Фізіологія людини - Вільям Ф. Ґанонґ 2002
Фізіологія нервових і м'язових клітин
Збудлива тканина: нервова
Йонні основи збудження і провідності
Клітинні мембрани нейронів, як і інших клітин, мають багато різного типу йонних каналів. Деякі з них пасивні, тобто постійно відкриті, тоді як інші є потенціалозалежними, а ще інші - ліґандозалежними. Функціюванням цих каналів, зокрема Na+- і К+-каналів, пояснюють електричні явища в нервах.
Йонні основи мембранного потенціалу спокою
Як зазначено в Розділі 1, з нейронів та інших клітин відбувається активне транспортування Na+, тоді як у клітини - К+. Шляхом дифузії К+ виходить з клітин, a Na+ дифундує в клітини, проте завдяки К+ -каналам проникність мембрани для К+ є значно вищою, ніж для Na+. Оскільки мембрана непроникна для більшості аніонів клітини, то вихід К+ з клітин не супроводжується відповідним відтіканням аніонів, завдяки чому виникає поляризація мембрани. Її зовнішня поверхня має позитивний, а внутрішня - негативний заряд.
Йонні потоки під час потенціалу дії
Зміни проникності мембрани для Na+ і К+, що відбуваються протягом потенціалу дії, відображені на рис. 2-12. Проникність для йона обернено пропорційна до його електричної опірності в мембрані і є мірою проникності цього йона.
Незначне зменшення мембранного потенціалу спокою спричинює відтікання К+ і надходження Сl-, що забезпечує відновлення мембранного потенціалу спокою. Однак коли деполяризація сягає 7 мВ, то прохідність потенціалозалежних Na+-каналів збільшується (активування Na+-каналів), і як тільки досягнуто критичного рівня деполяризації, проникність мембрани для Na+ збільшується настільки, що він лавиноподібно надходить у волокно, створюючи високу концентрацію та електричні градієнти, що тимчасово перекривають реполяризувальні чинники. Потенціал рівноваги для Na+ у нейронах ссавців, обчислений за рівнянням Нернста, дорівнює приблизно +60 мВ. Мембранний потенціал наближається до цього значення, проте упродовж потенціалу дії його не досягає, головно тому, що збільшення проникності для Na+ є короткочасним. Ворота Na+-каналів швидко закриваються (стан інактивування), це триває декілька мілісекунд - до відловлення мембранного потенціалу спокою. Крім того, напрям електричного градієнта для Na+ змінюється на протилежний під час овершута тому, що змінюється мембранний потенціал, і це обмежує надходження в аксон Na+. Третім чинником, що забезпечує реполяризацію, є активування відкриття воріт К+-каналів, яке триває довше, аніж активування Na+-каналів, і, отже, максимум провідності для К+ настає пізніше, ніж для Na+. Ефективний рух позитивних зарядів назовні клітини, зумовлений виходом йонів К+, завершує процес реполяризації. Явище слідової гіперполяризації пояснюють повільним інактивуванням К+- каналів.
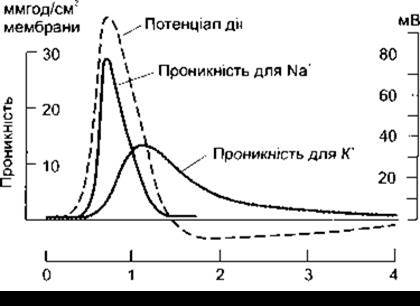
Рис. 2-12. Зміни проникності мембрани для Na+ і К+ у разі формування потенціалу дії у велетенському аксоні кальмара. Штрихова лінія, що відтворює криву потенціалу дії, нанесена на шкалу часу. Як бачимо, початкова електротонічна деполяризація ініціює зміну проникності для Na+, а це, відповідно, сприяє деполяризації (відтворено, за дозволом, з Hodkin AL. Ionic movements and electrical activity in giant nerve fibers. Proc R Soc Long Ser В 1958; 143.1).
Зменшення позаклітинної концентрації Na+ спричинює зменшення потенціалу дії, проте воно незначно впливає на мембранний потенціал спокою. Незначний вплив на мембранний потенціал спокою можна пояснити на підставі рівняння Ґольдмана (див. Розділ 1), оскільки проникність мембрани для Na+ в стані спокою порівняно невелика. І навпаки, збільшення позаклітинної концентрації К+ знижує мембранний потенціал спокою.
Хоча під час формування потенціалу дії Na+ проникає в нервові клітини, a K+ виходить з них, кількість йонів, задіяних у цих переміщеннях незначна щодо їхньої загальної кількості.
Факт, що нерв протягом активності отримує Na+ і втрачає К+, досліджено експериментально, проте помітну різницю виявлено лише в дослідах з тривалим багаторазовим подразненням.
Повільне відкривання та закривання воріт К+-каналів пояснює також явище акомодації аксона. Якщо деполяризація відбувається швидко, то відкривання Nа+-каналів зумовлює перевершення сил реполяризації, якщо ж індукована деполяризація повільна, то відкривання К+-каналів встигає зрівноважити поступове в цьому випадку відкривання Nа+-каналів, і потенціал дії не виникає.
Зменшення позаклітинної концентрації Са2+ посилює збудливість нервових і м’язових клітин шляхом зниження рівня деполяризації, що потрібно для ініціювання змін і проникності для Na+ й К+ та виникнення потенціалу дії. Навпаки, збільшення позаклітинної концентрації Са2+ стабілізує мембрану шляхом зниження збудливості.
Розподіл йонних каналів у мієлінових нейронах
Як зазначено в Розділі 1, деякі речовини, що специфічно пов’язані з Na+- і К+-каналами, можуть бути сполучені, позначені і застосовані для ідентифікування локалізації каналів у клітинній мембрані. Потенціалозалежні Nа+-канали сконцентровані у вузлах нервового волокна Ранв’є і початкових сегментах мієлінових нейронів. Початковий сегмент і перший вузол Ранв’є в чутливих нейронах є ділянками, де генеруються імпульси. Інші вузли Ранв’є - це ділянки, у яких відбувається сальтаторна провідність. Кількість Na+-каналів на 1 мкм2 мієлінового нейрона ссавців становить 50-75 на тілі клітини, 350-500 у початковому сегменті, до 25 на поверхні мієліна, 2000- 12000 у ділянці вузла Ранв’є і 20-25 у терміналах аксона. На аксонах безмієлінових нейронів ця кількість становить приблизно 110. У багатьох мієлінових нейронах Na+-Kaнали межують з К+-каналами, які відіграють важливу роль у процесі реполяризації.
Джерела енергії та метаболізм нерва
Більша частина енергетичних потреб нерва - приблизно 70% - потрібна для підтримання стану поляризації мембрани за допомогою активності Nа+-К+-АТФ-ази. Під час максимальної активності рівень метаболізму в нерві подвоюється (для порівняння: рівень метаболізму в скелетному м’язі збільшується в 100 разів). Гальмування утворень молочної кислоти не впливає на функцію м’яза.
Як і в м’язі, у нерві відбувається теплоутворення в стані спокою; початкове теплоутворення - під час потенціалу дії і затримане - після активності. Однак у нерві затримане теплоутворення, що виникає після поодинокого імпульсу, в 30 разів перевищує початкове. Є докази, що початкове теплоутворення відбувається протягом слідової деполяризації активніше, ніж під час піка потенціалу. Метаболізм у м’язах детально описаний у Розділі 3.