МЕДИЧНА БІОЛОГІЯ, АНАТОМІЯ, ФІЗІОЛОГІЯ ТА ПАТОЛОГІЯ ЛЮДИНИ - Я.І.Федонюк 2010
БІОЛОГІЯ
РОЗДІЛ 1. БІОЛОГІЧНІ ОСНОВИ ЖИТТЄДІЯЛЬНОСТІ ЛЮДИНИ
1.3. МОЛЕКУЛЯРНО-ГЕНЕТИЧНИЙ І КЛІТИННИЙ РІВНІ ОРГАНІЗАЦІЇ ЖИТТЯ
1.3.2. Структурно-хімічна функціональна організація еукаріотичних клітин
Морфофункціональна характеристика та класифікація хромосом людини
Хромосоми - структури ядра клітини, в яких знаходяться гени (ДНК). Це матеріальні носії спадковості. Хромосоми разом з позахромосомними структурами забезпечують збереження, відтворення, реалізацію і передачу генетичної інформації в ряді поколінь. Хромосоми здатні до самовідтворення. Перед кожним клітинним поділом вони подвоюються, а потім точно порівну розподіляються між дочірніми клітинами. Назву їм дав німецький гістолог В.Вальдейєр (1888) за здатність інтенсивно забарвлюватися основними барвниками (грец. chroma- колір, soma-тіло).
Головні хімічні компоненти хромосом -ДНК, основні (гістонові) і кислі (негістонові) білки, частки яких складають відповідно 40 %, 40 % і близько 20 %. ДНК знаходиться не у вільному стані, а в комплексі з білками. Білки по всій довжині хромосоми - гістони. У хромосомах спермієв деяких видів (лосось, оселедець) ДНК зв'язана не з гістонами, а з протамінами. Гістонові білки представлені п'ятьма головними фракціями (НІ, Н2А, Н2В, НЗ, Н4) і виконують структурну і регуляторну функції. Число фракцій негістонових білків перевищує 100. Серед них ферменти синтезу і процесингу РНК, редуплікації і репарації ДНК.
Хромосоми можуть знаходитися в двох структурно-функціональних станах: конденсованому (спіралізованому) і неконденсованому (деспіралізованому), які можуть взаємно переходити один в одний. При цьому лінійні розміри (довжина, товщина) хромосом змінюються. Спіралізований стан властивий хромосомам під час мітозу (мітотичні хромосоми), деспіралізований - між поділами (інтерфазні хромосоми). Мітотичні хромосоми видно в світловий мікроскоп у вигляді ниток, паличок, які добре забарвлюються основними барвниками. Функція мітотичних хромосом - транспортна: точний розподіл і перенесення генетичного матеріалу в дочірні ядра. Інтерфазні хромосоми максимально деспіралізовані, індивідуально не розрізняються і займають весь об'єм ядра, утворюючи хроматин. Функція інтерфазних хромосом - синтетична: синтез ДНК (реплікація) і синтез РНК (транскрипція).
Будову хромосом вивчають у метафазі, коли вони найбільш конденсовані і їх добре видно в світловий мікроскоп. Метафазна хромосома складається з двох сестринських хроматид, які розташовані поруч і з'єднані між собою в ділянці первинної перетяжки (центромери). Центромера - звужена, незабарвлена і найменш спіралізована ділянка хромосоми. Вона містить кінетохор (грец. kinesis - рух, phoros - несучий), до якого прикріплюються мікротрубочки веретена поділу. Центромера поділяє хромосому на два плеча. Кінці плеч називаються теломерами. Це ділянки із щільно упакованою ДНК, які перешкоджають з'єднанню хромосом між собою або з їхніми фрагментами. Позбавлений теломери кінець хромосоми стає "липким" і легко приєднує фрагменти хромосом.
За положенням центромери розрізняють метацентричні, субметацентричні та акроцентричні хромосоми (рис. 1.34).

Рис. 1.34. Типи хромосом людини в залежності від положення центромери (стадія метафази):
1 -акроцентрична; 2 - акроцентрична із супутником; 3 - субметацентрична; 4 - метацентрична.
У метацентричних хромосом центромера знаходиться посередині хромосоми і плечі рівної або майже рівної довжини (рівноплечі хромосоми). Субметацентричні - нерівноплечі хромосоми, в яких центромера зміщена від середини і одне плече довге, а друге коротке. Акроцентричні - паличкоподібні хромосоми, в яких центромера розміщена близько до одного з кінців і одне плече довге, а друге дуже коротке і часто малопомітне. Можуть виникати і телоцентричиі хромосоми в результаті втрати одного плеча, у них лишається тільки одне плече з центромерою на кінці. У нормі такі хромосоми не зустрічаються. Деякі хромосоми мають вторинні перетяжки, які знаходяться в прицентромерних районах довгих плечей (хромосоми 1,9, 16) і на кінцевих ділянках коротких плеч (хромосоми 13-15 і 21, 22). Вторинні перетяжки хромосом 13-15 і 21, 22 відокремлюють тонкою ниткою від основної частини невеликі кінцеві ділянки, які називаються супутниками, а самі хромосоми - супутниковими. У зоні вторинних перетяжок деяких хромосом розташовані ядерцеві організатори - місце утворення ядерець.
Ультраструктурна організація хромосом. Однохроматидна хромосома містить одну суперспіралізовану молекулу ДНК. Рівнів спіралізації існує декілька. Елементарною структурою хромосоми, яку розрізняють в електронному мікроскопі, є нитка діаметром 10-13 нм (1нм=10~3 мкм=10~ 6мм), яка являє собою комплекс ДНК і гістонових білків (нуклеогістон). По її довжині, як намистини на нитці, розташовані нуклеосоми. Нуклеосома має серцевину, або ядро, яке складається з 8 молекул гістонів чотирьох класів (гістоновий октамер, гістоновий кор) по дві молекули кожного класу Н2А, Н2В,НЗ і Н4 (рис. 1.35). Біспіраль ДНК (146 пар нуклеотидів) робить 1,75 витків на ній і переходить на серцевину наступної нуклеосоми. Міжнуклеосомні ділянки ДНК (лінкери) з'єднуються гістоном НІ. Закручування молекули ДНК на гістонові тільця зменшує довжину її біспіралі приблизно в 7 разів. На наступному рівні спіралізації виникає нитка діаметром 20-25 нм. Загальна довжина ДНК на цьому рівні зменшується в 40 разів. Шляхом наступної суперспіралізації з утворенням петель і багаторазового поздовжнього складування формуються мітотичні хромосоми. Пучки фібрил утворюють хромонеми. Вздовж хромонем розміщені ділянки (вузлики) більш щільної компактизації ДНК - хромомери. Завдяки суперспіралізаціі досягається щільна упаковка генетичного матеріалу, що дуже важливо при переміщеннях хромосом у мітозі. Про щільність упаковки свідчать наступні цифри. Довжина найбільшої, першої, хромосоми людини - 11 мкм, а довжина її молекули ДНК в розгорнутому вигляді - близько 7 см. У надкомпактному стані хромосоми неактивні: ферментам або білкам - регуляторам тяжко підійти до активних груп ДНК.
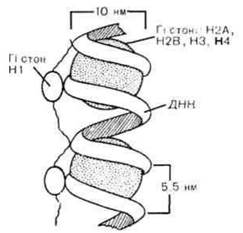
Рис. 1.35. Нуклеосомна організація елементарної нитки хромосоми.
Необхідною умовою експресії будь-якого району хромосом є його деконденсація.