МЕДИЧНА БІОЛОГІЯ, АНАТОМІЯ, ФІЗІОЛОГІЯ ТА ПАТОЛОГІЯ ЛЮДИНИ - Я.І.Федонюк 2010
БІОЛОГІЯ
РОЗДІЛ 1. БІОЛОГІЧНІ ОСНОВИ ЖИТТЄДІЯЛЬНОСТІ ЛЮДИНИ
1.3. МОЛЕКУЛЯРНО-ГЕНЕТИЧНИЙ І КЛІТИННИЙ РІВНІ ОРГАНІЗАЦІЇ ЖИТТЯ
1.3.3. Спадковий апарат еукаріотичних клітин і його функціонування - молекулярний рівень
Регуляція експресії генів у прокаріотів і еукаріотів
Усі соматичні клітини організму містять однаковий набір хромосом, однаковий набір генів і несуть повну генетичну інформацію, властиву даному виду. Проте клітини в одного і того самого організму різні за будовою, функцією, складом білків-ферментів. Основу цих відмінностей складає експресія (активність) різних частин генома. У клітині існують дві групи генів. Одні з них (конститутивні гени) експресуються постійно, тому що продукти, синтез яких вони кодують, клітині необхідні постійно (рибосомні білки, РНК-полімерази, молекули тРНК, рРНК, ін.). Інші гени "працюють" лише за певних умов, тоді, коли клітині необхідні ферменти, синтез яких вони кодують. Експресія цих генів регулюється генетичними і негенетичними факторами. Механізм регуляції експресії генів вперше в 1961 р. пояснили французькі мікробіологи Ф.Жакоб і Ж.Моно, які вивчали регуляцію синтезу ферментів у бактерії кишкової палички (Е.соїі). Цей механізм відомий як гіпотеза оперона (Нобелівська премія, 1968).
Оперон - генетична одиниця регуляції транскрипції генів у прокаріотів. Це послідовність нуклеотидів ДНК, яка об'єднує структурні гени, промотор, оператор і термінатор. Структурні гени тісно зчеплені між собою, розташовані один біля одного і несуть інформацію для синтезу ферментів
послідовних реакцій одного метаболічного циклу. Функціонують структурні гени одного оперону як одне ціле. До контрольних ділянок оперона належать промотор, оператор і термінатор. Промотор - ділянка ДНК оперона, до якої первинно прикріплюється РНК-полімераза і з якої починається транскрипція структурних генів. Оператор - ділянка ДНК оперона, яка розташована між промотором і структурними генами і з якою може зв'язуватися білок-репресор. Структурні гени синтезують матричну РНК, коли оператор включений, і припиняють синтез, коли оператор виключений. Оператор включається або виключається білком-репресором. Синтез білка-репресора кодується геном- регулятором, який може розміщуватися поруч з опероном або на відстані від нього. Білок-репресор може бути в двох формах- активній і неактивній. Перебуваючи в активній формі, репресор зв'язується з оператором і виключає його. Це виключає із функції весь оперон (транскрипція не відбувається). Включення оперона відбувається в тому випадку, коли білок-репресор знаходиться в неактивній формі і не здатний зв'язатися з оператором. У регуляції експресії генів беруть участь негенетичні фактори - ефектори. Ефектори - низькомолекулярні речовини, які взаємодіють з білками-репресорами і змінюють їх здатність зв'язуватися з оператором. Якщо комплекс (оператор+білок-регулятор) виключає транскрипцію, то такий контроль називається негативним, якщо включає - позитивним. Оперони працюють за типом індукції і за типом репресії.
Класичним прикладом роботи оперона за типом індукції є модель лактозного оперона (Іас- оперона) бактерії кишкової палички. Lac-оперон містить три структурних гена, які кодують синтез трьох ферментів, необхідних для метаболізму лактози (рис. 1.47). Робота оперона знаходиться під контролем гена-регулятора, який кодує синтез активного білка-репресора. За умови відсутності лактози білок-репресор зв'язується з оператором, транскрипція не відбувається (негативний контроль). Якщо в культуральному середовищі з'являється лактоза (замість глюкози), вона проникає в клітину і зв'язується з білком-репресором, він стає неактивним і не може зв'язатися з оператором. Оператор звільняється від білка-репресора і структурні гени можуть працювати (транскрипція відбувається). Поки в середовищі знаходиться лактоза, до тих пір працює оперон і утворються ферменти, необхідні для її розщеплення. Таким чином, лактоза виступає одночасно як субстрат розщеплення і як індуктор свого власного розщеплення. Негативний контроль роботи lac-оперона пояснює сутність феномену індукції: немає індуктора (лактози) - оперон не працює, є індуктор(лактоза) - оперон активно працює.
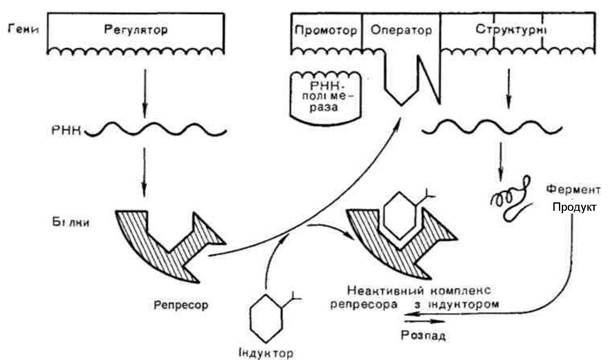
Рис. 1.47. Схема регуляції транскрипції структурних генів прокаріотичної клітини за типом індукції.
Феномен репресії ферментів пояснює модель роботи триптофанового оперона.
Триптофан, а, отже, і фермент для його синтезу (тригпофансиніеіаза) потрібні клітині в нормальних умовах завжди.
Триптофановий оперон переважно включений і транскрипція структурних генів відбувається за рахунок того, що ген-регулятор кодує синтез неактивного білка- репресора, який не може приєднатися до оператора і виключити його. Якщо в клітині з'являється надлишок триптофану (виникає потреба позбутися його), деяка кількість триптофану виступає як корепресор. Корепресор зв'язується з неактивним репресором і активує його. Комплекс (репресор+корепресор) приєднується до оператора і виключає його, що виключає весь оперон і синтез триптофансинтетази припиняється (негативний контроль).
Особливістю триптофанового оперона є наявність у ньому лідерної послідовності нуклеотидів ДНК (аттенуатора), яка розташована між промоторно-операторною ділянкою і першим структурним геном. Сутність феномену аттенуації полягає в тому, що навіть при незначному надлишку триптофана транскрипція обривається в ділянці аттенуатора РНК-полімеразою. Аттенуатори, таким чином, здійснюють більш економну регуляцію роботи оперона. Триптофанів оперон працює за типом зворотного зв'язку.
Описані системи регуляції структурних генів носять пристосувальний характер. У першому випадку синтез ферментів запускається надходженням у клітину субстрата розщеплення, в іншому- утворення фермента припиняється, як тільки зникає потреба в синтезі певної речовини. Регуляція біосинтезу білків здійснюється, крім транскрипції, на етапі трансляції, посттрансляційної модифікації, формування ознак організму.
Контроль експресії генів в еукаріотів значно складніший і ще мало вивчений. В еукаріотів відсутні оперони, подібні до оперонів бактерій. Гени, які кодують послідовні реакції біосинтезу, можуть знаходитися в різних ділянках однієї хромосоми або навіть у різних хромосомах. Первинний транскрипт (про-іРНК) зазнає посттранскрипційної модифікації (процесинг). Геном вищих еукаріотів значно складніший (у прокаріотів - одна хромосома, у людини - 46). У клітинах еукаріотів ядерна оболонка просторово розділяє процеси транскрипції (в ядрі) і трансляції (у цитоплазмі). На експресію еукаріотичних генів впливає ампліфікація генів - багатократне збільшення числа копій однакових генів. На відміну від прокаріотів гени еукаріотів мають інтрони. Ще одна особливість генома еукаріотів - наявність спеціальних "підсилюючих" сегментів ДНК - енхансерів. Активність генів в еукаріотів регулюється ендокринною системою. Багато гормонів є генетичними індукторами, тобто володіють здатністю включати і виключати різні гени.