноїдних риб |
60-75 |
Втрата маси, % |
10-12 |
Щільність посадки дволітків у зимувальні стави, |
|
тис. екз/га |
|
коропа |
110-120 |
рослиноїдних риб |
130-160 |
Вихід дворічок із зимувальних ставів, % |
|
коропа |
90 |
рослиноїдних риб |
80 |
Втрата маси, % |
До 10 |
У зимовий період здійснюють регулярний контроль за станом риби і якості води зимувальних ставів. Не можна допускати, щоб зимувальні стави були занесені глибоким снігом, що передбачає своєчасне зметання снігу для забезпечення прозорості льоду, постійно стежити за станом водоподаючих труб, лотків, водоспусків, збиваючи з них лід.
У зимувальних ставах встановлюється 15 - 30-добовий водообмін. Це означає, що за такий період має відбутись повна заміна води. Посилювати проточність без вагомих причин недоцільно, оскільки це призводить до підвищення рухливості і, як наслідок, до виснаження риби. Потрібно стежити за рівнем води, не допускати зниження чи підвищення її горизонту, щоб не спричинити руйнування криги або пошкодження водоспуску і переливання води. Для контролю за станом риби роблять кілька ополонок розміром 1 х 2,0; 1,5 х 2,5м, іноді у вигляді літери «Г», їх потрібно щоденно очищати від криги і зверху закривати щитами, очеретяними матами з метою виключення замерзання.
Кількість води, потрібної для подачі кисню у зимувальні стави, визначають за формулою
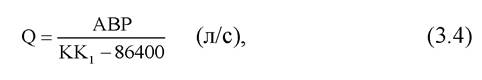
де А - кількість цьоголітків, посаджених у став, екз.; В - середня маса цьоголітків, г; Р - витрата кисню на 1 кг маси цьоголітків за одну добу, мл; К, Кі - вміст кисню у воді на припливі й витоку, мг/л.
За температури води до 3°С на 1 кг цьоголітків витрачається 250 мл кисню, за 15 - 16 °С - до 5000 мл, що підтверджує потребу постійно контролювати стан хімічного і термічного режимів у зимувальних ставах.
Температура води є дуже важливим показником, оскільки саме її відповідність видоспецифічним особливостям риб у зимовий період є найважливішою умовою благополучної зимівлі. Спеціальними дослідженнями встановлено, що оптимальна температура дорівнює 1-2°С. Зниження її до 0,1 - 0,2°С спричинює захворювання риб від переохолодження. За температури понад 4°С риба починає рухатись, витрачає енергетичні запаси, виснажується, стає чутливішою до несприятливих чинників навколишнього середовища і різних хвороб, що є причиною багатьох проблем, які виникають за невідповідності зимівлі вимогам виду.
Г азовий режим впливає на підсумки зимівлі не меншою мірою, ніж температурний. У зв'язку з цим не менш, як три рази за зимівлю здійснюють повний гідрохімічний аналіз води. Після льодоставу аналізи води на вміст розчиненого у ній кисню доцільно проводити кожні 2 - 5 днів, а в разі зниження вмісту кисню - щоденно. Час відбирання проб не має істотного значення в зимувальний період, оскільки лід суттєво перешкоджає проникненню світла у став за умови втрати ним прозорості, що обмежує фотосинтетичну діяльність водоростей. Проби води в зимувальних ставах відбирають у двох точках: на водоподачі (з глибини 0,5 - 0,7м, на відстані 2 - 3м від спадного струменя води) і біля водоспуску (ближче до дна). За погіршення газового режиму чи наявності різниці у вмісті кисню на водоподачі і водоскиду (2,0 - 3,0 мг/л) проби відбирають і в центрі ставу. Нормальним вважають вміст розчиненого у воді кисню на витоку - 4-5мг/л, вуглекислого газу - до 20мг/л при рН 7 - 8 і окиснюваності 10 - 25мг Ог/л.
Зі зниженням вмісту кисню до 3 мг/л і менше воду треба аерувати, перекачувати з ополонки в ополонку різними механізмами, машинами, влаштовувати на водоподавальних каналах чи лотках східчасті перепади, столики-аератори під водоподавальною спорудою. Доброї аерації можна досягнути за допомогою пересувних і стаціонарних повітряних компресорних установок.
За нестачі у воді кисню, захворюваннях і надмірному виснаженні цьоголітків починається їх рух у ставу, риба підпливає до контрольних ополонок. Для з'ясування причини руху цьоголітків виловлюють і досліджують, визначають коефіцієнт вгодованості, аналізують фізико- хімічні умови. Спостереження за зимуючою рибою і ставами записують у спеціальний журнал. Іноді для контролю за станом риби у зимувальному ставу встановлюють саджалки (розміром 1 х 0,5 х 0,5 м), куди саджають 100 цьоголітків. Підіймаючи саджалки 2-3 рази за зиму, рибу оглядають, вимірюють, зважують, визначають коефіцієнт вгодованості, враховуючи, що її загибель може настати за зменшення коефіцієнта вгодованості нижче 2,0, вмісту жиру - 0,5, білка - 7% для коропа.
Зимівля риби старших вікових груп (ремонтного і маточного поголів'я) здійснюється у зимово-ремонтних і зимово-маточних ставах (табл. 3.17).
За нормальних умов старші вікові групи перезимовують майже без відходу. Зимівлю плідників рослиноїдних риб слід проводити окремо від коропа, хоча вимоги до умов зимівлі в них однакові.
Таблиця 3.17. Результати зимівлі піддослідних риб
Номер садка |
Вид риб |
Посаджено цьоголіток |
Виловлено річняків |
Вихід, % |
||||
тис.екз/га |
середня маса, г |
т/га |
||||||
тис.екз/га |
т/га |
|||||||
1 |
Білий товстолобик |
810 |
57,7 |
46,74 |
550 |
28,27 |
67,9 |
|
2 |
Строкатий товстолобик |
810 |
77,8 |
62,92 |
750 |
43,98 |
92,6 |
|
3 |
Білий амур |
810 |
14,1 |
11,42 |
740 |
8,79 |
91,3 |
|
4 |
Короп |
810 |
92,6 |
75,01 |
650 |
69,61 |
80,2 |
|
Щільність посадки ремонтного і маточного поголів'я у зимувальні стави визначають з урахуванням загальної їх маси. Згідно з чинними нормативами щільність посадки коливається від 10 до 20 т/га. Під час зимівлі дволітків, ремонтного молодняку і плідників здійснюють ретельний догляд за рибою і контроль за умовами її життя. Результати спостережень за ходом зимівлі (дані гідрохімічного, термічного і гідрологічного режимів, характеристика поведінки риби, можлива загибель та її причини) записують у журнал.
Розвантажують (обловлюють) зимувальні стави залежно від кліматичних і погодних умов з березня до середини травня. Після скресання криги на зимувальних ставах за температури 3 - 5 °С починають пересаджування однорічок у нагульні стави, різновікових ремонтних груп - у літньо-ремонтні стави, а плідників - у переднерестові або пристосовані для цього інші категорії ставів. Тривале затримання з пересаджуванням риби може призвести до загибелі значної кількості рибопосадкового матеріалу. Якщо з якихось причин пересаджування неможливе, організовують підгодівлю риби безпосередньо у зимувальних ставах. Це дає змогу підтримувати молодь у задовільному фізіологічному стані і, крім того, до зариблення нагульних ставів провести безпосередньо у зимувальних ставах курс профілактичної або лікувальної годівлі однорічок, ремонтного матеріалу різних вікових груп.
Під час облову зимувальних ставів воду спускають, рибу виловлюють спочатку неводами по приспущеній воді, а потім у рибозбірній ямі чи рибовловлювачі. Рибу, яка перезимувала, перелічують, встановлюють вихід відносно кількості особин, посаджених у став восени; визначають її середню масу і вгодованість, епізоотичний стан (табл. 3.18). Кожну партію обробляють в антипаразитарних ваннах, у транспортних резервуарах або безпосередньо за наявності відповідних умов у ставах. Вихід однорічок після зимівлі має становити 70 - 80, старших вікових груп - 95 - 100 %.
Поряд із класичним методом зимівлі у спеціалізованих ставах використовують вирощувальні стави, особливо у південних районах. Щільність посадки риби на зимівлю розраховують на глибоководну частину ставу, на ділянку, де шар непромерзаючої води досягає 1,0 - 1,2 м. Вихід однорічок відносно кількості посаджених цьоголітків становить 60 - 75%, втрати за масою невеликі, оскільки риба забезпечена кормами у пізньоосінній і ранньовесняний періоди, а також під час тривалих зимових потеплінь, що часто має місце у господарствах півдня України.
Значний інтерес представляє вплив зимівлі на гематологічні показники піддослідних риб табл. 3.19. Показники крові зимуючих риб протягом досліду характеризувалися певними коливаннями.
Для трьох вивчених видів риб далекосхідного комплексу характерне збільшення кольорового показника крові з тенденцією зменшення вмісту гемоглобіну в крові (статистично вірогідно тільки для білого товстолобика) і зниження кількості еритроцитів (статистично вірогідно для білого і строкатого товстолобика). Зміна вмісту загального білка сироватки крові у зв'язку із зимівлею статистично недостовірна. Гематологічні показники коропа мали незначні коливання і були статистично недостовірними.
Зменшення вмісту в крові формених елементів і гемоглобіну пояснюється зниженням загального рівня обміну речовин риб у зимових умовах.
Таблиця 3.18. Вплив зимівлі на лінійні розміри (L, l), масу тіла (р) і вгодованість коропа (Ку) рослиноїдних риб (M±m)
Вид риб |
Період |
L, cм |
l, см |
р, г |
КВ |
||
Білий товстолобик |
До зимівлі |
18,1 ± 0,1 |
16,0 ±0,1 |
57,7± 1,1 |
(Р > 0,999) |
1,70 ± 0,01 |
(Р > 0,999) |
Середина зимівлі |
17.9 ±0,5 |
14,8 ±0,4 |
56,6 ±3,6 |
1,55 ± 0,01 |
|||
Після зимівлі |
19,1 ± 0,7 |
14,9 ±0,08 |
51,4 ± 0,7 |
1,52 ± 0,06 |
|||
Строкатий товстолобик |
До зимівлі |
18,0 ±0,3 |
15,2±0,3 |
77,8 ± 4,0 |
(Р > 0,999) |
1,96 ± 0,01 |
(Р > 0,999) |
Середина зимівлі |
17,6 ±1,4 |
15,0 ±0,4 |
66,3 ± 6,01 |
1,96 ± 0,02 |
|||
Після зимівлі |
17,3 ±0,3 |
14,3 + 0,3 |
58,5 ± 3,7 |
1,91 ±0,03 |
|||
Білий амур |
До зимівлі |
10,6 ±0,1 |
8,7 ±0,1 |
14,1 ± 0,5 |
(Р>0,999) |
2,14 ± 0,005 |
(Р > 0,999) |
Середина зимівлі |
11,2±0,3 |
9,4 ±0,2 |
13,9 ±0,9 |
1,67±0,005 |
|||
Після зимівлі |
10,4 ±02 |
8,6 ±0,1 |
11,9 ±0,6 |
1,84 ± 0,05 |
|||
Короп |
До зимівлі |
17,6 ±0,4 |
14,6 ±0,3 |
92,6 ± 8,7 |
(Р>0,999) |
2,97 ±0,09 |
(Р > 0,999) |
Середина |
16,4 ±0,3 |
13,8 ±0,3 |
76,1 ±5,9 |
2,89 ± 0,04 |
|||
Після зимівлі |
18,9 ±0,3 |
15,7 ±0,3 |
107,1 ± 5,6 |
2,65 ± 0,01 |
|||
Таблиця 3.19. Вплив зимівлі на гематологічні показники коропа й рослиноїдних риб (М ± m)
Вид риб |
Період |
Загальний |
Еритроцити, млн |
Лейкоцити, |
Гемоглобін, |
Кольоровий |
||
Білий товстолобик |
До зимівлі |
4,92±0,75 |
1,58 ±0,08 |
46,(5 ± 2,35 |
11,6 ±0,3 |
1,95 ±0,08 |
||
Середина зимівлі |
2,24 ±0,08 |
35,4 ± 0,08 |
9,5 ±0,31 |
1,30 ±0,03 |
||||
Після зимівлі |
4,10±0,40 |
1,08±0,1 |
34,0 ±0,24 |
8,2 ± 0,2 |
2,34 ± 0,1 |
(Р> 0,99) |
||
Строкатий товстолобик |
До зимівлі |
7,69 ±0,46 |
2,34 ±0,10 |
35;4 ± 2,90 |
9,2 ±0,5 |
2,12 ± 0,07 |
||
Середина зимівлі |
2,19 ±0,23 |
24,6 ± 4,21 |
8,68 ± 0,36 |
1,21 ± 0,04 |
||||
Після зимівлі |
6,20+0,28 |
1,05 ± 0,04 |
(Р>0,99) |
34,0 ± 0,40 |
8,36 ± 0,46 |
2,39 ± 0,08 |
(Р> 0,95) |
|
До зимівлі |
5,62 ± 0,59 |
1,17 ± 0,25 |
42,6 ± 5,95 |
7,4 ±0,2 |
1,93 ± 0,15 |
|||
Білий амур |
Середина зимівлі |
1,60 ±0,08 |
41,8 ±1,83 |
7,33 ±0,21 |
1,31 ±0,06 |
|||
Після зимівлі |
4,9212,10 |
0,90 ±0,03 |
42,4 ±0,21 |
7,24 ±0,27 |
2,41 ± 0,05 |
|||
До зимівлі |
4,82 ±0,07 |
1,04 ± 0,22 |
(Р>0,99) |
28,8 ±2,06 |
7,00 ±0,001 |
2,10 ± 0,16 |
||
Короп |
Середина зимівлі |
0,59 ±0,12 |
29,0 ± 0,40 |
5,45 ±0,41 |
2,97 ± 0,20 |
|||
Після зимівлі |
5,45 ± 1,25 |
1,09 ±0,04 |
30,0 ±0,01 |
7,36 ± 0,56 |
2,10 ± 0,08 |
(Р> 0,99) |
||
Збільшення ж кольорового показника (вмісту гемоглобіну в одному еритроциті) обумовлено збільшенням дихальної функції крові, що компенсує зниження загального числа гемоглобіну й еритроцитів.
Проведені дослідження дозволяють стверджувати, що в ідентичних умовах спостерігається характерна для кожного виду специфічність зміни гематологічних показників.
Доцільно розглянути вплив зимівлі на біохімічний склад тіла риб (табл. 3.20, 3.21). Аналіз матеріалів свідчить про наявність видової специфічності мінливості. У коропа й білого амура склад тіла до зимівлі й після її майже не змінився (за винятком підвищення вмісту вологи в тілі білого амура), що характерно й статистично вірогідно для всіх трьох видів далекосхідного комплексу.
У тілі білого і строкатого товстолобика за зимівлю статистично вірогідно й значно зменшилася масова частка жиру (у перерахуванні на суху і сиру речовину). У тілі білого товстолобика зменшилася також масова частка протеїну (у перерахуванні на суху речовину).
Дослідження показали, що в умовах рибальського колгоспу «Придунайський рибалка" можлива зимівля великих цьоголіток коропа, і рослиноїдних риб за більш високих у порівнянні з нормативними щільностями посадки, а очевидний вплив зимівлі на екстер'єрні та інтер'єрні показники не є критичним для цих видів.
Ми усвідомлюємо той факт, що показники зимівлі в саджалках не можна інтерполювати на зимувальні ставки в цілому. Але проведені експериментальні роботи свідчать про доцільність розширення цих досліджень у перспективі і перевірки отриманих результатів у виробничих умовах.
Етологія зимуючої риби за наявності відповідних умов характеризується відносною нерухомістю, спокоєм, відсутністю суттєвих переміщень по зимувальному ставу. Підвищена рухливість, інтенсивне переміщення по зимувальному ставу, концентрація в місцях водоподаючих споруд свідчить про порушення умов зимівлі або стану здоров’я риби.
Таблиця 3.20. Біохімічний аналіз піддослідної риби, посадженої на зимівлю, % (М±m)
Показник |
Короп |
Білий товстолобик |
Строкатий товстолобик |
Білий амур |
Волога |
75,22±0,39 |
74,07±0,39 |
77,24±0,41 |
75,07±0,40 |
Суха речовина |
24,78±1,33 |
25,92±0,39 |
22,96±0,89 |
24,92±0,40 |
Протеїн |
||||
- на сиру речовину |
13,94±0,47 |
14,10±0,35 |
12,92±0,80 |
13,77±0,97 |
- на суху речовину |
26,48±4,57 |
28,82±1,12 |
23,08±0,61 |
28,70±1,84 |
Жир |
||||
- на сиру речовину |
6,78±1,54 |
7,57±0,43 |
5,22±0,06 |
7,20±0,61 |
- на суху речовину |
10,84±0,82 |
12,52±0,37 |
14,38±0,56 |
11,15±0,75 |
Зола |
||||
- на сиру речовину |
2,66±0,12 |
3,25±0,12 |
3,26±0,14 |
2,75±0,16 |
Таблиця 3.21. Біохімічний аналіз піддослідної риби, після зимівлі, % (М±m)
Показник |
Короп |
Білий товстолобик |
Строкатий товстолобик |
Білий амур |
|||
Волога |
74,70±0,28 |
77,32±0,21 |
(0,999) |
78,37±0,12 |
(0,95) |
77,32±0,37 |
(0,999) |
Суха речовина |
25,30±0,88 |
22,48±0,88 |
(0,99) |
21,62±0,40 |
22,67±0,86 |
||
Протеїн |
|||||||
- на сиру речовину |
13,42±0,14 |
13,50±0,50 |
12,87±0,16 |
13,30±0,10 |
|||
- на суху речовину |
32,02±2,40 |
18,26±1,59 |
19,65±1,16 |
24,52±1,72 |
|||
Жир |
|||||||
- на сиру речовину |
6,90±0,94 |
4,75±0,45 |
4,65±0,32 |
6,27±0,64 |
|||
- на суху речовину |
10,18±0,52 |
16,49±0,26 |
16,43±0,43 |
12,60±0,40 |
|||
Зола |
|||||||
- на сиру речовину |
2,57±0,10 |
3,70±0,13 |
(0,95) |
3,55±0,06 |
(0,90) |
2,80±0,10 |
|
Забезпечення розглянутих умов зимівлі риби, яке базується на відповідному термічному підґрунті, забезпечує досягнення нормальних виходів після зимівлі; гарантує нормальний фізіолого - біохімічний стан.
Сучасне рибництво до цього часу практично не має (за виключенням коропа і частково форелі) типових породних груп, порід за аналогією з тваринництвом. Процес доместикації в рибництві знаходиться в початковій стадії розвитку і, займаючись безпосереднім розведенням риб, рибовод переважно працює фактично з вихідними дикими формами конкретних видів риб. В зв'язку з цим розведення риб передбачає наявність достатнього" обсягу знань в галузі розмноження риб в природних умовах. Іншими сло - вами, необхідно вивчити філогенез виду в зв'язку з екологією розмноження. За Г.В.Нікольським, розмноження розглядається як ланка онтогенезу риби, що забезпечує у взаємозв'язку з іншими ланками відтворення популяції і збереження виду.
На відміну від теплокровних домашніх тварин і птахів, розведення риб досить специфічне, що обумовлено їх видовою різноманітністю, яка поєднується з виключно високою значимістю абіотичних, біотичних і антропогенних факторів. При цьому перша і друга група можуть впливати на риб безпосередньо, а третя група — антропічні, або антропогенні фактори, можуть діяти як безпосередньо, так і опосередковано через зміни кількісних і, як наслідок, якісних характеристик умов існування і розмноження риб. На жаль, об'єктивна реальність свідчить про те, що інтенсивність впливу антропогенних факторів зберігає тенденцію підвищення, що обумовлено нарощенням господарської діяльності людини в планетарному масштабі. Змінюються гідрологічний режим, фізико-хімічні показники континентальних і морських вод, що негативно впливає на видовий склад, чисельність і біомасу гідробіонтів, зокрема риби.
При цьому необхідно враховувати те, що матеріали, які характеризують екологічну пластичність певного виду риб, необхідно диференціювати в зв'язку з життєвими циклами. При даному підході стає очевидним, що процес розмноження риб характеризується досить вузькою екологічною валентністю і є виключно консервативним. Ця біологічна особливість знаходить своє фактичне відображення в динаміці чисельності популяцій риб, коли порушується екологія розмноження і, як наслідок, в окремі роки з'являються маловрожайні покоління. Так, випадіння певного виду риб із промислу відбувається протягом досить тривалого часу. Після чого іде ряд низьковрожайних поколінь молоді, причому кожне наступне покоління нижче урожайності попереднього. Якщо тенденція у часі зберігається, це приводить до повного припинення розмноження і зникнення виду не тільки із промислу, але і зі складу іхтіофауни даної водойми. При цьому водойма може зберегти своє рибогосподарське значення. Навіть за повного припинення розмноження основних промислових риб у водоймі, в результаті викликаного порушення екології розмноження, вона може бути використана в якості нагульної і експлуатуватися за принципом пасовищної аквакультури, що знайшло своє втілення в практиці світового рибництва. Проте, згадана практика збереження рибогосподарського значення водойм змогла стати реальністю тільки тому, що були розроблені теоретичні основи штучного риборозведення і вирощування молоді цінних видів риб до життєстійких стадій, які використовуються в якості інтродуцентів та вселенців для спрямованого формування іхтіофауни штучних і природних водойм.
Повертаючись до специфічних особливостей розмноження кожного виду, необхідно розглянути цей процес в адаптаційному плані. Специфічність або видоспецифічні особливості цього процесу не що інше, як пристосування до певних умов розмноження і розвитку молоді, що забезпечують циклічність поповнення і необхідну для збереження виду і підтримання чисельності популяцій в ареалі. У свою чергу чисельність поповнення і його якість залежить від кількості і якості нерестової популяції, умов ембріогенезу і раннього постембріогенезу.
Розмноження риб має специфічні особливості, характерні для водних тварин і обумовлені життям у воді. На відміну від теплокровних наземних тварин, що мешкають у воді, у абсолютної більшості риб запліднення яйцеклітин відбувається поза материнським організмом, в зовнішньому середовищі — воді. Ікра і молочко або ікринка і сперматозоїд до запліднення деякий час знаходяться у воді, поза особинами-плідниками, де і здійснюється проникнення сперматозоїда в яйцеклітину і відбувається утворення зиготи, що свідчить про наявність запліднення.
Розглядаючи розмноження риб і формуючи уявлення про предмет, необхідно окреслити коло питань, що визначають і складають цей виключно важливий процес у життєвому циклі всього живого, і риб зокрема. На думку А.П.Іванова, сюди потрібно віднести розвиток і формування статевих залоз, нерест, запліднення, ембріональний і постембріональний розвиток. Відносно останнього положення було б доцільно уточнити, що мова йде про ранній постембріогенез. Регуляція репродуктивної системи в риб, як і в інших класів хребетних тварин, регулюється системою ендокринних залоз, що знаходиться у свою чергу під контролем центральної нервової системи. Незважаючи на відому автономність взаємодії репродуктивної, ендокринної і нервової систем, на цей комплекс постійно впливають різноманітні фактори зовнішнього середовища, які також проявляють цілеспрямовану дію на розвиток різних ланок репродуктивної системи на всіх етапах онтогенезу. Регуляція функціонування репродуктивної системи має багаторівневий каскадний характер. Взаємодія різних рівнів регуляції в системі епіфіз - гіпофіз -гонада забезпечується системами прямого і зворотного зв'язку.
У гіпофізі - центральній ланці нейрогуморальної регуляції, основними гормонами, що здійснюють регуляцію розвитку гонад на всіх етапах онтогенезу, активно функціонують два, які розрізняються за фізико- хімічними властивостями і біологічній дією - ЛГ - і ФСГ-подібні гонадотропні гормони (ГТГ), що визначають за допомогою впливу на стероїдогенез і відповідний розвиток генеративних і соматичних елементів гонад. Відносний розвиток і функціональні взаємовідносини цих часток гонади між собою визначає весь подальший гонадогенез.
Основою фізіологічної регуляції розвитку і підтримки визначеної статі в риб різних систематичних груп є статева специфічність гіпофізарних ГТГ. Порушення або зміна цієї структурпо-функціональної специфічності ГТГ змінює певним чином характер стероїдогенезу в гонадах, приводячи до різноманітних порушень статевої функції, це явище переважно простежується у вигляді стерилізації гонад, інверсії статі, передчасного фізіологічного старіння репродуктивних систем.
Абсолютна більшість промислових видів риб, об'єктів культивування і розведення — різностатеві. Статеві залози самок — яєчники, в яких розвиваються зрілі статеві клітини — яйця (ікринки). Статеві залози самців — сім'яники, що продукують зрілі чоловічі статеві клітини — сперматозоїди. Сім'яники самців костистих риб мають власні сім'япроводи, які відкриваються в сечівник. При сперміації зрілі спермії звільняються із тканини сім'яника і виводяться через сім'япроводи в зовнішнє середовище.
Овуляція і сперміація — складні процеси, що мають нейрогуморальне управління. Знання механізму цих процесів має важливе значення за штучного розведення риб, і тому необхідно володіти достатнім обсягом інформації в даній галузі. Овуляція і сперміація характеризуються певними, послідовно замінюючими один одного цитологічними процесами. В плані фізіології ці процеси є не що інше, як функціональний метаморфоз тканини під дією гормонів. Фолікулярна тканина розпадається, видавлює із себе овоцити і спермії, утворюється оваріальна і сперміальна рідина. Овуляція відбувається або зразу в усьому ястику, і тоді риба здатна до одноразового нересту, або вона захоплює тільки частину дозріваючих овоцитів, і тоді риба здатна до багаторазового порційного нересту. В зв'язку зі змінами в екології розмноження можливий перехід від одночасного до порційного ікрометання у ряду видів риб. Проте це явище належить розглядати як виключення. Сперміація відбувається, як правило, в меншому масштабі, зрілі спермії виявляються в сім'яниках тривалий час і самці звичайно беруть участь в нересті неодноразово, що досить широко використовується за штучного відтворення.
Плодючість самок окремих видів риб досить суттєво відрізняється, що характерно і для розмірів ікринок певних видів. Акулові мають дуже невисоку плодючість, але більша частина нащадків виживає завдяки їх розвитку всередині материнського організму або в середині міцної яйцевої оболонки. Жовток яєць скатів і акул схожий за формою і за складом до пташиного, він оточений справдішнім «білком». Яйця акулових розвиваються досить тривалий час — іноді до двох років. На відміну від розглянутого, дрібна ікра риб, які характеризуються високою плодючістю, гине масово. Але у загальному визначенні плодючість костистих риб коливається в широченних межах, що обумовлено об’єктивними умовами виживання. У видів, що мають крупну ікру — лососей, нототеній — на кожен грам маси припадає по одній ікринці, у живородних на один грам припадає декілька десятків ікринок, у коропових — декілька сотень, а у деяких морських риб, що мають дуже дрібну ікру, на 1 грам тіла — по тисячі ікринок.
Кількість сперми і вміст у ній сперміїв в різних видів риб значно відрізняється. Загальна кількість сперми, яка продукується рибою протягом репродуктивного періоду, може перевищувати масу її сім'яників. Ця обставина в значній мірі пояснює реальну можливість багаторазової участі самців в нересті. При цьому тканини сім'яників продовжують продукувати сперму. У плідників райдужної форелі протягом 40 діб нерестового періоду можна отримати до 77 мл сперми (20 еякулятів), в щуки значно менше — до 6 мл (8 еякулятів), у крупних плідників товстолобиків можна отримати за один раз до 25 мл сперми, у осетрових — до 1 л. Об'єм сперматозоїдів в спермі (сперматокріт) теж досить різний. У лососевих сперматозоїди складають 25% сперми, у коропа — 45, у морського карася — 11, а в деяких камбал — навіть 97%. В сім'яниках спермії нерухомі. їх активізація відбувається лише при розбавленні секретом придатка сім'яника, а для деяких риб необхідні особливі умови активації. Встановлено, що спермії форелі не активуються в кислому середовищі, інактиваторами рухливості сперміїв в деяких видів стають іони калію, яких багато в сперміальній рідині.
В науковій літературі, присвяченій відтворювальній системі риб, поряд із сім'яниками і яєчниками часто використовується термін — гонади стосовно до статевих залоз самок і самців. Можливі і вирази типу — гонади самок або гонади самців. Функціонально гонади риб є продуцентами яєць (ікринок) і сперматозоїдів.
Ікринки і сперматозоїди утворюються з первинних статевих клітин, диференціація яких здійснюється ще в період ембріонального розвитку, їх розміри становлять 9-20 мкм. Довжина сперматозоїдів, здатних запліднювати ікру, коливається у різних видів риб від 30 до 60 мкм, яйця мають різні розміри — від частки міліметра до декількох сантиметрів.
Різні види риб стають статевозрілими в різному віці, навіть у риб одного виду в межах ареалу вік досягнення статевої зрілості може варіювати в широких межах, деякі відмінності спостерігаються і у видів однієї популяції. Явище, що розглядається, необхідно сприймати як адаптацію виду в процесі філогенезу до умов середовища з метою оптимізації відтворення і існування виду. В зв'язку з цим очевидну зацікавленість у деяких видів риб становить вік досягнення їх статевої зрілості.
Більш раннє дозрівання характерне для видів з коротким життєвим циклом, риби з більш подовженим циклом дозрівають пізніше. Струмкова і райдужна форель дозріває у віці 3-4 років, пелядь на 4-5 році життя, чудський сиг у віці 2 років, щука досягає статевої зрілості на 3-4 році життя. Білий товстолобик за відповідних умов дозріває в три-п'ятирічному віці, білий амур — в 6-7 років, на півдні України — в п'ятирічному віці. Лин, сазан і карась стають статевозрілими на 3-4 році життя, сом досягає зрілості на 4-5 році, судак — у віці 3-4 роки. В нересті беруть участь самиці окуня від 2 років і старше, форелеокунь стає статевозрілим у віці 4-5 років, змієголов дозріває на третій рік життя, лобан — на шостий-восьмий, гостроніс — на третьому році життя.
При цьому необхідно відмітити, що більшість коропових, окуневих і лососевих риб досягають статевої зрілості у віці 2-6 років. Поряд з цим осетрові стають статевозрілими в 6-12-річному віці, окремі ж види в межах ареалу, що входять до складу даних рядів, дозрівають у 18-річному віці. При цьому, як свідчать дані деяких авторів, які співпадають з рибницькою практикою, статева зрілість в самців настає на 1-2 роки раніше, ніж у самок, окремі статевозрілі особини певних видів риб нерестують не кожного року.
Абіотичні і біотичні фактори, які в ряді випадків тісно пов'язані, а то і обумовлені антропогенним впливом зовнішнього середовища, можуть мати за певних умов визначальне значення. За наявності рівних факторів виключно значима роль належить термічному режиму і забезпеченню риб відповідними кормами. В залежності від факторів середовища, розвиток статевих клітин може прискорюватися або уповільнюватися, а в деяких випадках навіть зупинятись.
Розглядаючи вік досягнення статевої зрілості виду в межах ареалу, можна помітити наявність трьох груп риб, що не пов'язано з видовою приналежністю, а зумовлено загальними закономірностями. Південні популяції дозрівають раніше; ті що займають проміжне географічне положення — пізніше, а найбільш пізнє дозрівання характерне для північних популяцій. При цьому коливання окремих особин в межах ареалу можуть досягати 2-3 років, що доцільно враховувати при організації штучного відтворення. Цікавий і той факт, що порушення в часі настання статевої зрілості під впливом факторів середовища можуть відбуватися не тільки у особин одного виду з різних популяцій, але і в одних і тих же видів даної популяції. Так, найбільш крупний представник ряду осетроподібних — білуга, досягає статевої зрілості у віці від 10 до 18 років, що орієнтує на формування стад в процесі доместикації.
В зв'язку з динамікою розвитку статевих залоз, найбільшу зацікавленість, з нашої точки зору, становить інформація, яку пропонує А.П.Іванов, де стадії стиглості статевих залоз самок і самців поєднуються з гістологією, що характеризує фактичне відображення процесу.
Динаміка розвитку яйцеклітин (овогенез) і сперматозоїдів (сперматогенез) в жіночих і чоловічих статевих залозах — тривалий і складний процес, нормальний перебіг якого залежить від цілої низки зовнішніх і внутрішніх факторів.
Кожна статева клітина в процесі свого достигання повинна пройти кілька послідовних стадій. При цьому доцільно відрізняти два періоди, перший — до досягнення статевої зрілості, починаючи від появи первинних статевих клітин і закінчуючи утворенням зрілих статевих продуктів, та другий — періодичне дозрівання певної частини статевих продуктів протягом міжнерестового періоду. В першому випадку розглядається процес формування зрілих яйцеклітин і сперматозоїдів в статевих залозах самиць і самців, починаючи від ювенального до статевозрілого віку. В другому — ми розглядаємо аналогічний процес, але з особинами, що досягли статевої зрілості, вже приймали участь в нересті і мають нащадків. Перший період досить тривалий і пов'язаний з віком досягнення статевої стиглості конкретних видів риб і відповідними екологічними умовами. Другий, що характеризує закономірне достигання вже статевозрілих особин через певний інтервал часу, значно коротший, але тривалість його може бути несхожою у різних видів риб. Зокрема, лящ, сазан, судак і багато інших видів риб розмножуються щорічно, осетрові — іноді через 3-5 років, а деякі види — через 9 років. Виключення становлять види, які нерестують один раз в житті, після чого гинуть, це тихоокеанські лососі — кета, горбуша.
Довготривалі дослідження в галузі динаміки розвитку репродукційної системи у риб здійснювались на базі видів, які відносяться до різних систематичних груп, що обумовило наявність деяких відмінностей, пов'язаних з видовою специфікою. Таким чином, не дивлячись на спільність підходів до цієї проблеми ряду авторів, шкала зрілості риб під впливом видової специфічності не має універсальності, що затруднює, а то і унеможливлює її використання.
Згідно з даними, які наводить А.П.Іванов, для коропових і окуневих існують шкали зрілості, запропоновані іншими провідними вченими для основних об’єктів аквакультури. Кожна із цих шкал становить виключний науковий інтерес, має очевидне практичне значення, але діапазон використання її обмежений, можливе застосування тільки для даних груп риб. Не беручи під сумнів гідність цих шкал, О.Ф.Сакун і Н.А.Буцька розробили дві універсальні шкали для абсолютної більшості промислових груп риб. Перша шкала для самок відображує загальний хід достигання статевих продуктів, а друга — у самців. Запропоновані оптимізовані і адаптовані шкали дозволяють визначати стадії зрілості яєчників і сім'яників за зовнішнім виглядом і гістологічною будовою. На основі цих двох шкал розроблена універсальна шкала стиглості статевих залоз самок і самців з коротким описом овогенезу і сперматогенезу.
Описана шкала стадій стиглості статевих залоз, як відзначає А.П.Іванов, може бути застосована при роботі з рибами, яким притаманна одночасна ікровіддача. Проте є ряд видів риб, для яких характерне порційне ікровикидання. У цих риб овоцити дозрівають асинхронно і це явище асинхронності розвитку овоцитів проявляється в період трофоплазматичного росту (ІІІ стадія стиглості яєчників). При переході яєчників в VI стадію стиглості не всі овоцити одночасно закінчують трофоплазматичний ріст.
У зв'язку з цим після того, як самка відкладе першу порцію ікри, яєчник переходить не в VI стадію, а в ІІІ стадію стиглості, яку позначають Зг або 6-3г. На цій стадії в яєчниках присутні фолікули, що луснули, і овоцити протоплазматичного росту. Після відкладання самкою другої порції ікри яєчник переходить в ІІІ, стадію стиглості. Така зміна стадій стиглості яєчників порційно нерестуючих видів риб продовжується до того часу, доки самка не відкладе останню порцію, що є умовою переходу яєчників в VI стадію зрілості.
Для самців з порційним нерестом характерна певна розтягнутість процесу, а сперматогенез продовжується весь нерестовий період, відповідно змінюються і стадії стиглості сім'яників.
В завершальній стадіях розвитку яйцеклітин характерне інтенсивне накопичення жовтка і формування оболонок, а у сперматозоїдів в цей період утворюється хвіст, формується система руху і укорінення в яйцеклітину. Особливістю динаміки маси статевих залоз є те, що в міру наближення нересту їх маса різко збільшується, а накопичені в них речовини в загальному балансі речовин в організмі досить суттєві.
Зміни відносної маси статевих залоз самців і самок одного виду проходять паралельно. Відносна маса зрілих гонад значно більша в самок, ніж в самців. При цьому, для деяких видів риб маса гонад може складати більше 30% загальної маси риб, що знаходиться в переднерестовому стані.
Тропічні види риб, які багаторазово нерестують протягом року, продукують загальну масу ікри, значно переважаючу масу тіла риби. Статеві залози риб мають витягнуту форму і в більшості видів представлені парним органом, підвішеним в порожнині тіла на брижах. В процесі овуляції яйцеклітини виходять із фолікул через розриви і потрапляють в порожнину яєчника або безпосередньо в порожнину тіла, звідки виводяться назовні (у воду). В порожнину тіла з оболонки гонад ікра і сперма видаляються у круглоротих. Потім яйцеклітини і сперматозоїди через пори потрапляють в сечовий синус або сечовипускний канал і виводяться назовні через клоаку або урогенітальну папілу. Осетрові і акулові риби виводять ікру або яйця через мюллерові протоки, сформовані з сечових протоків головної нирки. Сперма із них виводиться по ниркових канальцях, які у акул перетворюються в специфічний айдигів орган.
В абсолютної більшості костистих риб кожен яєчник має свій яйцепровід, який закінчується генітальною порою. У окремих видів лососевих риб, які мають ікринки великого діаметру, статеві залози під час достигання розриваються, і ікра опиняється в порожнині тіла, звідки вона через широкий лійкоподібний отвір яйцепроводу виводиться в зовнішнє середовище. Сім'яники костистих риб мають власні сім'япроводи, які відкриваються в сечовик. В процесі сперміації стиглі сперматозоїди звільняються із тканини сім'яника і виводяться в зовнішнє середовище через сім'япроводи.
Жіночі статеві залози не тільки продукують яйцеклітини, в яєчниках синтезуються жіночі статеві гормони, представлені естрогенами і прогестероном. Естрогени у нижчих хребетних тварин виділяються у вигляді 17-естрадіолу або естрону. В крові вони знаходяться в сполуці з білком. Найбільш активний — естрадіол. Естрогени можуть вироблятися і в сім'яниках (інтерстиціальними клітинами), а також тканинами надниркових залоз.
В невеликих концентраціях естрогени стимулюють диференціювання фолікул, а в значних дозах — пригнічують цей процес, знижуючи секрецію фолікулостимулюючого гормону. Вони стимулюють синтез білка, сприяючи затримці в організмі натрію, кальцію, фосфатів, води і справляють позитивний, стимулюючий вплив на процес появи вторинних жіночих статевих ознак, які обумовлюють статеву поведінку. Прогестерон стимулює вироблення фолікулостимулюючого гормону.
Чоловічі статеві клітини продукують специфічні чоловічі гормони — андрогени, найбільш активним з яких є тестостерон. Виробляються андрогени інтерстиційними клітинами Лейдіга. Вплив андрогенів, в певній мірі, визначає особливості будови тіла самців, поява шлюбного вбрання, розвиток гоноподій у деяких живородних риб, а також відповідні етологічні особливості, сприяє росту тканин і синтезу білків, підвищує інтенсивність еритропоезу, посилює кровотік в тканинах. Низька концентрація гормону посилює сперматогенез, а підвищена — гальмує. Андрогени можуть також продукуватись яєчником і корою надниркових залоз.
Очевидну теоретичну зацікавленість і практичну значимість представляють дані, що висвітлюють питання статі риб. У риб відсутня одноманітна система первинних спадкових структур, які визначають стать особини. Для одних видів риб стать визначається, як і у людини, (ХУ) системою хромосом. Так у рибця-самиці в диплоїдному наборі виявлено дві однакові хромосоми (XX), а самці мають дві різні хромосоми (ХУ). Для японських йоржів присутність різних хромосом (ХУ) свідчить про те, що це самка, а пара однакових (XX) — властива самцям.
У деяких видів реалізація статі залежить від наявності чи відсутності статевих хромосом в диплоїдному наборі. Так, зокрема, самки фундулуса мають дві статеві хромосоми (XX), а в самців хромосоми відсутні, тоді як у лабіринтових рибок-коллізій статеві хромосоми присутні тільки в самців.
В абсолютній більшості випадків стать у риб визначається поліченою структурою цієї ознаки, а генетичні структури, які визначають статеву належність розселені в декількох хромосомах (короп, лососі, осетрові). В ряді випадків система визначення статі різна у близьких видів, зокрема, у вугрів, а то і у різних популяцій одного виду.
Зовнішній прояв ознак чоловічого або жіночого типу розвитку знаходить своє вираження в першу чергу у формуванні статевих залоз — сім'яників або яєчників. На ранній стадії онтогенезу зовнішній вигляд і внутрішня будова риби, як правило, не дозволяє віднести особину до тієї чи іншої статі, і особини називаються ювенальними. Поряд з цим, ранні стадії розвитку репродуктивної системи риб досить складні і багатогранні. Особливістю первинних статевих клітин у риб є те. що вони здатні розвиватися як за чоловічим, так і за жіночим типом.
У деяких видів риб ця особливість обумовила ювенальний гермафродитизм, коли одночасно розвиваються як чоловічі, так і жіночі статеві клітини, одні із яких пізніше пригнічуються, не впливаючи в подальшому на стать особини. Поряд з цим досить часто спостерігаються такі явища, як протоандрія і прототинія, що характеризуються тим, що на певній стадії розвитку всі статеві клітини розвиваються тільки за чоловічим або за жіночим типом. При цьому виключно значима роль гормонів; фактичне співвідношення чоловічих та жіночих статевих гормонів в організмі риби визначає напрямок розвитку первинних статевих клітин.
Інтерстиційні клітини статевих залоз, окрім поживної і опорної функцій, несуть ще й ендокринне навантаження, секрети статевих залоз мають величезний вплив не тільки на репродуктивну систему, але і на весь організм.
Поряд з типовим характером функціонування репродуктивної системи, у риб відоме і явище гермафродитизму, яке функціонально диференціюється на декілька типів. Таке явище досить розповсюджене серед окунеподібних, анчоусів, що світяться і коропозубих. При цьому є види риб, які, будучи в ранньому віці самками, пізніше стають самцями, а для деяких видів характерна протилежна картина. Ця особливість базується на тому, що в статевих залозах однієї й тієї ж особини, йде одночасний розвиток як чоловічих, так і жіночих статевих клітин, а особинам властива поведінка чоловічого чи то жіночого типу. Окремі види морських окунів характеризуються тим, що у них в статевих залозах відбувається одночасний розвиток яйцеклітин і сперматозоїдів, а тип шлюбної поведінки в процесі репродуктивного циклу може змінюватись протягом декількох хвилин, що супроводжується зміною шлюбного забарвлення.
Велику зацікавленість як в теоретичному, так і в практичному плані, має таке явище, як інверсія статі. Статеві гормони в процесі формування особин відіграють суттєву роль в становленні статі у риб. Використання статевих гормонів дозволяє змінювати стать, перетворювати самців в самиць і навпаки. Іншими словами, отримана реальна можливість забезпечити ефект інверсії статі в експериментальних і виробничих умовах, що в ряді випадків має практичне значення. В практиці для інверсії статі використовують статеві гормони і їх аналоги. Речовини, аналогічні за впливом з чоловічими гормонами, які викликають маскулінізацію (перетворення в самців), іменуються андрогенами, а речовини, що викликають фемінізацію (перетворення в самок), іменують естрогенами. Кращі результати інверсії отримані при використанні особин раннього постембріогенезу — стадії личинки. Практично цілковите перетворення самиць в самців відмічене за згодовуванні кормів, які мали в наявності метилтестостерон біля 50 мг/г. В експериментах період коливався від 20 до 120 діб. В процесі дослідження встановлено, що перетворення самців в самиць відбувається при згодовуванні корму з різними естрогенами (етіпілестрадіолу, естрону, естрадіолу) в кількості 20-60 мг/кг протягом 20-140 діб.
Підсумовуючи сказане, необхідно відмітити, що відтворювальна система риб різних видів характеризується достатньою спільністю анатомії, гістології та фізіології, але це не виключає наявності цілого ряду видоспецифічних особливостей, які з'являються в процесі філогенезу, і стійко зберігаються в онтогенезі.