Фізіологія людини - Вільям Ф. Ґанонґ 2002
Фізіологія нервових і м'язових клітин
Збудлива тканина: нервова
Збудження і провідність
Нервовим клітинам властивий низький поріг збудження. Подразник може бути електричного, хімічного або механічного походження. Відомі два типи реакцій на фізико-хімічні подразники: місцева, яка не поширюється, залежно від місця виникнення її називають синаптичною, генераторною або електротонічними потенціалами; і така, що поширюється - потенціали дії, або нервові імпульси. Для нервів та інших збудливих тканин характерні лише реакції електричного походження, що є головною «мовою» нервової системи. Вони зумовлені змінами проникності клітинної мембрани для іонів, що залежать від стану йонних каналів. Аксон передає нервовий імпульс до свого закінчення. Нерви не передають імпульси пасивно, як це відбувається, наприклад, у телефонній мережі; проходження нервових імпульсів, навіть швидке, є набагато повільнішим, ніж протікання електричного струму дротами. Нервова тканина насправді - це порівняно слабкий пасивний провідник: для отримання сигналу до 1 В на іншому кінці 1-метрового аксона, якщо нема активних процесів у нерві, потрібен багатовольтовий потенціал. Передавання нервового імпульсу в нерві є активним самостійним процесом, який поширюється зі сталою амплітудою і швидкістю. Цей процес часто порівнюють з процесом горіння доріжки, укладеної крупинками пороху: запалювання кожної крупинки спричинює запалювання сусідньої безпосередньо спереду, і полум’я неухильно та рівномірно поширюється вздовж доріжки до її кінця.
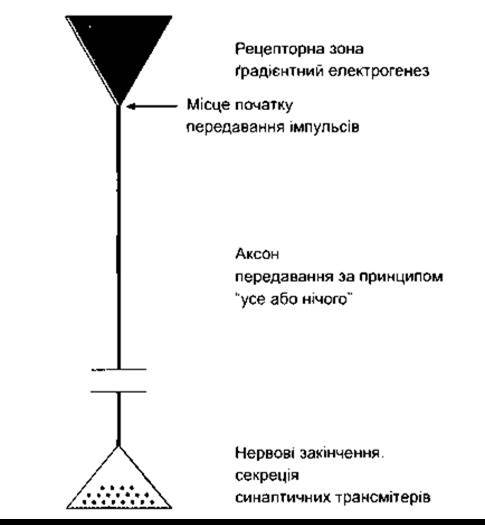
Рис. 2-4. Функційна організація нейрона. Локальні потенціали накопичуються в рецепторній зоні, а потенціал дії виникає в ділянці, що поблизу рецепторної зони (стрілка). Аксон поширює потенціал дії до нервових закінчень, де він зумовлює виділення синаптичних трансмітерів.
Електричні процеси, що відбуваються в нейроні, є швидкоплинними, їх вимірюють у мілісекундах (мс); зміни потенціалів теж незначні; їх вимірюють у мілівольтах (мВ). Поєднання мікроелектродної техніки, у якій застосовують мікроелектроди діаметром до 1 мкм, з використанням електронних підсилювачів і катодного осцилоскопа дає змогу детально досліджувати процеси електричної активності в нервах. Сучасні прилади підсилюють зміни потенціалів у 1000 і більше разів, а катодний осцилоскоп забезпечує майже безінерційне й одночасне відтворення електричних процесів у нерві.
Катодний осцилоскоп
Катодний осцилоскоп (КО) застосовують для вимірювання електричних процесів, що відбуваються в живих тканинах. У КО електрони, що їх випромінює катод, спрямовані у вигляді сфокусованого пучка на екран скляної трубки, у якій розміщений цей катод (рис. 2-5). Екран вкритий однією з численних субстанцій (фосфори), що випромінює світло під впливом електронів. З кожного боку пучка електронів розміщена вертикальна металева пластина. У випадку подання напруги на пластини позитивно заряджена пластина притягує негативно заряджені електрони, які відштовхує негативно заряджена пластина. У разі поступового збільшення напруги, яку подають на вертикальні пластини (Х-пластини), і потім її раптового зменшення і подальшого збільшення, пучок електронів рухається поступово до позитивно зарядженої пластини, згодом повертається до попереднього положення і знову відхиляється до позитивно зарядженої пластини. Застосування такого типу «зубчастої» напруги спричинює відхилення пучка електронів упоперек екрана трубки, швидкість цього відхилення пропорційна до ступеня збільшення застосованої напруги.
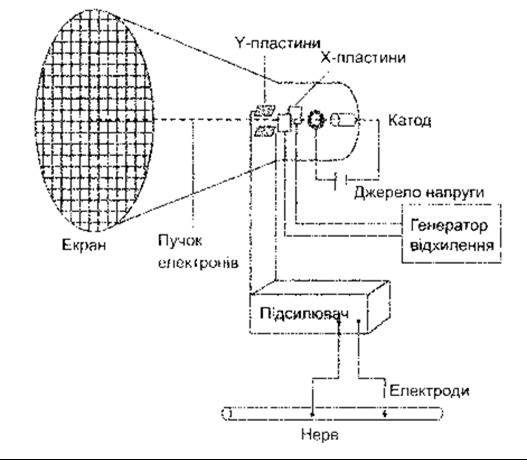
Рис. 2-5. Катодний осцилоскоп Спрощене зображення основної схеми для реєстрації змін потенціалів у нерві.
Інша пара пластин (Y-пластини) розміщена горизонтально, одна пластина вище, а друга нижче від місця проходження пучка електронів. Напруга, яку подають на ці пластини, відхиляє пучок електронів догори і донизу, коли він перетинає впоперек екран трубки. Значення вертикального відхилення пропорційне різниці потенціалів між горизонтально розміщеними пластинами. Якщо ці пластини приєднані до електродів, розміщених на нерві, то можна зафіксувати будь-які зміни потенціалів у нерві у вигляді вертикального напряму відхилень пучка електронів під час його руху впоперек трубки.
Реєстрація змін потенціалів в ізольованих аксонах
Аксони ссавців мають невеликий діаметр (20 мкм або й менше), тому їх важко ізолювати від інших аксонів нерва, тоді як велетенські безмієлінові нервові клітини містяться в значних кількостях в організмі безхребетних і їхнє відокремлення не викликає великих труднощів. Такі велетенські клітини є, наприклад, у крабів (Carcinus) і каракатиць (Sepia), проте найбільшого розміру аксони виявлені в тілі кальмара (Loligo). Шийна ділянка м’язової мантії кальмара містить поодинокі аксони діаметром до 1 мм. Головні властивості цих аксонів близькі до головних властивостей аксонів ссавців.
Мембранний потенціал спокою
Якщо два електроди, розміщені на поверхні ізольованого аксона, приєднати через підсилювач до КО, то прилад не зафіксує зміни напруги. Однак якщо один з електродів увести через мембрану всередину клітини, то прилад зареєструє постійну різницю потенціалів між негативно зарядженим середовищем усередині клітини і позитивно зарядженою поверхнею. Мембранний потенціал спокою властивий майже всім клітинам. Механізм генерації мембранного потенціалу спокою розглянуто в Розділі 1. У нейронах його значення дорівнює приблизно -70 мВ.
Латентний період
У випадку нанесення подразнення і передавання аксоном нервових імпульсів виникають характерні короткочасні зміни потенціалів, які називають потенціалом дії і реєструють у разі досягнення імпульсами поверхневого електрода.
Одразу ж після подразнення виникає короткочасне, змінного характеру, відхилення від базової лінії - артефакт подразника. Цей артефакт зумовлений поширенням електричного струму від подразнювального електрода до реєструвального. Таке явище звичайно простежується, навіть незважаючи на старанне екранування, проте воно має важливе позитивне значення, оскільки реєстрація його на екрані катодної трубки дає змогу точно визначити час виникнення подразнення. Після артефакту подразника настає ізопотенціальний інтервал (латентний період), який завершується з початком потенціалу дії і відповідає часові, упродовж якого імпульс проходить аксоном від точки подразнення до реєструвального електрода. Тривалість цього періоду пропорційна до відстані між подразнювальним і реєструвальним електродами, а також обернено пропорційна до швидкості проходження. Якщо тривалість латентного періоду і відстань між електродами відомі, то можна обчислити швидкість в аксоні. Наприклад, припустимо, що відстань між катодним подразнювальним електродом і розміщеним на поверхні електродом (рис. 2-6) дорівнює 4 см. Постійний електричний струм, що слугує подразником, звичайно подають з катода. Якщо латентний період триває 2 мс, то швидкість проходження - 4 см за 2 мс, тобто 20 м/с.
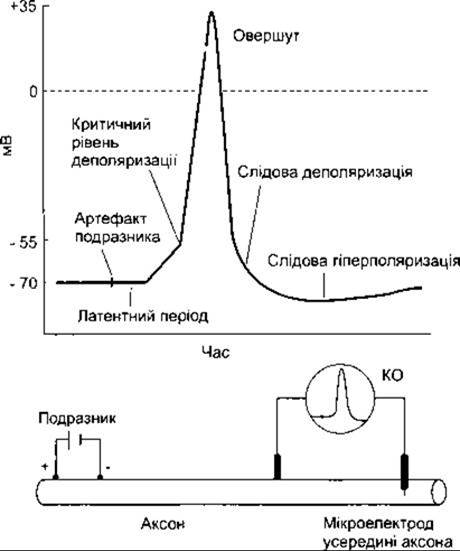
Рис. 2-6. Потенціал дії в нейроні. Один з електродів міститься всередині клітини.
Потенціал дії
Першим проявом виникнення потенціалу дії є деполяризація мембрани. Після початкових 15 мВ швидкість деполяризації зростає. Точку, від якої розпочинається ця зміна швидкості, називають критичним рівнем деполяризації, або порогом. Стосовно до цього, крива, яку відтворює осцилоскоп, швидко досягає і перевищує рівень ізопотенціальної лінії (потенціал нуль), сягаючи приблизно +35 мВ (овершут). Згодом потенціал швидко зменшується до рівня потенціалу спокою. Як тільки реполяризація досягає приблизно 70, швидкість реполяризації знижується і крива наближається до рівня потенціалу спокою повільніше. Швидке збільшення і згодом раптове зменшення потенціалу називають піковим потенціалом аксона, а повільне зменшення наприкінці процесу - слідовою деполяризацією. Після досягнення початкового рівня, що відповідає значенню потенціалу спокою, крива, хоч і незначно, знову підіймається в напрямі гіперполяризації, реєструючи явище слідової гіперполяризації. Потенціал дії, зареєстрований в умовах, коли один електрод перебуває всередині клітини, називають монофазовим, оскільки процес відбувається, головно, в одному напрямі.
Співвідношення частин кривої, зображеної на рис. 2-6, умисно дещо спотворене з метою ліпшої ілюстрації фаз потенціалу дії. Крива, складові частини якої реально відтворюють часові характеристики і значення потенціалу в нейроні ссавців, показана на рис. 2-7. Зазначимо, що збільшення потенціалу дуже швидке і не дає змоги чітко простежити зміни внаслідок деполяризації та критичний рівень деполяризації, а значення слідової гіперполяризації відповідає лише 1-2 мВ, хоча триває цей період майже 40 мс. Тривалість слідової деполяризації в цьому випадку дорівнює приблизно 4 мс. У багатьох нейронах вона ще менш тривала і має мале значення. Зміни в значеннях слідової поляризації можуть відбуватися без суттєвих змін у решті фаз потенціалу дії. Наприклад, якщо нерв упродовж тривалого часу передає імпульси, то значення слідової гіперполяризації звичайно збільшується.
Закон «усе або нічого»
Якщо потенціал дії реєструють в умовах, як це зображено на рис. 2-6 - з розміщенням реєструвальних електродів на значній відстані від електродів, за допомогою яких завдають подразнення, то можна з’ясувати мінімальне значення постійного струму (порогова сила), який, діючи упродовж певного часу, зумовлює виникнення потенціалу дії. Між силою струму і тривалістю його дії є залежність. У разі використання слабкого струму тривалість його дії збільшується, а в разі сильного - зменшується. Співвідношення між силою струму і тривалістю його дії описує спеціальна крива. Повільне збільшення струму не призводить до збудження в нерві, оскільки нерв адаптується до такого типу збудника; цей процес називають акомодацією. У випадку порогової сили подразника виникає потенціал дії. Подальше зростання сили подразника не призводить до збільшення значення чи інших показників потенціалу дії за умови, що інші дані експерименту незмінні. Підпороговий, недостатній, подразник не викликає появи потенціалу дії. Потенціал дії постійної амплітуди і форми незалежно від сили подразника виникає за умови, якщо цей подразник є пороговим або надпороговим. Отже, потенціал дії підлягає закону «все або нічого».
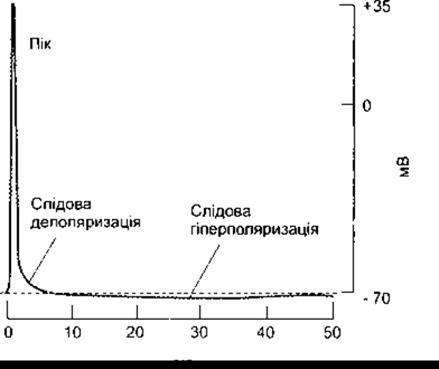
Рис. 2-7. Реєстрація потенціалу дії у великому мієліновому волокні ссавців, що відображає реальне співвідношення його фаз.
Електротонічні потенціали, локальна відповідь і критичний рівень деполяризації
Хоча підпорогові подразники не викликають появи потенціалу дії, вони впливають на мембранний потенціал. Цей ефект можна простежити за умови розміщення реєструвальних електродів на відстані декількох міліметрів від подразнювального електрода і застосування підпорогового подразника упродовж деякого часу. Використання підпорогового струму, який подають з катода, спричинює локальну деполяризувальну зміну потенціалу, значення якого різко збільшується та експоненціально, упродовж певного часу, зменшується. Крива цієї реакції спадає швидше, якщо відстань між подразнювальним і реєструвальним електродами збільшувати. Анодний струм зумовлює таку ж за тривалістю зміну потенціалу в напрямі гіперполяризації. Такого типу зміни потенціалу називають електротонічними потенціалами, а залежно від напряму струму (тобто спричинені дією катода чи анода), - відповідно, кателектротонічними або анелектротонічними. Названі зміни потенціалу - це пасивні зміни поляризації мембрани, зумовлені додаванням або відніманням заряду в разі дії певного електрода. За умови низького значення струму, що спричинює деполяризацію або гіперполяризацію приблизно 7 мВ, їхні значення пропорційні до сили подразника. Якщо подразник значної сили, то ця закономірність справджується для анелектротонічних типів змін потенціалу, проте не виявляється в разі подразнювання з катода; у випадку дії катода зміна потенціалу є сильнішою, ніж можна було б сподіватися, орієнтуючись на силу подразника. Врешті, якщо катодне подразнення є достатньої сили, щоб викликати деполяризацію 15 мВ, за умови, що мембранний потенціал спокою -55 мВ, то мембранний потенціал раптово зменшується і розпочинається поширення потенціалу дії.
Непропорційно високого рівня зміни потенціалу достатнього за силою катодного подразника, що спричинює 7-15 мВ деполяризації, виникають унаслідок відкриття воріт потенціалозалежних Nа+-каналів (див. нижче); їх називають локальною відповіддю (рис. 2-8). Точка, у якій ініційоване різке сходження до пікового потенціалу, є критичним рівнем деполяризації. Отже, катодний струм, який спричиняє деполяризацію рівня 7 мВ, зумовлює винятково пасивну реакцію мембрани як наслідок додавання негативних зарядів. Катодний струм, що спричинює деполяризацію 7-15 мВ, теж незначно впливає на процес деполяризації. Сили реполяризації все ще переважають сили деполяризації, і потенціал зменшується. У разі деполяризації 15 мВ сили деполяризації достатні, щоб переважити сили реполяризації, унаслідок чого виникає потенціал дії.
Подразнення звичайно завдають з катода, оскільки катодні подразники мають деполяризувальний вплив. Анодні струми, зміщуючи мембранний потенціал далі від критичного рівня деполяризації, пригнічують утворення імпульсу. Однак припинення дії анодного струму може спричинити овершут мембранного потенціалу в напрямі деполяризації. Цей рикошет часом буває настільки сильним, що одразу ж після припинення анодного подразнення спричиняє збудження в нерві.
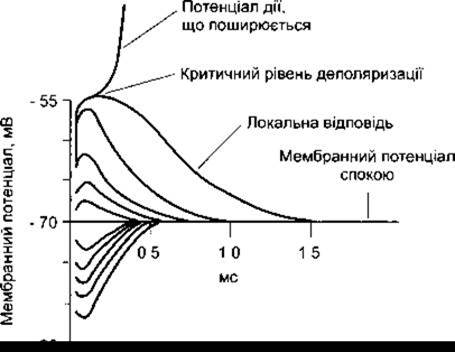
Рис. 2-8. Електротонічні потенціали і локальна відповідь. Криві зміни мембранного потенціалу в нейроні після завдання подразнення, що відповідає значенням 0,2; 0,4; 0,8 та 1,0 порогового струму, розміщені на шкалі часу. Криві реакцій, розташовані нижче від горизонтальної лінії, відтворюють процеси, що виникають у разі дії анодного струму, а криві, що розташовані вище, - у разі дії катодного струму. Подразник порогового рівня використаний двічі. За першим разом він спричинив потенціал дії, а за другим - не спричинив.
Зміна збудливості упродовж електротонічних потенціалів і потенціалу дії
У випадку потенціалу дії, як і в разі кателектротонічних та анелектротонічних потенціалів, а також локальної відповіді змінюється рівень порога подразнення нейрона. Гіперполяризувальні анелектротонічні відповіді підвищують рівень порога, а деполяризувальні кателектротонічні - знижують його, оскільки наближають мембранний потенціал до критичного рівня деполяризації.
Під час локальної відповіді рівень порога знижується, однак упродовж фази підіймання і фаз, що супроводжуються зниженням після пікового потенціалу, нейрон не чутливий до подразнення. Цей рефрактерний період поділяють на період абсолютної рефрактерності, який відповідає часу збільшення потенціалу від критичного рівня деполяризації до точки проходження однієї третини реполяризації, і період відносної рефрактерності, який триває від цієї точки до початку слідової деполяризації.
Під час періоду абсолютної рефрактерності жоден подразник будь-якої сили не викликає збудження нерва, проте під час періоду відносної рефрактерності подразник, більший за силою від застосованого раніше, може це зробити. У разі слідової деполяризації рівень порогової сили знову знижується, а у випадку слідової гіперполяризації - зростає. Залежність рівня порога від фаз потенціалу дії відображена на рис. 2-9.
Електрогенез потенціалу дії
У стані спокою мембрана нервової клітини поляризована. Зовнішня поверхня мембрани має позитивний заряд, а внутрішня - негативний. У разі формування потенціалу дії ця полярність змінюється й упродовж короткого часу стає насправді оберненою (рис. 2-10). Позитивне перезарядження мембрани у двох напрямах попереду і позаду потенціалу дії переходить у ділянку, де містяться негативні заряди, зумовлені потенціалом дії. Завдяки переміщенню позитивних зарядів знижується полярність мембрани попереду потенціалу дії. Така електротонічна деполяризація спричинює локальну відповідь, і, як тільки досягнуто критичного рівня деполяризації, збудження поширюється, що, відповідно, далі допереду електротонічно деполяризує мембрану. Завдяки такій почерговості подій збудження безмієліновим аксоном рівномірно поширюється до його кінця. Отже, здатність нервового імпульсу самопоширюватись зумовлена циркуляцією струму з наступною деполяризацією мембрани аж до критичного рівня попереду потенціалу дії. Поширюючись, нервовий імпульс не деполяризує до критичного рівня ділянку позаду себе, оскільки ця ділянка є рефракторною.
Потенціали дії, що виникають у синаптичних контактах і чутливих закінченнях, теж залежать від електротонічної деполяризації мембрани нервової клітини до критичного рівня (див. Розділи 4 та 5).
Механізм сальтаторної провідності
Провідність у мієліновому аксоні залежить від подібних процесів поширення струму. Однак мієлін є ефективним ізолятором, тому циркуляція струму крізь нього незначна.

Рис. 2-9. Зміни збудливості клітинної мембрани нейрона під час поширення нервового імпульсу. Як видно, збудливість обернено пропорційна до рівня порога (модифікована схема, відтворена за дозволом з Morgan СТ. Physiological Psychology. McGraw-Hill, 1943).
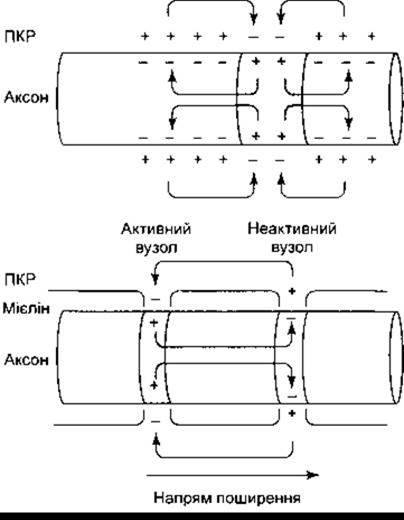
Рис. 2-10. Локальний струм в аксонах / рух позитивних зарядів навколо імпульсу. Угорі: безмієліновий аксон. Унизу: мієліновий аксон; ПКР - позаклітинна рідина.
Ось чому деполяризація в мієліновому аксоні поширюється стрибкоподібно, наче сальто, від одного вузла нервового волокна (перетяжка Ранв’є) до іншого зі згасанням струму в активному вузлі, що спричинює електротонічну деполяризацію аж до критичного рівня у вузлі, розміщеному попереду потенціалу дії (див. рис. 2-10). Такого типу поширення деполяризації від вузла до вузла називають сальтаторною, або стрибкоподібною провідністю. Це швидкий процес, провідність у мієліновому аксоні відбувається у 50 разів швидше, ніж провідність у найшвидшому безмієліновому волокні.
Ортодромна й антидромна провідність
Аксон здатний передавати збудження в обох напрямах. Якщо потенціал дії ініційовано посередині аксона, то поширення імпульсів в обох протилежних напрямах зумовлює електротонічна деполяризація з кожного боку від місця подразнення.
Проте в живому організмі імпульси звичайно поширюються лише в одному напрямі, тобто від синаптичного контакту або рецепторів аксоном до його кінця. Таку провідність називають ортодромною. Провідність у протилежному напрямі - антидромна. Оскільки синапси, на відміну від аксона, забезпечують провідність лише в одному напрямі, то будь-які антидромні імпульси, що утворюються, не спроможні пройти крізь уже перший синапс, який трапляється на їхньому шляху, і згасають.
Двофазовий характер потенціалу дії
Відмінності мембранного потенціалу спокою від потенціалу дії, описані раніше, ґрунтуються на записуванні потенціалів за допомогою двох електродів: один з них розміщений на поверхні аксона, а інший - усередині. Якщо обидва реєструвальні електроди розмістити на поверхні аксона, то осцилоскоп не зафіксує різниці потенціалів між ними в стані спокою. Коли нерв подразнюють, то поза цими двома електродами відбувається проходження імпульсу, що зумовлює характерні послідовності змін потенціалу. Як тільки хвиля деполяризації сягає ближчого до місця подразнення електрода, - першого, - то цей електрод реєструє негативний заряд щодо другого (рис. 2-11). Якщо ж імпульс поширюється в частині нерва між двома електродами, то осцилоскоп зареєструє відсутність потенціалу, тобто ізолінію. В разі надходження імпульсу до другого електрода, він зареєструє другий потенціал і перший стає позитивно зарядженим щодо другого. Загальноприйнято приєднувати до підсилювача електроди так, щоб перший був негативно зарядженим щодо другого, і тоді запис фіксує відхилення догори. Далі настає ізоелектричний інтервал і після нього крива відхиляється донизу. Таку послідовність називають двофазовим потенціалом дії (див. рис. 2-11). Тривалість ізоелектричного інтервалу пропорційна до швидкості провідності в нерві і відстані між двома реєструвальними електродами.
Провідність в об’ємному провіднику
Оскільки рідини організму містять значну кількість електролітів, то нерви в організмі діють в оточенні добре провідного середовища, яке називають об’ємним провідником. Описані вище моно- і двофазний потенціали дії досліджені в умовах непровідного середовища поза організмом. Зміни потенціалів, які реєструють у разі зовнішньоклітинного дослідження, в оточенні об’ємного провідника, тобто за умов цілісного організму, головно відповідають описаним вище, проте в цьому випадку виникають перешкоди, спричинені поширенням струму через об’ємний провідник. Ці перешкоди зумовлені впливом чинників, пов’язаних з орієнтуванням електродів стосовно напряму поширення потенціалу дії, відстанню між записувальним електродом, що перебуває над активною тканиною, та індиферентним електродом. Загалом під час реєстрації потенціалу дії в середовищі об’ємного провідника простежуються електропозитивні відхилення в обох напрямах від електронегативного піка.
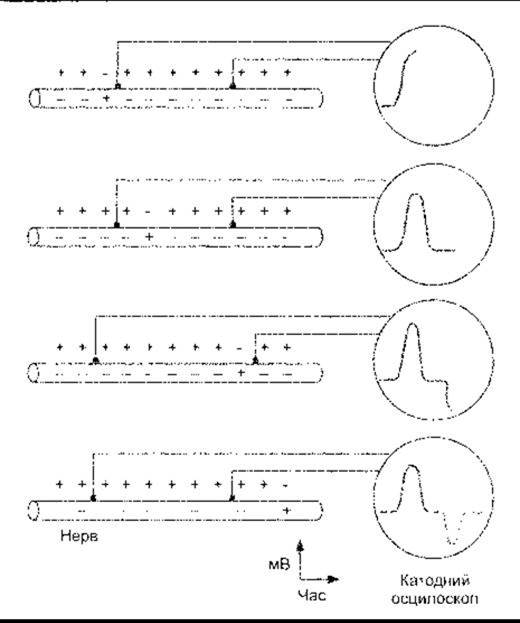
Рис. 2-11. Двофазовий потенціал дії. Обидва реєструвальні електроди розміщені на поверхні мембрани нейрона.