ОСНОВИ МЕДИЧНОЇ БІОЛОГІЇ - 2012
Морфологія клітини. Структурні компоненти цитоплазми та ядра
Клітинна теорія. Вивчення умовної норми життєдіяльності клітин є однією з необхідних сходинок у розумінні і пізнанні патологічних процесів, які студенти продовжуватимуть вивчати на кафедрах патологічної анатомії і фізіології, внутрішніх і хірургічних хвороб, а також на кафедрах гігієнічного профілю, де хвороби вивчаються на організмовому, системному, тканинно-органному рівнях.
Клітинна теорія, яка стверджує, що клітина є елементарною одиницею будови і розвитку всіх організмів, була створена в 1839 році (Т. Шванн і М. Шлейден).
До 30-х років ХІХ ст. було опубліковано багато робіт про клітинну будову організмів. Загально визначеним стало уявлення про клітину як елементарну мікроскопічну структуру рослин. Німецький ботанік М. Шлейден першим прийшов до висновку, що у будь-якій рослинній клітині є ядро. Німецький зоолог Т. Шванн у 1939 р. опублікував працю «Мікроскопічні дослідження про відповідність у структурі та рості тварин і рослин». У цій роботі були закладені основи клітинної теорії. На подальший розвиток вчення про клітину мала праця Р. Вірхова «Целюлярна патологія». Р. Вірхов увів у науку ряд нових уявлень про роль клітинних структур в організмі та вивів основні положення, які блискуче підтвердились: «кожна клітина — з клітини», «поза клітиною немає життя», «найбільше значення в життєдіяльності клітин має не оболонка, а її вміст: цитоплазма й ядро».
Уявлення про клітину як найменшу одиницю життя доведено Т. Шванном та Р. Вірховим, які вважали, що кожна клітина характеризується всіма властивостями живого.
Основні положення клітинної теорії зберегли своє значення до теперішнього часу, хоча за останні роки були одержані нові дані про структуру та життєдіяльність клітин. Сучасна клітинна теорія включає такі положення:
1. Клітина - елементарна структурно-функціональна та генетична одиниця живого.
2. Клітини різних організмів гомологічні за своїм походженням, будовою, хімічним складом, основними проявами життєдіяльності.
3. Розмноження клітин відбувається шляхом ділення вихідної (материнської) клітини.
4. У багатоклітинних організмів, які розвиваються з однієї клітини - зиготи, спори, тощо - різні типи клітин формуються завдяки їхній спеціалізації в процесі онтогенез організму та утворюють тканини.
5. Із тканин складаються органи, які об’єднані між собою та підпорядковані нервово- гуморальній та імунній системами регуляції.
Відповідно сучасним уявленням живі організми являють собою відкриті, самокорегуючі та самовідтворюючі системи, важливими компонентами яких є нуклеїнові кислоти та білки. В ДНК закодована інформація про структуру білків, в свою чергу білки -складні молекули, які мають функціональну специфічність та визначають у першу чергу реалізацію генетичної інформації і властивості живого: здатність до саморепродукції та саморегуляції, забезпечення потоку речовин та енергії під час обміну, спадковість та мінливість, чутливість, рух, адаптацію. Така сукупність властивостей вперше визначається на клітинному рівні, тому клітина є елементарною структурно-функціональною одиницею життя.
Неклітинні і клітинні форми життя
В органічному світі можна виділити 2 форми життя: клітинні і неклітинні.
Неклітинні форми життя — це віруси, що утворюють царство Vira, яке поділяється за нуклеїновими кислотами на рибовіруси та дезоксирибовіруси. Віруси людини і хребетних тварин відносяться до 17 родин, з них 6 - ДНК-вмісні віруси (ДНК-геномні) та 11 - РНК- вмісні (РНК-геномні). Віруси -- це форми живого, які проявляють властивості живого тільки паразитуючи в клітинах про- та еукаріот. Віруси мають мікроскопічні розміри, але коливаються у широких межах: від 15 - 18 нм до 300 - 400 нм та мають різноманітну форму: паличкоподібну, сферичну, сперматозоїдну та ін.
Віруси відрізняються від про- та еукаріотів структурною організацією та функціональними особливостями. Вони містять тільки одну з нуклеїнових кислот — ДНК або РНК, не мають клітинної будови. Віруси не здатні до росту та бінарного поділу. Вони формуються, набуваючи визначеної форми та точних розмірів, які не змінюються. Саморепродукція вірусів відбувається в клітинах шляхом реплікації НК. У вірусів відсутній автономний процес обміну речовин, немає власних систем накопичення енергії та синтезу білків, вони використовують ферментні системи, енергетичні ресурси живих клітин та переключають роботу клітин на виробництво нових вірусних часточок.
Віруси існують у двох формах: позаклітинній — віріон і внутрішньоклітинній — вірус. За будовою поділяються на прості та складні.
Віріони, які складаються з НК і білка, називають простими. НК вкрита ззовні білковою оболонкою — капсидом, який виконує захисну функцію — ізоляцію геному вірусу від зовнішнього середовища. Разом НК та білок утворюють нуклеокапсид.
Складні віріони мають НК, білок, ліпіди, вуглеводи та білки-ферменти. Ліпіди, вуглеводи та білки входять до складу зовнішньої оболонки, яку називають суперкапсидом.
Вірусні ДНК мають молекулярну масу в 10-100 разів меншу, ніж молекулярна маса ДНК бактерій. У геномі містяться до кількох сотень генів. ДНК вірусів можуть бути одноланцюговими або дволанцюговими. Вони здатні замикатися в кільце і в такій формі реплікуються, транскрибуються, набувають стійкості до ферментів — ендонуклеаз і вбудовуються у клітинний геном.
Вірусні РНК не відрізняються від РНК клітин. Одноланцюгові РНК залежно від функцій поділяються на 2 групи. Одні здатні виконувати функцію і-РНК (позитивний геном), інші можуть бути матрицею для утворення і-РНК в процесі зворотної транскрипції (негативний геном).
Віруси, які пристосувалися до паразитування в клітинах бактерій називаються бактеріофагами. Фаги мають більш складну будову, ніж більшість вірусів. В них розрізняють головку і відросток. На дистальному кінці відростка може бути базальна пластинка і нитки, які здійснюють процес адсорбції фага до бактеріальної клітини. Капсид голівки фага побудований за кубічним типом симетрії, а відросток — за спіральним. В голівці є нуклеїнова кислота. При проникненні в клітину бактерії, фаги вприскують нуклеїнову кислоту через канал відростка, а білки залишаються поза клітиною. В клітині фагова НК починає інтенсивно розмножуватись, викликаючи її лізис. Інколи проникнення фагів в клітину не супроводжується лізисом бактерії, а ДНК фагів включається в геном бактерій і передається її нащадкам.
Віруси мають велике значення, так як більшість з них викликають хвороби рослин, тварин і людини. До вірусних хвороб людини відносяться: віспа, тайговий енцефаліт, кір, грип, снід та ін. Крім того, віруси можуть забезпечувати перенесення генетичного матеріалу (трансдукція).
Клітинні форми життя, в залежності від наявності сформованого ядра поділяються на прокаріот та еукаріот.
Прокаріоти — організми, клітини яких не мають ядра.
Еукаріоти — організми, клітини яких мають одно або декілька ядер.
Прокаріоти — окреме надцарство організмів, до якого відносять різні групи бактерій та ціанобактерії (синьо-зелені водорості).
Клітини прокаріот дрібні за розмірами (0,1-0,25 мкм — мікоплазми; 1-10 мкм — бактерії, синьо-зелені водорості). Прокаріотичні клітини мають клітинну оболонку, цитоплазму. Оболонка представлена клітинною стінкою складної будови та цитоплазматичною мембраною. На поверхні може утворюватися слизова капсула.
В цитоплазмі знаходяться органели: рибосоми та мембранні утворення — мезосоми, фотосинтезуючі мембрани, мембранні вирости, пухирці.
В середині клітини виділяється зона локалізації спадкового матеріалу нуклеоїд, де знаходиться бактеріальна хромосома — кільцева молекула ДНК, яка не з’єднана з білками гістонами.
Прокаріотична клітина не має типових мембранних органел: ЕПС, комплекса Гольджі, мітохондрій, пластид, вакуоль, лізосом. Також відсутні центріолі, слабо розвинена система мікротрубочок та мікрофіламентів.
Прокаріотичні клітини мають інтенсивний обмін речовин, в них частіше відбуваються мутації та швидше відбуваються процеси адаптації до середовища. Ділення клітин — дробіння (бінарний поділ) з попереднім подвоєнням молекули ДНК.
До еукаріот відносяться представники трьох царств живого: Рослини, Гриби, Тварини. Їх клітини мають ядро, мембранні органели (ЕПС, комплекс Гольджі, лізосом та інш.), розвинені немембранні органели (рибосоми, система мікротрубочок та мікрофіламентів, центріолі). Спадковий матеріал представлений лінійними хромосомами, які включають ДНК, гістони, кислі білки.
Основний спосіб ділення -- мітоз -- складний еволюційно закріплений процес ділення, який забезпечує точну передачу та розподіл спадкового матеріалу.
Структурно-функціональна організація еукаріотичної клітини
Еукаріотичні клітини побудовані з трьох основних компонентів: мембрани, цитоплазми та ядра. Виділяють 2 підтипа клітинної організації: одноклітинні організмами та клітини багатоклітинних організмів (рослин, тварин, грибів).
Порівняльна таблиця про- і еукаріот
Клітинні структури |
Еукаріоти |
Прокаріоти |
Клітинна стінка |
У рослин та грибів клітинні стінки жорсткі і містять полісахаріди (основний компонент: у рослин - целюлоза, у грибів - хітин). Клітини тварин не мають стінок. |
Жорсткі містять полісахаріди й амінокислоти. Основний компонент - мурен. |
Ядро |
Ядро відділене від цитоплазми ядерною мембраною. |
Відсутні |
Спадковий матеріал |
Лінійні хромосоми, ДНК-гістони, кислі білки |
Кільцева ДНК |
Органели ЕПС |
Органел багато. Одномембранні: ЕПС, комплекс Гольджі, лізосоми, вакуолі, пероксисоми. Двомембранні: ядро, мітохондрії, хлоропласти. Немембранні: рибосоми, центріолі, мікротрубочки та мікрофіламенти. |
Органел мало або відсутні; немембранні. Рибосоми дрібні, чутливі до антибіотиків. |
Цитоплазма |
Добре розвинутий цитоскелет, рух цитоплазми, ендо- й екзоцитоз. |
Не має цитоскелета, руху цитоплазми |
Дихання |
Аеробне - у мітохондріях. |
Анаеробне, або аеробне. У бактерій - в мезосомах, у синьо-зелених водоростей - на цитоплазматичних мембранах. |
Поділ клітин |
Мітоз (або мейоз). |
Простий поділ, мітозу не має. |
Хімічний склад живих організмів відносно подібний. Всі хімічні елементи, що входять до складу клітин поділяють на чотири групи:
- макроелементи (вміст яких від 1% та вище): кисень, вуглець, азот, гідроген, кальцій та фосфор;
- олігоелементи (вміст 0,1-1%): калій, натрій, хлор, сірка, магній та залізо;
- мікроелементи (вміст менше 0,01%): цинк, марганець, кобальт, фтор, бром, йод;
- ультрамікроелементи (концентрація від 10-4 до 10-6 %) - належить решта елементів: бор, літій, алюміній, кремній, кадмій, селен та ін.
Подібність елементного складу живої і неживої природи свідчить про спільність їхнього походження.
Серед неорганічних сполук живих організмів вода виконує особливу роль. Вона є основним середовищем де відбуваються всі процеси обміну. Її вміст у більшості живих організмів становить 60 - 80 %, а у деяких (медузі) - до 98 %. Вода має унікальні хімічні та фізичні властивості. У неї відносно високі температури кипіння, плавлення та випаровування, що зумовлено взаємодією між молекулами води. Молекула води (Н2О) полярна. В ній два атоми водню сполучаються з атомом кисню міцним ковалентним зв’язком, що виникає за рахунок утворення спільної пари електронів по одному від кожного атома. Молекула води електронейтральна, але на її полюсах розміщені позитивний і негативний заряди (полярність). Саме тому молекули води можуть притягуватись одна до одної за рахунок сил електростатичної взаємодії між частково негативним зарядом на атомі кисню однієї молекули та позитивним зарядом на атомі водню іншої. Такий тип зв’язку називається водневим, він у 15-20 разів слабший за ковалентний.
Вода значно кращій розчинник, ніж більшість інших рідин. Тому всі речовини поділяють на таки, що добре розчинні у воді - гідрофільні (багато кристалічних солей, кислоти та ін.), не розчинні - гідрофобні (ліпіди, деякі білки та ін.) та амфіфільні (поєднують властивості обох класів). Більшість хімічних реакцій відбуваються тільки у водних розчинах. Вода визначає фізичні властивості клітини: об’єм, внутрішньоклітинний тиск. З нею пов’язана регуляція теплового режиму організмів. Їй властива висока теплоємність, тобто здатність поглинати тепло за незначних змін своєї температури. Саме завдяки цьому вода забезпечує в клітині терморегуляцію.
Речовини, які мають скелети з ковалентно зв’язаних атомів вуглецю називають органічними сполуками. Майже всю суху речовину клітини складають білки, вуглеводи, ліпіди та нуклеїнові кислоти. Це відносно великі структури з високою молекулярною масою - макромолекули (біополімери).
Серед клітинних макромолекул за функціональним значенням і в кількісному відношенні провідна роль належить білкам. У тваринній клітині вони становлять 40-50%. Білки - це високомолекулярні полімери, мономерами яких є амінокислоти. Велика різноманітність білків забезпечується комбінаціями лише 20 амінокислот, які називають основними. Усі амінокислоти, що входять до складу білків, мають спільну групу атомів, яка складається з аміногрупи (- NH2), для неї характерні лужні властивості, та карбоксильної групи (- СООН) з кислотними властивостями, які зв’язані одним і тим самим атомом вуглецю. А відрізняються одна від одної бічними ланцюгами (радикалами, чи R-групами), які у різних амінокислот неоднакові за хімічною структурою, електричним зарядом, розчинністю у воді. Амінокислоти класифікують відповідно до можливості їхнього синтезу в організмах - замінні та незамінні. Перші синтезуються в організмі людини чи тварин, а другі потрапляють до них лише з харчовими продуктами (їх синтезують рослини, гриби, бактерії). Білкові молекули утворюються за допомогою пептидних зв’язків між аміногрупою однієї амінокислотами та карбоксильною групою іншої. З’єднані в процесі трансляції амінокислоти утворюють поліпептид. До складу простих білків входять тільки амінокислотами. До складу складних білків - ліпіди (ліпопротеїди), вуглеводи (глікопротеїди).
Білки відрізняються між собою не тільки кількістю мономерів та складом амінокислот, але й послідовністю їх розташування у поліпептидних ланцюгах - первинна структура. Різноманітність варіантів первинних структур білків дуже велика: навіть три пептиди, які складаються з трех амінокислот можуть мати 6 варіантів. При розташуванні у просторі амінокислоти групуються в середині молекули поліпептида, а виникнення локальних водневих зв’язків між пептидними групами призводить до утворення α-спіралей - вторинної структури. Регулярну вторинну структуру мають фібрилярні білки - довгі, видовжені, погано розчинні у воді (колаген, актин, міозин та ін.). Просторове розташування поліпептида у вигляді глобули носить назву третинної структури. Глобулярні білки мають вигляд сфери та добре розчинні у воді (гемоглобін, більшість ферментів). Якщо до складу білка входить декілька поліпептидів - їх взаємне розташування у просторі називають четвертинною структурою.
Основні функції білків: 1) структурна - з білків утворюються всі елементи, як клітини, так і організму в цілому; 2) захисна - білки створюють захисні покриви організму, утворюють антитіла; 3) регуляторна - білки-гормони беруть участь у регуляції багатьох метаболічних процесів; 4) каталітична - білки-ферменти прискорюють хімічні реакції організму та забезпечують усі функції клітин; 5) рецепторна - білки-рецептори сприймають сигнали, перетворюють та передають їх у відповідну ділянку клітини або організму; 6) транспортна функція - білки здатні зв’язувати та транспортувати неорганічні іони і специфічні органічні речовини; 7) скоротлива, або рухова - забезпечує здатність клітин, тканин чи організму змінювати форму, рухатись; 8) енергетична - при розщепленні білків у клітині вивільняється енергія.
Вуглеводи - це органічні сполуки, загальна будова яких відповідає формулі Сх(Н2О)у. У тваринних клітинах вуглеводи присутні в незначній кількості (1-5%), у рослинних їх значно більше (70-90%).
Вуглеводи поділяють на три основні класи: моносахариди, олігосахариди, полісахариди. Моносахариди (прості цукри) мають загальну формулу СпН2пОп. У природі найпоширеніші гексози: фруктоза, глюкоза. Із пентоз більш відомі рибоза та дезоксирибоза, що входять до складу РНК та ДНК. Олігосахариди - полімерні вуглеводи, в яких моносахаридні ланки з’єднані ковалентним (глікозидним) зв’язком. Серед олігосахаридів найпоширеніші дисахариди: сахароза, лактоза, мальтоза та ін. Більшість вуглеводів полісахариди - біомолекули з високим ступенем полімеризації (крохмаль, целюлоза, глікоген та ін.). У живих організмах олігосахариди зустрічаються у комплексі із білками (глікопропеїни) та ліпідами (гліколіпіди).
У живих організмів вуглеводи виконують дві основні функції - енергетичну та будівельну. Полісахариди та олігосахариди розщеплюються до моносахаридів з наступним окисленням до СО2 та Н2О. При повному розкладі граму цих речовин вивільнюється 17,6 кДж енергії. Будівельна функція вуглеводів полягає в тому, що вони входять до складу опорних елементів.
Ліпіди - не розчинні у воді (гідрофобні) органічні сполуки, які можна вилучити з клітини за допомогою неполярних розчинників. Практично всі ліпіди є складними ефірами жирних кислот та спирту. Жири є основною речовиною жирових включень в клітині. Їх вміст становить від 5% до 15% сухої маси, а в жировій тканині - до 90%. Основні функції ліпідів: 1) енергетична - при повному розщепленні 1 г жирів до СО2 і Н2О виділяється 38,9 кДж енергії; 2) будівельна - ліпіди складають основу біологічних мембран, входять до складу нервових волокон; 3) захисна - захист органів від механічних пошкоджень; 4) теплоізоляційна - жири накопичуються у підшкірній жировій клітковині.
Скороченні позначення амінокислот, їх замінність (з) або незамінність (н) для людини.
N n/n |
Назва амінокислоти |
Скорочення |
1 |
Аланін (з) |
Ала |
2 |
Аргінін (з), (незамінна для дітей) |
Арг |
3 |
Аспарагін (з) |
Асн |
4 |
Аспарагінова кислота (з) |
Асп |
5 |
Валін (н) |
Вал |
6 |
Гістидин (з), (незамінна для дітей) |
Гіс |
7 |
Гліцин(з) |
Глі |
8 |
Глутамін (з) |
Глн |
9 |
Глутамінова кислота (з) |
Глу |
10 |
Ізолейцин (н) |
Іле |
11 |
Лейцин (н) |
Лей |
12 |
Лізин (н) |
Ліз |
13 |
Метіонін (н) |
Мет |
14 |
Пролін (з) |
Про |
15 |
Серин (з) |
Сер |
16 |
Тирозин (з) |
Тир |
17 |
Треонін (н) |
Тре |
18 |
Триптофан (н) |
Три |
19 |
Фенілаланін (н) |
Фен |
20 |
Цистеїн (з) |
Цис |
Нуклеїнові кислоти (ДНК, РНК) - складні високомолекулярні біополімери,
мономерами яких є нуклеотиди. Основна функція - збереження та передача генетичної інформації.
Структурні компоненти цитоплазми
Як тваринні, так і рослинні клітини являють собою гетерогенну систему. У клітині розрізняють ядро з ядерцем і цитоплазму, в якій містяться органели та включення. Зверху клітина вкрита оболонкою, або клітинною мембраною
Цитоплазма - це внутрішнє середовище клітини, що міститься між плазматичною мембраною і ядром, та забезпечує взаємодію органоїдів. Вона являє собою колоїдний розчин органічних і мінеральних речовин. Внутрішнє середовище клітини характеризується відносною сталістю будови та властивостей (гомеостаз). У цитоплазмі розрізняють периферичний ущільнений шар - ектоплазму і внутрішній, що безпосередньо прилягає до ядра - ендоплазму. Цитоплазма складається із цитозолю (матрикс), внутрішньоклітинних органел і включень. Цитозоль складає приблизно 55% загального об’єму клітини, не містить органел та становить складну безбарвну колоїдну систему клітини. В складі цитозолю є розчинні білки, вуглеводи, ліпіди. РНК, неорганічні речовини. До неорганічних речовин належать вода, солі, гази. Цитозоль може перебувати у різних станах: рідкому стані - золю та твердому - гелю. Перехід з одного стану в інший називають фазовим переходом. Коли цитоплазма знаходиться у стані золю, спостерігається максимальна швидкість біохімічних реакцій, а у твердому стані швидкість обміну речовим значно менша. Перехід цитозолю з стану золя у стан гелю і навпаки зумовлює циклоз - рух цитоплазми.
Цитоскелет - це сітка білкових фібрил та мікротрубочок, що вкривають зсередини цитоплазматичну мембрану і пронизують внутрішній простір клітини. До складу цитоскелету входять такі структурні елементи: мікротрубочки - утворені білковими фібрилами, що містять тубулін; мікрофіламенти - мають здатність скорочуватись, містять актин; проміжні філаменти - утворені комбінацією кількох філаменів. Функції цитоскелету:
✵ Підтримка об’єму та форми клітини за рахунок фібрілярної сітки, яка вкриває зсередини мембрану (кореткс);
✵ Зміна форми клітини за рахунок скорочення або розтягнення білкових фібріл;
✵ Пересування органел та транспорт везикул;
✵ Утворення мультиферментних компонентів;
✵ Утворення веретена поділу;
✵ Утворення ворсинок та джгутиків у найпростіших;
✵ Утворення міжклітинних контактів - демосом. Мікрофіламенти, які через десмосому проникають у сусідню клітину забезпечують зв’язок між ними.
✵ Забезпечують скорочувальну функцію м’язових волокон (актиноміозиновий комплекс).
Органели цитоплазми (органоїди) — це постійні диференційовані структури цитоплазми, які мають певні функції і будову. Розрізняють органели загального і спеціального призначення.
Органели спеціального призначення характерні для клітин, що виконують спеціалізовану функцію: міофібрили - скоротливі елементи м'язових клітин, війки епітелію в трахеї і бронхах, мікроворсинки всмоктувальної поверхні епітелію тонкої кишки тощо.
До органел загального призначення відносяться: ендоплазматичний ретикулум, рибосоми, лізосоми, комплекс Гольджі, мітохондрії, пластиди, клітинний центр, мікротрубочки.
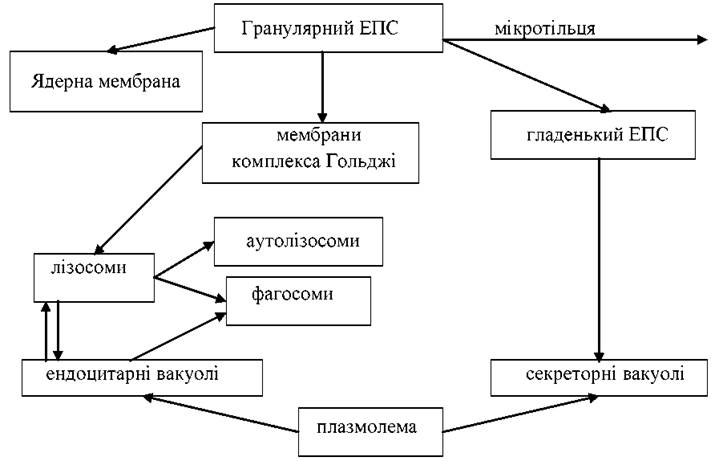
Органели цитоплазми поділяють на мембранні та немембранні компоненти. Мембранні структури цитоплазми являють собою відокремлені або взаємозв’язані відсіки, вміст яких відділено мембранами, від гіалоплазми та плазмолеми. Це замкнуті об’ємні зони (компартменти), які спеціалізовані для виконання певних функцій. Мембранні структури цитоплазми можна поділити на дві групи:
1. Канальцева і вакуолярна системи, до яких відносять ендоплазматичний ретикулюм, вакуолі, комплекс Гольджі та лізосоми, мікротільця, одномембранні органели.
2. Двомембранні напівавтономні органели — мітохондрії та пластиди. Вони мають зовнішню та внутрішню мембрани, які не переходять одна в одну на відміну від двомембранної ядерної оболонки, в якій зовнішня мембрана може з’єднуватися з мембранами ендоплазматичного ретикулюма.
Немембранні органели — це рибосоми, центріолі, мікротрубочки, мікрофіламенти, міофібрили.
Канальцева і вакуолярна система представлена трубчастими або сплющеними порожнинами (цистернами) відмежованими мембраною від гіалоплазми і розміщеними по всій цитоплазмі. Цистерни іноді мають пухирчатоподібні розширення.
Ендоплазматична сітка (ретикулум) створена сіткою мембранних трубочок, цистерн і овальних везикул. Цистерни можуть утворювати щільно упаковані шарові структури — ділянки найактивнішого білкового синтезу, які називають ергоплазмою. Розрізняють два типи ЕПС: зерниста (гранулярна) та гладенька (агранулярна). На мембрані зернистої ЕПС багато рибосом (полісом), тому на ній синтезуються білки. Агранулярна ЕПС не має рибосом. Вона приймає участь в обміні жирів, вуглеводів, стероїдних гормонів та інших речовин. В канальцях і цистернах відбувається накопичення і транспорт речовин від місць синтезу в зони формування гранул. Безпосереднім продовженням ЕПС є ядерна мембрана та зовнішня мембрана (плазмолема). ЕПС збільшує площу внутрішніх поверхонь клітини, поділяючи її на відсіки.
В клітинах різних органів ЕПС можуть виконувати специфічні функції. Наприклад, в печінкових клітинах з розвиненою гладенькою ЕПС руйнуються та знешкоджуються токсичні речовини, деякі лікарські препарати. У пухирцях та канальцях гладенької ЕПС послугованої мускулатури депонуються Са2+, необхідних для процесу скорочення.
Система вакуолярних структур цитоплазми являє собою єдине ціле, окремі елементи якої можуть переходити з одного в інший. Так зовнішня мембрана ядерної оболонки переходить у мембрани гранулярного ЕПС, мембрани останнього -- у мембрани агранулярного ЕПС і т.д.
Пластинчатий комплекс Гольджі утворений диктиосомами (від декількох десятків до тисяч на клітину). Диктиосома утворена з 3-12 плоских дископодібних цистерн, по краях яких відокремлюються пухирці (везикули). Деякі ділянки — розширення цистерн утворюють більші пухирці (вакуолі). На комплексі Гольджі утворюються секреторні пухирці та вакуолі, що містять білки та інші сполуки, які виділяються клітиною. Попередник секрету (просекрет) надходить у диктиосому з зони синтезу, тут відбуваються деякі хімічні модифікації, відокремлюється мембраною на порції (сегрегація).
Комплекс Гольджі накопичує, дегідратує та виводить із клітини продукти обміну речовин, бере участь у синтезі лізосом, пероксидом, в утворенні акросоми сперматозоїда, синтезує зерна жовтка в яйцеклітинах, бере участь в утворенні клітинної перетяжки при цитокінезі. В ньому синтезуються полісахариди, глікопротеїни, гліколіпіди.
Мітохондрії — двомембранні, напівавтономні органели, які є в усіх еукаріотичних клітинах; округлої або паличкоподібної форми, іноді розгалуджені; розміри: товщиною 0,5 мкм, довжиною — 5,0-10,0 мкм для більшості клітин. У будові мітохондрій розрізняють дві мембрани: зовнішню і внутрішню. Зовнішня мембрана гладенька без складок і виростів, має великі пори. Внутрішня мембрана напівпроникна, утворює складки - кристи (гребінь, виріст), що оточені матриксом; на яких міститься багато ферментів. Кількість крист залежить від виду клітин (їх більше в активних клітинах). Між зовнішньою та внутрішньою мембранами вузький міжмембранний комплекс. Внутрішня порожнина мітохондрій — матрикс. В матриксі знаходяться зерна (діаметр 20-40 нм), в яких накопичуються Са2+ і Mg2+, а також полісахариди (глікоген). Також він містить 2-6 копій кільцевих молекул ДНК (без білків гістонів), рибосоми, т-РНК, ферменти реплікації ДНК, транскрипції і трансляції. За розмірами і структурою рибосом, організацією ДНК — цей апарат схожий з таким у прокаріот. Мітохондріальні ДНК включають гени синтезу мітохондріальних р-РНК, т-РНК, а також білків внутрішньої мембрани органели. Більшість мітохондрій закодовані в ядерних ДНК і утворюються в цитоплазмі. Розмножуються мітохондрії поділом. Всі мітохондрії синтезують АТФ; в них енергія хімічних зв’язків органічних речовин перетворюється на енергію фосфатних зв’язків АТФ (дихальні ферменти і ферменти синтезу АТФ лежать на внутрішній мембрані мітохондрій; АТФ використовується в усіх процесах, які потребують енергії).
Лізосоми — одномембранні органели, іноді ззовні вкритий волокнистим шаром білку, з високим вмістом гідролітичних ферментів. Вони містять комплекс ферментів для розщеплення жирів, вуглеводів та білків, беруть участь у виведенні органоїдів і частин клітини, які відмирають. Серед лізосом виділяють три типи: первинні лізосоми, вторинні лізосоми, залишкові тільця.
Первинні лізосоми — це дрібні мембранні міхурці, діаметром 100 нм, заповнені ферментами в неактивному стані.
Вторинні лізосоми — утворюються з первинних в результаті злиття з ендоцитозними вакуолями. В них відбуваються процеси травлення речовин, які надходять під час ендоцитозу, гідролітичними ферментами. Вторинні лізосоми поділяються на гетеролізосоми (фаголізосоми), аутолізосоми (цитолізосоми).
Фаголізосоми розщеплюють чужорідні речовини, які надходять у клітину шляхом фагоцитозу і піноцитозу.
Аутофагосоми розщеплюють власні компоненти клітини.
Залишкові тільця (телолізосоми) — це вторинні лізосоми, у яких процес переварювання завершений, в них знаходяться неперетравлені речовини. Залишкові тільця можуть бути або виділені з клітини (екзоцитоз), або залишитись у клітині до її загибелі.
Мікротільця — збірна група мембранних органел, це пухирці діаметром 0,1-1,5 мкм з дрібнозернистим матриксом і з білковими включеннями. До цих органел відносять, зокрема, пероксисоми. Вони містять ферменти оксидази, які каталізують дезамінування амінокислот і утворення пероксиду водню (Н2О2). Пероксид водню — токсичний продукт, руйнується ферментом пероксидазою (каталаза).
Пероксисоми зв’язані з мембранними ЕПС і утворюються на розширених ділянках цистерн комплекса Гольджи. В тварин і деяких рослин пероксисоми приймають участь у перетворенні жирів та вуглеводів.
Вакуолі — мембранні органели рослинних клітин, які виконують важливі функції та заповнені клітинним соком. В молодих клітинах вони дрібні, далі ростуть, зливаються утворюючи одну або декілька великих, які займають до 80% об’єму цитоплазми. Функції: регуляція осмотичного тиску у клітинах; накопичення поживних речовин; виконання специфічних функцій: травні вакуолі, скоротливі вакуолі.
Рибосоми — це округлі невеликі тільця діаметром 20-30 мм. Розташовуються в гіалоплазмі та на мембранах гранулярної ЕПС. Рибосоми складаються з малої та великої субодиниць, об’єднання яких відбувається у присутності інформаційної РНК. У кожній субодиниці знаходиться по молекулі р-РНК та білки.
Рибосоми, які об’єднуються з і-РНК нагадують низку намиста і таку структуру називають полісомою. Полісоми вільно розміщуються у цитоплазмі або прикріплюються до мембран гранулярною ЕПС. На полісомах гіалоплазми синтезуються білки, які використовуються для власних потреб, а на полісомах гранулярної ЕПС відбувається синтез білків, які секретуються клітиною (травні ферменти, муцин слини та ін.)
Клітинний центр (центріолі) — немембранні органели, за даними електронного мікроскопу мають вигляд порожнистого циліндра (шириною — 0,15 мкм, довжиною — 0,30,5 мкм). Стінки циліндра утворені 27 мікротрубочками, згрупованими у 9 триплетів. Біля центріолей знаходяться саталіти, фокуси сходження мікротрубочок, волокнисті корінці, додаткові мікротрубочки, які утворюють особливу зону — центросферу. Центріолі (2-3) і центросфера утворює клітинний центр. Клітинний центр змінює свою структуру під час клітинного циклу. Наприклад, з профази до телофази центріолі мають схожу будову. Під час мітозу утворюється 2 клітинних центра, в яких знаходиться по дві центріолі (материнська та дочірня, в якій материнська оточена широкою зоною тонких фібрил).
В кінці телофази, коли утворюються нові інтерфазні ядра порушується веретено поділу, материнська і дочірня центріолі відходять на незначну відстань та втрачають взаємоперпендикулярне розміщення.
Функції центріолей — утворення апарата ділення під час мітозу, забезпечення розходження хроматид (хромосом) в анафазі мітозу, а також формування та організація цитоплазматичних мікротрубочок (цитоскелету) під час інтерфази. Центріолі характерні для тваринних клітин, їх немає у вищих рослин, у нижчих грибів і деяких найпростіших. Утворені нитки мітотичного веретена складається також з мікротрубочок.
В цитоплазмі еукаріотичних клітин містяться 2 види структурних скоротливих елементів немембранної природи — мікротрубочки та мікрофіламенти.
Мікротрубочки — трубчасті утворення різної довжини (діаметром 24 нм, товщиною стінки 5 нм). Знаходиться у вільному стані у цитоплазмі клітин, а також є структурними елементами війок, джгутиків, ниток мітотичного веретена, центріолей.
Мікротрубочки збираються з субодиниць білкової природи шляхом їх полімеризації (при наявності Mg2+). В цитоплазмі вільні трубочки визначають форму клітин та направлений рух цитоплазматичних компонентів.
Мікрофіламенти — довгі і тонкі структури цитоплазми товщиною 7 нм, які можуть утворювати пучки. Особливо багато їх у кортикальному шарі цитоплазми. Існують актинові мікрофіламенти (містять скоротливий білок — актин), які забезпечують клітинні форми руху, наприклад, амебоїдний. Вони виконують також каркасну (опірну) функцію та визначають внутрішньоклітинний рух органел та гіалоплазми.
Під плазмолемою, а також у навколо ядерному просторі знаходяться пучки мікрофіламентів товщиною 10 нм — проміжні мікрофіламенти. В різних видах клітин вони побудовані з різних білків. Наприклад, в фібробластах знаходять лимептином, в епітеліальних — каратани, в м’язових — десмін. Система проміжних мікрофіламентів також динамічно рухлива як мікротрубочок та актинових філаментів. При діленні клітини вони розпадаються на дві підковоподібні структури, а після утворення дочірніх клітин знов оточують ядро. Тому, їх основна функція механічна, каркасна. Це доводить такий факт, що у багатьох епітеліальних клітин, особливо покривного епітелію, проміжні мікрофіламенти утворюють товсті пучки монофібрил, які додають клітинам пружність та жорсткість.
Пластиди - двомембранні органели клітин рослин та деяких тварин (джгутикових). У вищих рослин пластиди поділяють на три типи: хлоропласти, лейкопласти, хромопласти. Хлоропласти - забарвленні у зелений колір завдяки ферменту хлорофілу. Основна функція - фотосинтез. Лейкопласти - безбарвні, забезпечують синтез і гідроліз крохмалю та білків. Хромопласти - пластиди жовтого або червоного та ін.. коліру, що надають забарвлення пелюсткам, плодам, листам.
Клітинні включення — це непостійні структури, які то виникають то зникають у процесі життєдіяльності клітини. Вони містяться у цитоплазмі чи у клітинному соку вакуолей рослинних клітин у твердому або рідкому станах і можуть мати вигляд кристалів, зерен (гранул) чи краплин. Це насамперед запасні речовини. Крохмаль у вигляді зерен зберігається у цитоплазмі рослинних клітин. У тваринних клітинах запасається полісахарид глікоген (у вигляді зерен чи волоконець), різні ліпіди та білки (у яйцеклітині вони мають назву «жовток»).
З метою вивчення структури та функціонування клітини на сучасному етапі застосовують новітні методи дослідження.
1. Електронна мікроскопія - вивчення дрібних структур клітини за допомогою електронного мікроскопу.
2. Цитохімічні і цитоспектрофотометрічні методи - для визначення розташування та кількісного вмісту в клітині хімічних речовин.
3. Швидкісне центрифугування - для виділення і вивчення часток, що входять до складу цитоплазми.
4. Метод рентгеноструктурного аналізу - вивчають молекулярну структуру речовин у клітині, розміри і просторове розміщення молекул та атомів.
5. Метод ауторадіографії (мічених атомів) - досліджують біохімічні процеси в клітині.
6. Метод полімеразної ланцюгової реакції (ПЛР) - виявляють наявність або відсутність певних послідовностей ДНК.
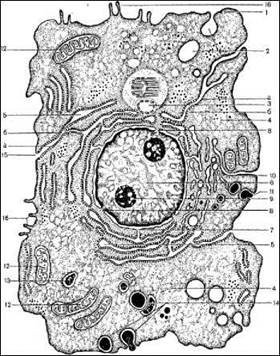
Рис. 2. Ультрамікроскопічна будова клітини (схема)
1 - зовнішня мембрана клітини (плазмалема); 2 - піноцитозні міхурці; 3 - центросома (клітинний центр); 4 - гіалоплазма; 5 - гранулярна ендоплазматичний ретикулум: а) α-цитомембрана; б) рибосоми; 6 - ядро; 7 - сполучення перинуклеарного простору з порожнинами, що утворені а цитомембранами; 8 - ядерні пори; 9 - ядерце; 10 - комплекс Гольджі; 11 - секреторні вакуолі; 12 - мітохондрії; 13 - лізосоми; 14 - стадії фагоцитозу; 15 - сполучення плазмалеми з а-цитомембранами; 16 - мікроворсинки.
Генетичні зв’язки клітинних мембран можна відобразити такою схемою: