ОСНОВИ МЕДИЧНОЇ БІОЛОГІЇ - 2012
Молекулярні основи спадковості. Характеристика нуклеїнових кислот. Трансформація і трансдукція
Трансформація (лат. transformatio - перетворення) - перенесення генетичної інформації від бактерії до бактерії за допомогою ізольованої ДНК. Явище трансформації вперше в 1928 р. відкрив англійський мікробіолог Ф. Гриффітс, який працював з двома штамами пневмококів (Streptococcus pneumonia).
Штам (S) має полісахаридну капсулу і відрізнявся високою вірулентністю (викликав пневмонію), інший (R) - безкапсульний, був авірулентний (не викликав захворювання). Здатність пневмококів-S спричинювати захворювання пов'язана з наявністю капсули, яка захищає їх в організмі від фагоцитозу. У серії дослідів на білих мишах Гриффітс одержав наступні результати:
✵ при введенні живого R-штаму миші виживали;
✵ при введенні живого S-штаму миші гинули від пневмонії;
✵ при введенні S-штаму, вбитого нагріванням, миші виживали;
✵ при введенні вбитого нагріванням S-штаму разом з живим R-штамом більшість мишей гинула, а з їхньої крові висівався живий S-штам.
Був зроблений висновок, що властивості вбитих бактерій - наявність капсули та вірулентність передалися живим бактеріям. Отже, відбулося перетворення (трансформація) авірулентного штама R у вірулентний S ( R - S).
Хімічну природу трансформуючого фактору встановили в 1944 р. американські генетики О. Евері, К. Мак-Леод і М. Мак-Карті. Ним виявилася ДНК.
Трансформація відбувається не в усіх клітин, а лише в тих, які здатні до неї. Такі клітини називаються компетентними. Механізм трансформації полягає в наступному: 1) дволанцюгова молекула ДНК бактерії-донора адсорбується на поверхні клітини-реципієнта; 2) поглинання клітиною-реципієнтом ДНК донора; 3) перетворення дволанцюгової ДНК донора в одноланцюгові фрагменти під дією нуклеаз клітини; 4) приєднання фрагмента одноланцюгової ДНК до хромосоми реципієнта; 5) експресія інтегрованого гена донора в клітині, внаслідок чого клітина набуває нових ознак, які успадковуються.
Трансдукція (лат. transductio - перенесення) - перенесення фрагментів ДНК від бактерії-донора до бактерії-реципієнта за допомогою помірних бактеріофагів. Помірні бактеріофаги, вивільнюючись з бактерії, у якій вони паразитували, захоплюють частину ДНК і переносять її у нову бактерію, передаючи властивості попередніх бактерій. Трансдукція надзвичайно поширена серед бактерій. Вона не тільки підтверджує генетичну роль ДНК, а використовується також для вивчення структури хромосом і генів, у генній інженерії.
Нуклеїнові кислоти: ДНК і РНК
Існують два типи нуклеїнових кислот (НК) - дезоксирибонуклеїнова (ДНК) і рибонуклеїнова (РНК). У клітинах прокаріотів і еукаріотів містяться обидві нуклеїнові кислоти (ДНК і РНК), віруси мають лише одну з них (ДНК або РНК). НК містяться в усіх живих організмах і виконують унікальні біологічні функції, пов'язані зі збереженням, реалізацією і передаванням спадкової інформації в ряді поколінь.
Дезоксирибонуклеїнова кислота (ДНК) - високомолекулярна органічна сполука, біополімер, мономерами якого є нуклеотиди, з'єднані між собою в довгий полінуклеотидний ланцюг. Кожен нуклеотид ДНК (дезоксиробонуклеотид) складається з трьох компонентів: 1) цукру пентози - дезоксирибози (звідси і назва цієї кислоти - дезоксирибонуклеїнова); 2) однієї з чотирьох азотистих основ, які належать до різних класів: аденін (А) і гуанін (Г) - до класу пуринових, цитозин (Ц) і тимін (Т) - до класу піримідинових основ; 3) залишку фосфорної кислоти.
Макромолекула ДНК - це два довгі полімерні ланцюги. У ланцюгу ДНК нуклеотиди сполучаються між собою за допомогою фосфодіефірних зв'язків, які утворюються між 3’- і 5’- гідроксильними групами дезоксирибоз сусідніх нуклеотидів. У полінуклеотидному ланцюзі ДНК виділяють два кінці. Один кінець несе гідроксильну групу (-ОН), яка зв'язана з 3’-вуглецем цукру дезоксирибози, а на іншому кінці знаходиться залишок фосфорної кислоти в 5’-положенні цукру. У складі нуклеотиду дезоксирибоза розташована посередині - між фосфорною кислотою і азотистою основою. Відносна молекулярна маса ДНК сягає 1500000 - 2000000 і більше.
ДНК характеризується певними закономірностями, встановленими в 1950 р. американським біохіміком Е. Чаргаффом (народився в м. Чернівці). Правила Чаргаффа:
1) сума пуринових основ дорівнює сумі піримідинових основ: А+Г=Т+Ц;
2) кількість гуаніну і тиміну дорівнює кількості аденіну і цитозину: Г+Т=А+Ц, або (Г+Т)/(А+Ц)= 1;
3) вміст аденіну дорівнює вмісту тиміну, а вміст гуаніну дорівнює вмісту цитозину (правило еквівалентності): А=Т, Г=Ц.
4) склад ДНК різних організмів може варіювати лише за відношенням (Г+Ц)/(А+Т), але для організмів даного виду це відношення є постійним: у людини воно дорівнює 1,54, у бактерії кишкової палички - 1,0.
5) Існують ДНК: АТ-типу, де А+Т>Г+Ц і ГЦ-типу, де Г+Ц>А+Т. АТ-тип ДНК характерний для вищих рослин, тварин і людини, а ГЦ-тип - для грибів бактерій вірусів.
Азотисті основи та нуклеотиди, які вони утворюють
Назва азотистої основи |
Позначення |
Назва нуклеотиду |
||
Аденін |
А |
dAp |
Дезоксиаденозин-*5’-фосфат* |
Аденозин-3’-фосфат |
Тимін |
Т |
dTp |
Тимідин-3’-фосфат |
- |
Урацил |
У |
Ур |
- |
Уридин-3’-фосфат |
Гуанін |
Г |
dGp |
Дезоксигуанозин-3’-фосфат |
Гуанодин-3’-фосфат |
Цитозин |
Ц |
dCp |
Дезоксицитидин-3’-фосфат |
Цитидин-3’-фосфат |
У 1953 р. американський біохімік Дж. Уотсон і англійський фізик Ф. Крік (Кембриджський університет) на основі даних хімічного і рентгеноструктурного аналізів запропонували модель просторової структури ДНК у вигляді подвійної спіралі (Нобелівська премія, 1962). За моделлю Уотсона-Кріка, молекула ДНК складається з двох полінуклеотидних ланцюгів, закручених вправо навколо спільної осі в подвійну спіраль. Два ланцюги в молекулі ДНК антипаралельні: один ланцюг має напрямок 5’>3’, інший - 3’>5’. Подвійна спіраль молекули ДНК утримується водневимизв’язками між основами протилежних ланцюгів за принципом комплементарності: аденін одного ланцюга завжди з’єднаний з тиміном іншого (А-Т), а гуанін - з цитозином (Г-Ц). Між аденіном і тиміном подвійний водневий зв’язок, між гуаніном і цитозином - потрійний. Спарені основи розташовані між двома ланцюгами перпендикулярно загальній осі. Вся структура нагадує гвинтову драбину, бічні частини якої утворені цукрофосфатним каркасом, а східці - спареними основами. Діаметр спіралі - 2 нм, відстань між азотистими основами впродовж осі спіралі - 0,34 нм, десять пар нуклеотидів утворюють один повний виток подвійної спіралі довжиною 3,4 нм.
Відомо декілька форм ДНК: 1) В-форма у нормальних фізіологічних умовах є основним структурним типом (структура за моделлю Уотсона-Кріка); 2) А-форма - виявлена у зневодненому середовищі (ДНК ширше (2 нм) і один виток спіралі включає 11 пар нуклеотидів); 3) С-форма має менше основ на один виток; 4) Z-форма - закручена вліво.
Рибонуклеїнові кислоти (РНК) високомолекулярні органічна сполуки, які представлені одним полінуклеотид ним ланцюгом. РНК - це біополімер, мономерами якого є рибонуклеотиди. Нуклеотид РНК, як і нуклеотид ДНК, складається з трьох компонентів: 1) цукру пентози - рибози (звідси і назва цієї кислоти - рибонуклеїнова); 2) однієї з чотирьох азотистих основ: аденін (А) і гуанін (Г) належать до класу пуринових, цитозин (Ц) і урацил (У) - до класу піримідинових основ; 3) залишку фосфорної кислоти.
Одноланцюгова РНК інколи може утворювати подвійну структуру, якщо різні частини ланцюга мають антипаралельні комплементарні фрагменти, які зв’язуються водневими зв’язками (А-У, Г-Ц). У деяких вірусів РНК виступає носієм спадкової інформації замість ДНК. В клітинах зустрічаються три основних види РНК, які виконують різноманітні функції: інформаційні або матричні РНК (і-РНК, м-РНК), транспортні РНК (т-РНК), рибосомальні РНК (р-РНК). Всі три види РНК синтезуються на матрицях-молекулах ДНК за допомогою ферментів РНК-полімераз за принципом комплементарності. Усі типи РНК синтезуються у вигляді РНК-попередниць, які після транскрипції зазнають певних змін, перетворюючись у біологічно активні РНК. Цей процес називається дозріванням, або процесингом.
Рибосомальна РНК (рРНК) - це велика одноланцюгова розгалужена молекула, яка має найбільшу довжину, містить до 3-5 тис. нуклеотидів та складає до 90 % усієї РНК клітини. рРНК утворюється в ядрі клітини у ядерці на ядерцевих організаторах хромосом. Оскільки рибосом у клітинах прокаріотів і еукаріотів багато, то гени, що кодують синтез рРНК, представлені багатьма копіями. У каріоплазмі рРНК разом з рибосомними білками утворюють мали та великі субодиниці рибосом. рРНК утворює структурний каркас рибосом для функціонування білоксинтезуючої системи клітини.
Транспортна РНК (тРНК) - найкоротша за розмірами, однониткова, має форму листка конюшини, містить 70-90 нуклеотидів та складає 10-20 % загальної клітинної РНК, на відміну від інших РНК, розчинна. Транспортні РНК транспортують активовані амінокислоти до місця біосинтезу білків (рибосом) для включення їх в поліпептидний ланцюг, що тут синтезується. Для кожної амінокислоти існує специфічна тРНК. Оскільки більшість амінокислот кодується кількома кодонами, кількість різних типів РНК перевищує кількість амінокислот (20). Молекула тРНК має чотири ділянки: транспортну, антикодон, ділянку приєднання фермента, ділянку зв’язування з рибосомою.
Загальна властивість усіх типів тРНК - на 3'-кінці (акцепторному) знаходиться триплет нуклеотидів ЦЦА. Кінцевий аденозин антикодона через 3 -ОН групу рибози акцептує (приєднує) активовану амінокислоту в процесі трансляції. тРНК слугують проміжними молекулами між іРНК і поліпептидом.
Інформаційна, або матрична, РНК (іРНК, мРНК) складає 0,5-3 % загальної кількості клітинної РНК. Довжина її може бути різною - від 300 до 3000 нуклеотидів, що залежить від довжини гена. Молекули іРНК утворюються на певних ділянках ДНК - структурних генах. Вони переносять генетичну інформацію про первинну структуру білка з ядра від ДНК у цитоплазму до рибосом. Інформаційну РНК називають матричною РНК (мРНК), тому що вона є матрицею (шаблоном), на якій будується поліпептид відповідно до інформації, яку вона містить. іРНК, яка несе інформацію про синтез однієї молекули білка, називається моноцисторонна. Якщо у молекулі іРНК декілька цитронів для різних білків - поліцистронна.
Інформаційна РНК утворюється у вигляді незрілої про-іРНК, що містить неінформативні ділянки - інтрони, що вирізаються під час процесінгу (дозрівання іРНК), та інформативні ділянки - екзони. Зріла іРНК виходить у цитоплазму, з’єднується з рибосомою і реалізує свою інформацію. Проте інколи іРНК може накопичуватися у клітині, зв’язуватися зі специфічними білками, які «консервують» її, і утворювати інформосоми. Фізіологічні зміни, які призводять до активації синтезу білка, слугують поштовхом для використовуванні РНК інформосом.
ДНК виконує такі функції: хімічна основа хромосомного генетичного матеріалу (гена); бере участь у синтезі РНК та ДНК; несе інформацію про структуру білків. Біологічна роль ДНК полягає у зберіганні, реалізації генетичної інформації протягом життя клітини, передачі спадкової інформації від материнських до дочірних клітин, від батьків до нащадків.
РНК виконують різні функції:
- іРНК, або мРНК переносить інформацію про структуру білка від ДНК до місця синтезу білка в рибосоми;
- тРНК переносить амінокислоти до місця синтезу білка, в рибосоми;
- рРНК входить до складу рибосом, формує їх функціональний скелет.
Реплікація ДНК (саморепродукція на молекулярному рівні) — складний біологічний процес реакції матричного синтезу, який забезпечує подвоєння молекули ДНК перед поділом клітини. Цей процес відбувається в ядрі клітини під час S-періоду інтерфази. Здатність до самоподвоєння - унікальна властивість ДНК, яка притаманна лише ДНК і непритаманна ніякій іншій хімічній речовині. Реплікація ДНК - найважливіший молекулярний процес, що лежить в основі всіх типів розмноження, а значить в основі забезпечення тривалого існування окремих організмів, популяцій і всіх видів живих організмів (підтримування сталості генотипів і фенотипів). Відбувається за напівконсервативним механізмом та за принципом комплементарності. Реплікація — ферментативний, енергозалежний процес.
Модель ДНК, запропонована Уотсоном та Криком дозволила зрозуміти принцип подвоєння молекули ДНК (реплікації ДНК). Біспіраль материнської молекули ДНК розділяється на два ланцюги і кожен з них слугує матрицею для синтезу за принципом комплементарності нового ланцюга. У результаті з однієї материнської молекули ДНК утворюються дві дочірні, які є точними копіями материнської. Оскільки в кожній новоутвореній дочірній молекулі один ланцюг старий (материнський), а інший - новий, такий механізм реплікації одержав назву напівконсервативного.
Синтез ДНК каталізує фермент ДНК-полімераза. Фермент ДНК-полімеразу вперше в 1957 р. виділив з бактерії кишкової палички (Escherichia coli) А.Корнберг. Існує декілька типів ДНК-полімераз. Процес реплікації ДНК забезпечується координованою роботою ряду білків та ферментів. Ці білки визначають відповідні гени ДНК, а це означає, що сама ДНК і визначає реплікацію. Інціаторні білки утворюють реплікаційну вилку. ДНК-топоізомерази розкручують ланцюги, ДНК-геліказа і дестабілізуючий білок розщеплюють ДНК на два окремих ланцюги, ДНК-полімераза та ДНК-праймаза каналізують полімеризацію нуклеотидтрифосфатів і утворення нового ланцюга, ДНК-лігази руйнують РНК-затравки та з'єднують між собою в один ланцюг короткі ділянки новосинтезованого полінуклеотидного ланцюга.
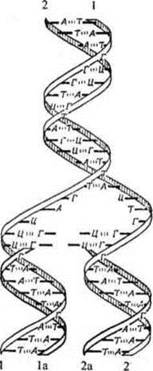
Рис. 7. Реплікація ДНК за схемою Дж. Уотсона і Ф. Кріка:
1, 2 - материнські ланцюги ДНК; 1a, 2а - дочірні ланцюги ДНК
Реплікація включає такі етапи:
I - Ініціація (лат. Initialis - початковий).
II - Елонгація ( подовження).
III - Термінація (лат. Terminalis - кінцевий).
Ініціація починається з визначення ферментами точки реплікації - точці огі (англ. origin - початок), де відбувається розкручування біспіралі ДНК, локальне розведення її ланцюгів, стабілізація однониткових ДНК, поява з двох розведених ланцюгів Y-подібної структури (реплікативної вилки). У вірусів прокаріот одна точка реплікації і ДНК являє собою одну одиницю реплікації — реплікон. У еукаріот молекули ДНК поділяються на декілька репліконів, а це означає, що мають декілька точок реплікації.
Відбувається активація дезоксирибонуклеотидів, які є «сировиною» для синтезу ДНК. Монофосфати дезоксирибонуклеотидів (дАМФ, дГМФ, дЦМФ, дТМФ) активуються при взаємодії з АТФ, утворюючи трифосфати (дАТФ, дГТФ, дЦТФ, дТТФ). Ця реакція називається фосфорилуванням і каталізується ферментом фосфорилазою.
Далі ферменти ДНК-гелікази руйнують водневі зв’язки між комплементарними азотистими основами утворюючи реплікаційне око. Ділянка, де утворюється реплікаційне око називається точкою реплікації. Вона містить близько 300 нуклеотидів і визнається специфічними білками.
Ферменти ДНК — топоізомерази (гірази) стабілізують молекулу ДНК вище точки реплікації попереджуючи суперспіралізацію шляхом розриву одного ланцюга в ДНК, який рухається навколо другого. Дестабілізуючі білки - SSB - білки приєднуються до вільних ланцюгів ДНК, що не сприяє спонтанному об’єднанню двох ланцюгів — утворюється реплікаційна вилка.
Елонгація. Вільні три фосфати дезоксирібонуклеотидів своїми азотистими основами приєднуються водневими зв’язками до азотистих основ обох ланцюгів ДНК по принципу компліментарності: А-Т, Г-Ц.
Фермент ДНК-полімераза починає приєднувати комплементарні активовані нуклеотиди (дезоксирибонуклеозидтрифосфати: дАТФ, дГТФ, дЦТФ, дТТФ) до одного з ланцюгів з вільним 3' кінцем. Так, поступово безперервно відбувається комплементарний синтез дочірнього ланцюга на одному ланцюгу ДНК, який має напрям зв’язків 5' 3' (від вуглецевого 5'-кінця до вуглецевого 3'-кінця молекул ДНК. Інший ланцюг антипаралельний, направлення зв’язків — 3' 5'. Дія фермента ДНК-полімерази в такому напрямку неможлива. У зв’язку з цим процес подвоєння в реплікаційній вилці є асиметричним. На одному ланцюзі він відбувається послідовно, безперервно та швидко. Цей ланцюг називають лідуючим (провідним). Синтез на другому ланцюзі відбувається повільно, тому такий ланцюг називають відстаючим.
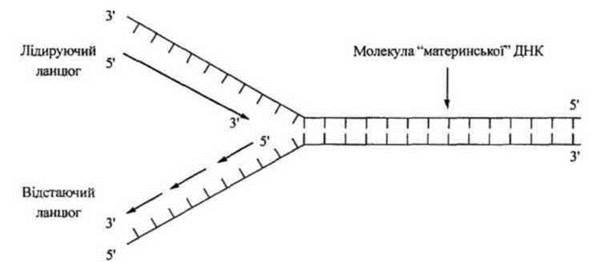
Рис. 8. Реплікація ДНК за схемою Р. Оказакі.
На відстаючому ланцюгу синтез дочірнього ланцюга відбувається преривчасто із запізненням. Спочатку утворюються короткі ланцюги РНК за шаблоном ДНК - РНК-праймери (10-60 нуклеотидів). Утворення праймерів каталізується РНК-праймазою. Праймери пізніше віддаляються, а порожнини, які утворилися, заповнюються дезоксирибонуклеотидами ДНК. ДНК-полімераза синтезує фрагменти дочірнього ланцюга ДНК в напрямку від 5' до 3' кінця. Тобто на місті праймера утворюються короткі фрагменти ДНК, які називаються фрагментами Оказаки. У прокаріот такі фрагменти містять 1000...2000 нуклеотидів, у еукаріот — 100...200 нуклеотидів. Далі ці фрагменти зшиваються (сплайсінг) ДНК-лігазами, праймери руйнуються і ДНК-полімераза добудовує комплементарні нуклеотиди ДНК.
Результатом синтезу є утворення 2-х ідентичних молекул ДНК, кожна з яких складається з одного материнського та другого дочірнього ланцюга.
Чітка компліментарність пар основ забезпечує точну реплікацію ДНК. Але іноді виникають помилки у приєднанні основ. Вони виправляються ДНК-полімеразою, яка для цього знову з’єднується з молекулами ДНК (репарація).
Термінація — завершення процесу реплікації. Утворені молекули ДНК спіралізуються, утворюючи хромосому, яка складається з двох хроматид.
Швидкість реплікації дуже висока (у прокаріот — 1000 нуклеотидів/сек; у еукаріотів - 100 нуклеотидів/сек). Ділянка ДНК, яка знаходиться під контролем однієї точки огі, називається одиницею реплікації (репліконом). У ДНК прокаріотів - одна точка огі і вся молекула подвоюється як один реплікон. У ДНК еукаріотів - багато точок огі і відповідно багато репліконів, що є необхідною умовою для того, щоб велетенські молекули ДНК еукаріотів встигли подвоїтися за період одного клітинного циклу.
Репарація ДНК
Клітини в процесі еволюції виробили певні механізми, які забезпечують стабільність спадкового матеріалу (ДНК), оберігають його від мутацій. Якби не було цих механізмів, організми загинули б від дії природних мутагенів, тому що частота генетичних пошкоджень ДНК висока. Більшість пошкоджень ДНК, які є потенціальним джерелом мутацій, усувається за допомогою кількох механізмів:
- висока хімічна стабільність самої молекули ДНК
- наявність спеціальних механізмів самокорекції і репарації виникаючих змін.
Репарація (лат. reparatio - відновлення) - процес відновлення первинної структури ДНК
після пошкодження її мутагенами. За механізмами розвитку розрізняють світлову (неексцизійну), темнову (ексцизійну) та рекомбінативну види репарації ДНК. Усі вони мають ферментативну природу, усувають лише однониткові пошкодження ДНК і контролюються спеціальними генами.
Світлова репарація (фоторепарація) відбувається за допомогою енергії видимого світла і усуває пошкодження, спричинені лише ультрафіолетовими променями. Цими пошкодженнями є димери - зчеплені між собою сусідні піримідинові основи того самого ланцюга ДНК. Димери виникають між двома тимінами, тиміном та цитозином, двома цитозинами, тиміном та урацилом, цитозином та урацилом, двома урацилами. Механізм світлової репарації полягає в наступному: фермент ДНК-фотолігаза специфічно зв'язується з пошкодженою УФ-опроміненням ділянкою ДНК, активується квантами денного світла і відщеплюється; відщеплюючись, фермент одночасно роз'єднує димери; нормальна структура ДНК відновлюється. Найбільш ефективним для світлової репарації є світло в блакитній частині спектра.
Темнова (ексцизійна, вирізаюча) репарація, на відміну від фоторепарації: не потребує енергії видимого світла; усуває пошкодження, зумовлені не лише ультрафіолетовими променями, але й іншими мутагенними факторами; здійснюється шляхом вирізання пошкодженого фрагмента нитки ДНК (ексцизіо - вирізати). Розрізняють наступні етапи темнової репарації: 1) впізнання пошкодженої ділянки ДНК ендонуклеазою; 2) надрізування ендонуклеазою ланцюга ДНК близько пошкодження; 3) вирізання пошкодженої ділянки ДНК з розширенням пролому в обидва боки екзонуклеазою; 4) матричний синтез фрагмента ДНК ("латки") на місці пролому (репаративна реплікація); 5) зшивання новоутвореної ділянки з основною ниткою ДНК за допомогою фермента лігази.
Постреплікативна репарація відбувається шляхом рекомбінації між двома утвореними подвійними спіралями ДНК. Приклад такої репарації — відновлення структури ДНК при виникненні тимінових димерів (Т-Т), коли вони не зникають під впливом світла (світова репарація) або під час дореплікативної ексцизійної репарації.
Світлова і темнова репарації усувають пошкодження ДНК до її реплікації (дореплікативна репарація). У випадку, якщо ці механізми не спрацювали, пошкодження ліквідуються після реплікації в дочірніх ланцюгах ДНК (постреплікативна репарація). Реплікативна репарація - сукупність процесів відновленні ДНК у ході реплікації.
При пошкодженні тих чи інших шляхів репарації виникають спадкові хвороби: пігментна ксеродерма, синдром Блюма, анемія Фанконі, юнацька прогерія (передчасне старіння).
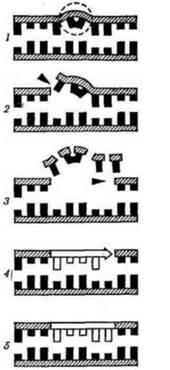
Рис. 9. Схема ексцизійної репарації ДНК:
1 - індуковане ультрафіолетовим світлом пошкодження молекули ДНК; 2 - розрізування одного ланцюга молекули ДНК дією ендонуклеаз; 3 - вирізування пошкодженої ділянки за допомогою екзонуклеаз; 4 - синтез нового ланцюга в результаті дії полімераз; 5 - зшивання новоутвореної ділянки за допомогою лігаз.
Пігментна ксеродерма - спадкова хвороба, яка пов'язана з порушенням темнової репарації ДНК. Проявляється вже в новонароджених світлобоязню.
Характерна підвищена чутливість шкіри хворих до ультрафіолетових променів. Це спричинює атрофію відкритих ділянок шкіри, гіперпігментацію, може розвинутися рак шкіри. Виділяють кілька форм пігментної ксеродерми. Тип успадкування - автосомно-домінантний і автосомно-рецесивний.
Принцип комплементарності — має на увазі встановлення водневих зв’язків між суворо визначеними азотистими основами двох ланцюгів: А=Т; Г=Ц. Послідовність нуклеотидів в одному ланцюзі молекули ДНК відповідає послідовності нуклеотидів в другому ланцюзі.
Принцип антипаралельності — має на увазі, що два полінуклеотидних ланцюга з’єднуються так, що 5'-й кінець одного з них з’єднується з 3'-ім кінцем другого ланцюга і навпаки. Один ланцюг спрямовується зверху до низу від 5'-ої до 3'-ої вуглеводневим атомом, а другий — з низу вверх.
![]()