Фізіологія людини - Вільям Ф. Ґанонґ 2002
Фізіологія нервових і м'язових клітин
Збудлива тканина: нервова
Нервові клітини
Морфологія
Нейрони центральної нервової системи ссавців мають різноманітні форми і розміри (рис. 2-1). Проте більшість із них складається з тих самих частин, що й типові спинномозкові рухові нейрони (рис. 2-2). Ці клітини мають від п’яти до семи відростків, які інтенсивно розгалужуються від тіла клітини; їх називають дендритами. Зокрема, в мозковій корі та корі мозочка дендрити формують невеликі шишкоподібні виступи-дендритні шипики. Типовий нейрон теж має довгий волокнистий відросток - аксон, який починається з дещо потовщеної ділянки тіла клітини - горбика аксона. Початкову частину аксона називають початковим сегментом. Ближче до закінчення аксон розгалужується на аксонні терміналі, кожна з яких закінчується численними синаптичними потовщеннями, які називають кінцевими ґудзиками, або аксонними телодендронами. Вони містять гранули або пухирці, у яких накопичуються синтезовані в нервах синаптичні трансмітери (див. Розділ 4).
Аксони багатьох нейронів мієлінізовані, тобто вкриті оболонкою мієліну, білково-ліпідного комплексу, утвореного багатьма шарами клітинних мембран - клітинами Шванна (нейролемоцитів) (рис. 2-3). Клітини Шванна - це гліоподібні клітини, розміщені вздовж периферійних нервів. Мієлінова оболонка утворюється внаслідок багаторазового (до 100) обгортання аксона мембраною клітини Шванна. Мієлін ущільнюється, якщо позаклітинні частини мембранного білка, що називають білком нуль (Р0-від англ. protein), змикаються з позаклітинними частинами прилеглої мембрани. Різні мутації в гені Р0 призводять до периферійних нейропатій; описано 29 різних мутацій, що є причиною як помірних, так і важких порушень. Мієлінової оболонки нема в ділянці термінальних розгалужень аксона і в періодичних (приблизно через кожен 1 мм розміром до 1 мкм) ділянках звужень, які називають вузловими перетяжками Ранв’є. Ізолювальну функцію мієліну описано нижче. Однак не всі нейрони в організмі ссавців укриті мієліном, деякі з них безмієлінові, тобто просто оточені клітинами Шванна без багаторазового обгортання аксона мембраною клітин Шванна й утворення мієліну. Безмієліновими є більшість нейронів безхребетних.
У ссавців нейрони головно вкриті мієліном, проте мієлін тут утворений здебільшого не клітинами Шванна, а олігодендроцитами (див. рис. 2-3). На відміну від клітин Шванна, які формують мієлінову оболонку між двома вузловими перетяжками Ранв’є одного нейрона, олігодендроцити поширюють свої відростки, що утворюють мієлін, до багатьох суміжних аксонів. У разі розсіяного склерозу, важкого автоімунного захворювання, у ЦНС відбувається багато-вогнищева деструкція мієліну. Втрата мієліну супроводжується сповільненням або припиненням передавання імпульсів демієлінізованими аксонами.
Розміри окремих нейронів справді вражають. Наприклад, якщо припустити, що тіло клітини спинномозкового нейрона, який іннервує м’язи стопи, має розмір тенісного м’ячика, то його дендрити можуть заповнити середнього розміру житлову кімнату, а аксон простягнеться на 1,6 км, будучи 13 мм у діаметрі. Загальноприйняті назви частин нейрона зрозумілі, якщо вони стосуються спинномозкових рухових нейронів та інтернейронів, проте виникають деякі незручності з використанням термінів дендрит і аксон стосовно інших типів нейронів. З функціонального погляду (див. нижче, а також Розділи 4 та 5) нейрони переважно складаються з чотирьох головних зон: рецепторної, або дендритної, зони, яка інтегрує множинні локальні зміни потенціалу, генеровані синаптичними зв’язками (рис. 2-4); зони, яка генерує потенціали дії, що поширюються (початковий сегмент спинномозкових рухових нейронів, початкова вузлова перетяжка Ранв’є шкірних чутливих нейронів); відросток аксона, який передає імпульси, що поширюються до нервових закінчень; і нервові закінчення, де потенціали дії зумовлюють вивільнення синаптичних нейротрансмітерів.
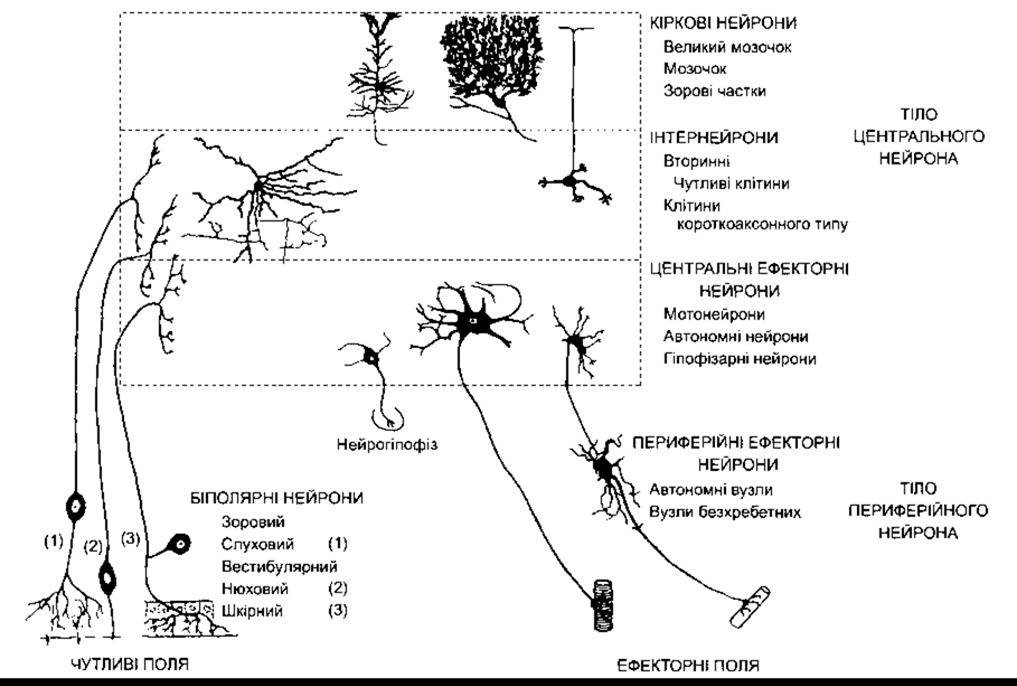
Рис. 2-1. Окремі типи нейронів нервової системи ссавців (відтворено за дозволом, з Bodian D. Introductory survey of neurons. Cold Spring Harbor Symp Quant Biol. 1952; 17:1).
Тіло клітини в більшості випадків розміщене в дендритній зоні кінця аксона, проте воно може бути і всередині аксона (наприклад, у слухових нейронах) або збоку від нього (наприклад, у шкірних нейронах; див. рис. 2-1). Розміщення тіла нейрона не впливає на рецепторну функцію дендритної зони і функцію передавання аксоном імпульсу.
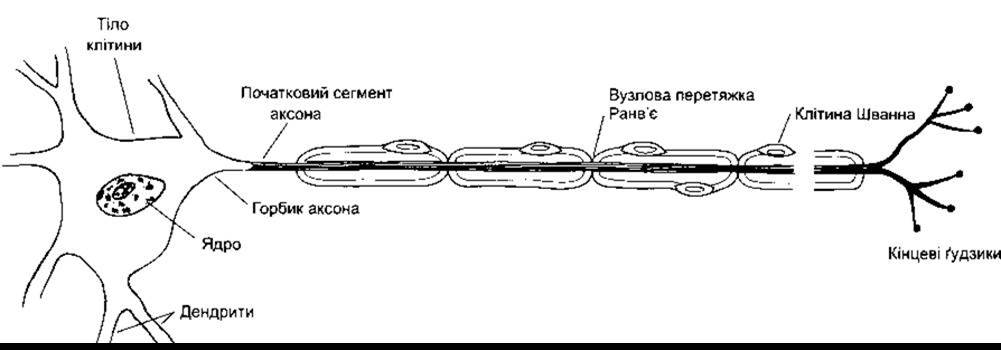
Рис. 2-2. Руховий нейрон з мієлінізованим аксоном.
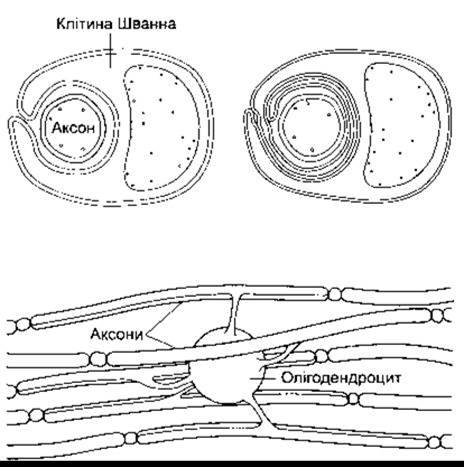
Рис. 2-3. Угорі: співвідношення клітин Шванна з аксонами периферійних нервів. Ліворуч зображено переріз безмієлінового аксона, а праворуч - мієлінового. Як бачимо, клітинна мембрана клітини Шванна згортається й охоплює аксон. Унизу: мієлінізація аксонів ЦНС олігодендроцитами. Один олігодендроцит поширює відростки понад 40 аксонів.
Зазначимо, що дендритні розгалуження нейронів помітно відрізняються за розмірами і складністю структури (рис. 2-1; див. також рис. 11-1 і 12-17). Окрім інтегрування пасивної електричної активності, дендрити в окремих випадках можуть генерувати і поширювати потенціал дії.
Синтез білків і аксонне транспортування
Нервові клітини належать до секреторних, проте відрізняються від інших клітин цього типу тим, що їхня секреторна зона розміщена переважно у кінці аксона, а тому значно віддалена від тіла клітини. Рибосом в аксоні і його терміналах небагато або й зовсім нема, синтез усіх потрібних білків відбувається в ендоплазматичній сітці та апараті Ґольджі, і лише згодом аксон транспортує їх до синаптичних ґудзиків аксоплазматичним транспортуванням. Отже, тіло нервової клітини забезпечує функційну і структурну цілісність аксона: у випадку перерізання аксона його дистальна частина підлягає переродженню (дегенерація Валлєра). Антероградне транспортування забезпечують мікротрубочки; його молекулярні механізми описані в Розділі 1. Швидке антероградне транспортування відбувається зі швидкістю приблизно 400 мм за добу, а повільне - близько 0,5-10,0 мм за добу. Ретроградне транспортування, тобто переміщення речовин у протилежному напрямі, має швидкість приблизно 200 мм за добу.
Синаптичні пухирці відновлюються в мембрані, проте деякі використані переміщуються до тіла нервової клітини і накопичуються в лізосомах. Окремі речовини, наприклад фактор росту нервів, а також різноманітні віруси, що проникають у нервові закінчення шляхом ендоцитозу, теж потрапляють до тіла нервової клітини внаслідок ретроградного аксонного транспортування.