Фізіологія людини - Вільям Ф. Ґанонґ 2002
Ендокринна система, метаболізм і репродуктивна функція
Статева диференціація і розвиток
Жіноча статева система - Гормони яєчника
Хімічна будова, біосинтез та метаболізм естрогенів
Естрогенами природного походження є 17β-естрадіол, естрон і естріол (рис. 23-31). Це С18-стероїди (див. рис. 20-7), тобто вони не мають бокової метальної групи у 10-му положенні чи ∆4-3-кето конфігурації в А-кільці. Головно їх секретують гранульозні клітини фолікулів яєчників, жовте тіло і плацента. Шлях біосинтезу передбачає їхнє утворення з андрогенів. Вони також формуються шляхом ароматизації андростендіону у кров’яному руслі. Ароматаза (CYP19) - це ензим, який каталізує перетворення андростендіону в естрон і тестостерону в естрадіол (див. рис. 23-31). Внутрішня тека клітин багата на рецептори ЛГ; ЛГ діє через цАМФ, збільшуючи перетворення холестеролу в андростендіон. Частина андростендіону перетворюється в естрадіол, який потрапляє в кровообіг. Клітини внутрішньої теки постачають андростендіон у гранульозні клітини, які утворюють естрадіол у разі забезпечення їх андрогенами (рис. 23-32). З’ясовано, що естрадіол, який вони утворюють у приматів, секретується у фолікулярну рідину. Ґранульозні клітини мають багато рецепторів ФСГ, який підтримує секретування ними естрадіолу, діючи через цАМФ для підвищення їхньої ароматазної активності. Зрілі гранульозні клітини набувають також рецепторів ЛГ, а ЛГ стимулює продукування естрадіолу.
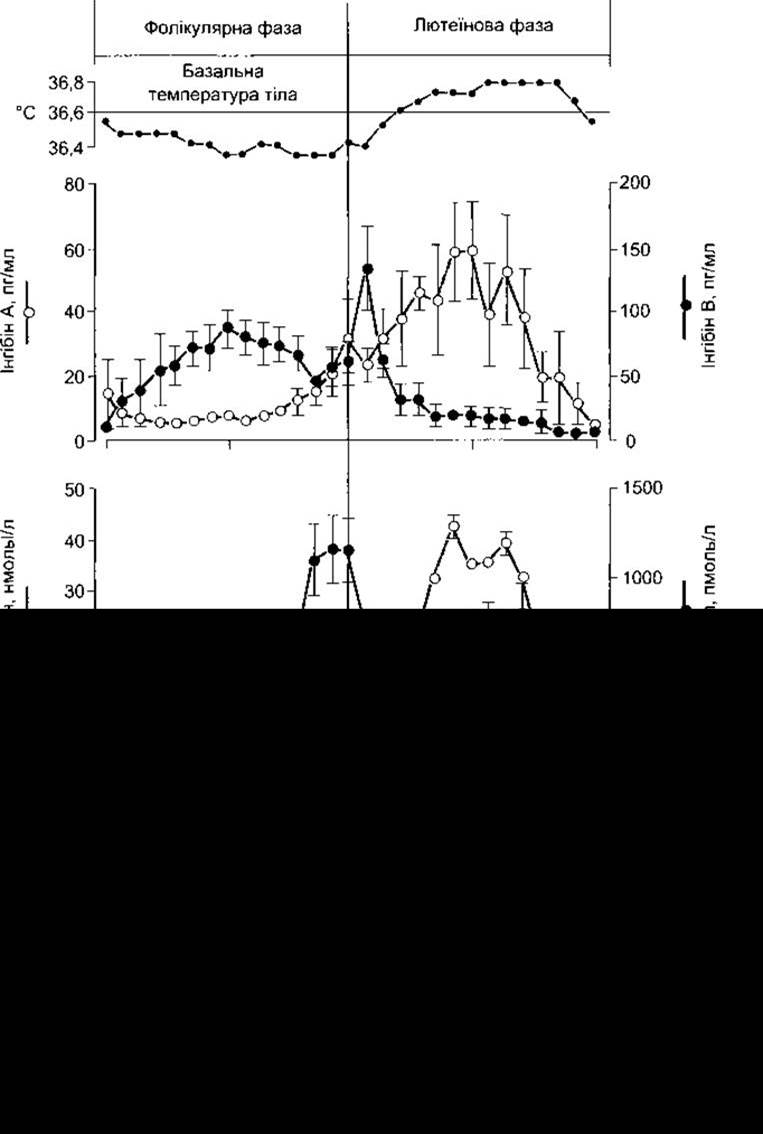
Рис. 23-30. Базальна температура тіла і концентрація гормонів у плазмі (значення ± стандартна похибка) під час нормального місячного циклу людини. Показники розташовані стосовно дня максимального рівня ЛГ у середині циклу.
Стромальна тканина яєчників здатна до утворення андрогенів та естрогенів. Імовірно, вони так і чинять, виділяючи непомітні кількості в пременопаузальних жінок у нормі. У крові 17ß-естрадіол - головний естроген, який секретується - перебуває в стані рівноваги з естроном. У подальшому естрон метаболізує до естріолу переважно у печінці. Зазначимо, що17β-естрадіол є найактивнішим із трьох естрогенів, а естріол - найслабшим. Два відсотки циркулювального естрадіолу є у вільній формі, а решта зв’язана з білком: 60% з альбуміном, 38% із глобуліном, що зв’язує гонадні стероїди (GBG - від англ. gonadal steroid-biuding globulin), який зв’язує тестостерон (див. табл. 23-5).
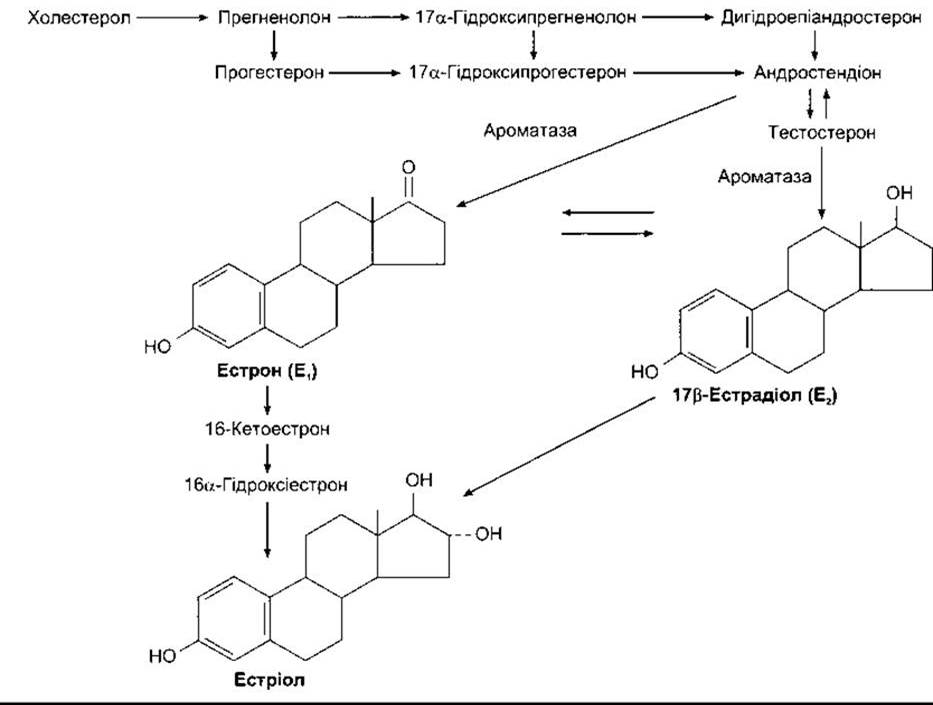
Рис. 23-31. Біосинтез та метаболізм естрогенів. Формули попередників стероїдів показані на рис. 20-8.
У печінці естрадіол, естрон та естріол перетворюються у глюкоронові і сульфатні кон’югати. Ці сполуки виділяються з сечею, як і інші метаболіти. Значна кількість секретується в жовчі та реабсорбується в крові (ентерогепатична циркуляція).
Секреція
Концентрація естрадіолу у плазмі під час місячного циклу показана на рис. 23-30. Практично весь цей естроген походить з яєчників. Виділяють два періоди максимальної секреції: безпосередньо перед овуляцією та усередині лютеїнової фази. Рівень виділення естрадіолу становить 36 мкг/добу (133 нмоль/добу) у ранню фолікулярну фазу, 380 безпосередньо перед овуляцією і 250 мкг/добу у середині лютеїнової фази (табл. 23-7). Після менопаузи виділення естрогену значно знижується.
Як зазначено вище, рівень естрадіолу у чоловіків становить 50 мкг/добу (184 нмоль/добу).
Вплив на жіночі статеві органи
Естрогени підтримують ріст фолікулів у яєчниках та посилюють рухливість маткових труб. Їхній вплив на циклічні зміни в ендометрії, шийці та піхві розглянуто вище. Вони також збільшують маткову кровотечу і впливають на гладку мускулатуру матки, що дуже важливо. Незрілі і кастровані жінки мають маленьку матку й атрофований та неактивний міометрій. Естрогени збільшують м’язову масу матки і вміст у ній скоротливих білків. Під їхнім впливом м’язи стають активнішими та збудливішими. Потенціали дії волокон частішають (див. Розділ 3). «Естроген-домінована» матка чутливіша до дії окситоцину.
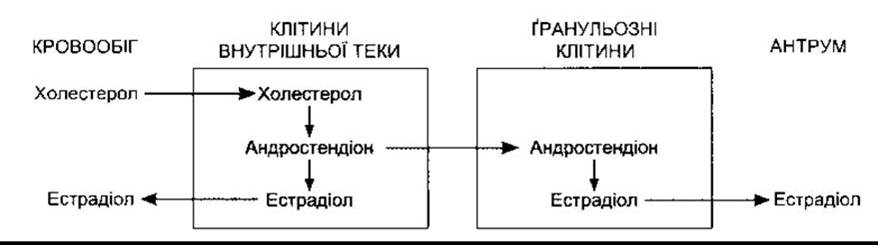
Рис. 23-32. Взаємодія між текою та ґранульозними клітинами у синтезуванні та секретуванні естрадіолу.
Таблиця 23-7. Показники 24-годинного продукування статевих стероїдів у жінок під час різних стадій місячного циклу
Статеві стероїди |
Стадії |
||
рання фолікулярна |
преовуляторна |
середина лютеїнової |
|
Прогестерон, мг |
1,0 |
4,0 |
25,0 |
17-Гідроксипрогестерон, мг |
0,5 |
4,0 |
4,0 |
Дигідроепіандростерон, мг |
7,0 |
7,0 |
7,0 |
Андростендіон, мг |
2,6 |
4,7 |
3,4 |
Тестостерон, мкг |
144,0 |
171,0 |
126,0 |
Естрон, мкг |
50,0 |
350,0 |
250,0 |
Естрадіол, мкг |
36,0 |
380,0 |
250,0 |
1 Модифіковано та відтворено за дозволом з Yen SSC, Jaffe RB, Barbieri RL: Reproductive Endocrinology, 4th ed. Saunders, 1999.
Тривале лікування естрогенами веде до гіпертрофії ендометрію. Якщо лікування естрогенами припинити, то виникне синдром відміни - відшарування з кровотечею.
Інколи кровотечі виникають під час лікування, коли естрогени призначають на тривалий час.
Вплив на ендокринні органи
Естрогени знижують секрецію ФСГ; за деяких умов пригнічують секрецію ЛГ (негативний зворотний зв’язок); за інших збільшують виділення ЛГ (позитивний зворотний зв’язок; див. нижче). Деколи жінкам призначають великі дози естрогенів на чотири-шість днів для запобігання вагітності після коїтусу у фертильний період (посткоїтальна контрацепція). У цьому випадку вагітності, очевидно, перешкоджає швидше порушення імплантації заплідненої яйцеклітини, ніж зміни у виділенні гонадотропіну.
Естрогени спричинюють посилене виділення ангіотензиногену (див. Розділ 24) та тиреоїдозв’язувального глобуліну (див. Розділ 18). Вони дають помітний білковий анаболічний ефект у курчат та великої рогатої худоби, можливо, завдяки стимулюванню виділення андрогенів наднирковими залозами. Тому лікування естрогенами часто комерційно використовують для збільшення маси домашніх тварин. У людини вони зумовлюють закриття епіфізів.
Вплив на ЦНС
Естрогени відповідають за еструс-поведінку у тварин, у людини ж вони підвищують лібідо, прямо впливаючи на певні нейрони гіпоталамуса (рис. 23-33). Вплив естрогенів, прогестерону та андрогенів на статеву поведінку розглянуто у Розділі 15.
Крім того, естрогени збільшують проліферацію дендритів на нейронах і кількість синаптичних закінчень у щурів. Зазначено, що у людини вони сповільнюють розвиток хвороби Альцгеймера (див. Розділ 16).
Вплив на молочні залози
Естрогени зумовлюють ріст проток молочних залоз і значно відповідають за збільшення грудей під час пубертату в дівчат. їх називають гормоном росту грудей. Збільшення грудей унаслідок місцевого використання естрогеновмісних шкірних кремів відбувається завдяки систематичному всмоктуванню естрогенів, хоча зумовлене також незначним місцевим впливом. Естрогени відповідають за пігментацію ареол, хоча, як звичайно, пігментація стає інтенсивно швидшою під час першої вагітності, ніж у періоді пубертату. Роль естрогенів у загальному регулюванні росту грудей і лактації розглянуто нижче.
Вторинні жіночі статеві ознаки
Зміни в організмі (збільшення грудей, матки і піхви), які виникають у дівчаток під час пубертату, частково зумовлені дією естрогенів, які є фемінізувальними гормонами, а частково - простою відсутністю тестикулярних андрогенів. Жінки мають вужчі плечі і широкі стегна (широкий несучий кут). Така будова тіла на додаток до жіночого типу розподілу жирової клітковини на грудях та сідницях трапляється також у кастрованих чоловіків. У жінок гортань зберігає допубертатні пропорції, і голос залишається високим. Волосся на їхньому тілі менше, а на голові більше, на лобку воно росте характерним трикутником з основою вгорі. Лобкове та аксилярне волосся у жінок росте швидше під дією андрогенів, ніж естрогенів, хоча лікування естрогенами може спричинити його незначний ріст. Андрогени походять із кори надниркових залоз і, менше, з яєчників.
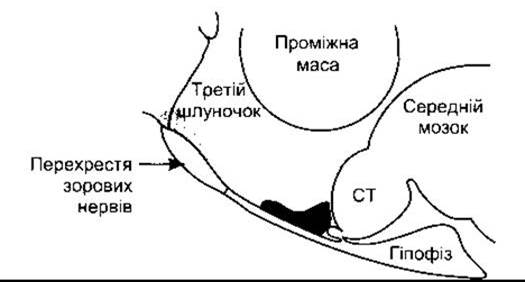
Рис. 23-33. Ділянки, де імплантація естрогену в гіпоталамус впливає на масу яєчників і статеву поведінку у щурів, спроектовані на саґітальному зрізі гіпоталамуса. Імплантанти, які стимулюють статеву поведінку, локалізовані у супрахіазматичній ділянці біля перехрестя зорових нервів (сіра ділянка), тоді як атрофію яєчників спричинюють імплантанти в аркуатному ядрі та навколишньому вентральному гіпоталамусі (кольорова ділянка); СТ - сосочкове тіло.
Інший вплив
У нормі в жіночому організмі відбувається затримка солі та води і збільшення маси тіла безпосередньо перед місячною. Деяку затримку солі і води зумовлюють естрогени. Крім того, простежується дещо посилене виділення альдостерону під час лютеїнової фази, що також приводить до передмісячної затримки рідини.
Естрогени спричинюють розрідження секрету сальних залоз, тому виявляють зворотний до тестостерону ефект і пригнічують утворення вугрів («чорних голівок») та акне. Печінкові долоні, зірчасті гемангіоми і незначне збільшення грудей, що бувають під час прогресування хвороб печінки, зумовлені збільшенням вмісту циркулювальних естрогенів. Таке збільшення зумовлене зниженням печінкового метаболізму андростендіону, який збільшує кількість андрогену, здатного до перетворення в естрогени.
Крім того, естрогени помітно впливають на зниження рівня холестеролу в плазмі (див. Розділ 17). Вони швидко спричинюють вазодилятацію, збільшуючи локальне утворення оксиду азоту (NO) (див. Розділ 31). Це пригнічує атерогенез і приводить до зменшення кількості випадків інфаркту міокарда та інших ускладнень атеросклерозу судин у пременопаузальних жінок. Малі дози естрогенів зменшують кількість серцево-судинних захворювань після менопаузи. Проте великі дози оральних активних естрогенів зумовлюють тромбози, очевидно, тому, що вони досягають печінки у високих концентраціях із портальною кров’ю і змінюють печінкове продукування факторів зсідання крові.
Механізм дії
Подібно до інших стероїдів (див. Розділ 1) естрогени зв’язуються із білковими рецепторами в ядрі. Такі комплекси взаємодіють з ДНК, сприяючи утворенню мРНК, що, відповідно, забезпечує формування нових білків, які регулюють клітинні функції. Клоновано гени двох естрогенових рецепторів: естрогеновий рецептор a (ERa) та естрогеновий рецептор ß (ERß). Незважаючи на подібність, локалізація цих рецепторів різна. Експресія ERa помірна порівняно з його високою експресією у матці, яєчках, гіпофізі, нирках, над’яєчках і надниркових залозах, тоді як експресія ERß висока в яєчниках, простаті, легенях, сечовому міхурі, головному мозку і кістках. Припускають, що регулювання функції яєчників системою гіпофіз-яєчники опосередковане ERa, тоді як естрогени, секретовані у фолікулах яєчників, діють через ERß. Миші чоловічої і жіночої статі, у яких блоковано ген ERa, є стерильними, у них розвивається остеопороз і триває ріст, оскільки їхні епіфізи не закриті. На відміну від цього, миші із блокованим геном ERß нормально розвиваються, розмножуються і лактують, хоча миші-самки народжують меншу кількість дрібніших за розмірами мишенят. Однак потрібно ще виконати додаткові дослідження для детальної характеристики впливу ERa і ERß та їхньої взаємодії.
Більшість дій естрогенів є геномними, тобто опосередковані ERa і ERß. Проте деякі дії дуже швидкі, тому важко повірити, що вони відбуваються завдяки продукуванню мРНК. Сюди можна зачислити дію на нейронні викиди у мозок і, очевидно, зворотний ефект на секрецію гонадотропіну. Такі факти приводять до гіпотези про наявність як негеномного, так і геномного впливу естрогенів, що опосередковані мембранними рецепторами. Подібна швидка дія прогестерону, тестостерону, глюкокортикоїдів та альдостерону може бути також забезпечена мембранними рецепторами.
Синтетичні естрогени
Етинілопохідний продукт естрадіолу (рис. 23-34) є сильним естрогеном, що, на відміну від естрогенів, які виникають природним шляхом, порівняно активний у разі призначення per os, оскільки резистентний до печінкового метаболізму. Активність природних гормонів у випадку призначення per os низька, оскільки портальна венозна сітка кишки переносить їх до печінки, де вони інактивуються, перш ніж потрапляють у загальний кровоплин. Деякі нестероїдні субстанції і декілька сполук, виявлених у рослинах, мають естрогенову активність. Рослинні естрогени зрідка зумовлюють порушення у людини, однак можуть мати небажану дію у тварин. Діетилстильбестрол (див. рис. 23-34) і споріднені сполуки є естрогенними, ймовірно, тому, що вони набувають стероїдоподібної кільцевої структури в організмі. Естрадіол послаблює припливи жару та інші симптоми менопаузи, а також запобігає розвиткові остеопорозу, зменшує прогресування атеросклерозу та випадки серцевих атак і приступів. Однак він стимулює ріст ендометрія та молочних залоз, може призвести до раку матки і молочних залоз. Тому триває активний пошук «чоловічих» естрогенів, які б зумовлювали кістковий і серцево-судинний вплив естрадіолу без ростостимулювального ефекту на матку і груди. Дві сполуки - тамоксифен і ралоксифен - є перспективними щодо цього. Хоча вони не усувають симптомів менопаузи, однак сприяють збереженню кісткової тканини і мають серцево-судинний ефект естрадіолу. Крім того, тамоксифен не стимулює молочні залози, а ралоксифен не стимулює ні молочні залози, ні матку. Клінічне використання цих двох препаратів активно досліджують.
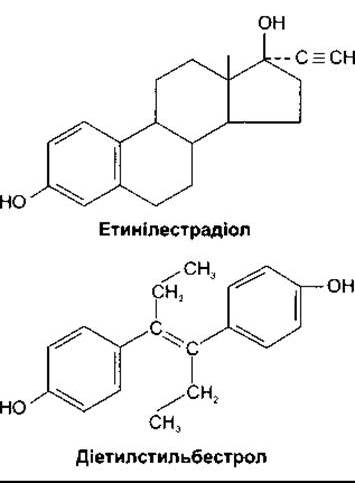
Рис. 23-34. Синтетичні естрогени.
Хімічна будова, біосинтез і метаболізм прогестерону
Прогестерон є С21 -стероїдом (рис. 23-35), що його виділяють жовте тіло, плацента і (в невеликій кількості) фолікул. Він важливий посередник під час біосинтезу стероїдів у тих тканинах, які виділяють стероїдні гормони, невелика його кількість потрапляє в кровоплин з яєчок та кори надниркових залоз. Очевидно, 17а-гідроксипрогестерон виділяється разом з естрогенами із фолікула яєчника, і його секреція відбувається водночас з nß-естрадіолом. Близько 2% циркулювального прогестерону перебуває у вільній формі (див. табл. 23-5), 80% - зв’язано з альбуміном, а 18% - з кортикостероїдозв’язувальним глобуліном (див. Розділ 20). Прогестерон має короткий період півжиття і перетворюється в печінці у прегнандіол, який кон’югує з глюкуроновою кислотою і виводиться з сечею (див. рис. 23-35).
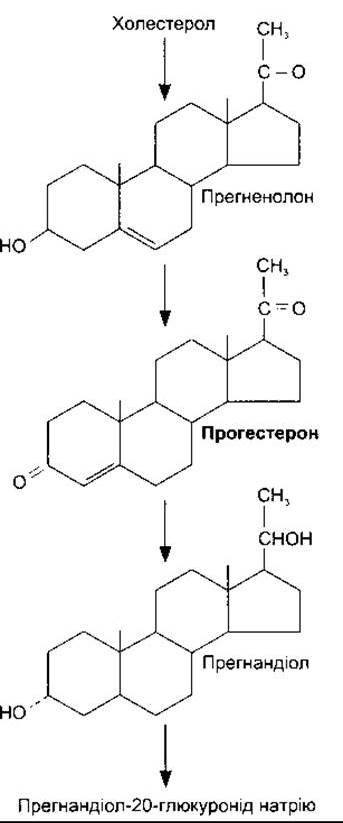
Рис. 23-35. Біосинтез прогестерону та головні шляхи його метаболізму. Утворюються також інші метаболіти.
Секреція
Рівень прогестерону у плазмі чоловіків становить приблизно 0,3 нг/мл (1 нмоль/л), у жінок - близько 0,9 нг/мл (З нмоль/л) під час фолікулярної фази місячного циклу (див. рис. 23-30). Різниця зумовлена тим, що клітини фолікулів яєчника виділяють невелику кількість прогестерону; а клітини теки передають прегненолон гранульозним клітинам, які перетворюють його у прогестерон. У пізній фолікулярній фазі виділення прогестерону посилюється. Під час лютеїнової фази жовте тіло виділяє велику кількість прогестерону (див. табл. 23-7), і в плазмі його рівень помітно підвищується, досягаючи максимального значення - 18 нг/мл (60 нмоль/л).
Стимулювальний вплив ЛГ на виділення прогестерону жовтим тілом відбувається внаслідок активування аденілатциклази і передбачає наступний крок, який залежить від синтезу білка.
Вплив
Головними органами-мішенями прогестерону є матка, молочні залози і мозок. Прогестерон відповідає за зміни в ендометрії і циклічні зміни в шийці та піхві, описані вище. Він має антиестрогенний вплив на клітини міометрія, послаблюючи їхню збудливість, чутливість до окситоцину та спонтанну електричну активність, збільшуючи їхній мембранний потенціал, а також зменшує кількість естрогенових рецепторів в ендометрії та підвищує рівень перетворення nß-естрадіолу до менш активних естрогенів.
У молочних залозах прогестерон стимулює розвиток часточок та альвеол, індукує диференціацію підготовленої естрогенами протокової тканини і забезпечує секреторну функцію молочних залоз під час лактації.
Ефект зворотного зв’язку прогестерону комплексний і виявляється на рівні гіпоталамуса та гіпофіза. Великі дози прогестерону пригнічують секрецію ЛГ і посилюють інгібіторний ефект естрогенів, запобігаючи овуляції.
Прогестерон є термогеном і, очевидно, відповідає за підвищення базальної температури тіла під час овуляції. Він стимулює дихання та альвеолярний РСО2 (РАСО2; див- Розділ 34) у жінок під час лютеїнової фази місячного циклу. Його рівень у цьому разі нижчий, ніж у чоловіків. Під час вагітності РСО2 зменшується, оскільки збільшується виділення прогестерону. Проте фізіологічне значення цієї респіраторної відповіді поки що невідоме.
Великі дози прогестерону призводять до виділення натрію з сечею, очевидно, блокуючи дію альдостерону на нирки. Гормон не створює помітного анаболічного ефекту.
Механізм дії
Дія прогестерону, як і інших стероїдів, зумовлена активністю ДНК щодо ініціації синтезу нових мРНК. Як зазначено у Розділі 1, без стероїда рецептори прогестерону перебувають у комплексі з білком теплового шоку, а після зв’язування прогестерону цей білок відділяється, що веде до експозиції ДНК-зв’язувального домену рецептора. Синтетичний стероїд міфепристон (RU486) зв’язується з рецептором, однак не вивільняє білків теплового шоку, а це блокує зв’язування прогестерону. Оскільки підтримка ранніх стадій вагітності залежить від стимулювального ефекту прогестерону на ріст ендометрія і пригнічення ним скоротливості матки, то міфепристон спричинює викидень. У деяких країнах міфепристон у поєднанні з простагландином використовують для вибіркових переривань вагітності.
Розрізняють дві ізоформи прогестеронових рецепторів, які утворюються шляхом диференційного процесу мРНК одного гена. Прогестероновий рецептор A (PRA) є зсіченою формою, яка після активування здатна пригнічувати деякі дії прогестеронового рецептора В (PRB). Фізіологічне значення двох ізоформ не визначене.
Речовини, які імітують дію прогестерону, деколи ще називають прогестиновими агентами, гестагенами, чи прогестинами. їх використовують разом із синтетичними естрогенами як оральні контрацептивні засоби (див. нижче).
Релаксин
Релаксин - це поліпептидний гормон, що його виробляють жовте тіло, матка, плацента і молочні залози у жінок та передміхурова залоза у чоловіків. Під час вагітності він розслаблює лобковий симфіз та інші з’єднання таза; розм’якшує і розширює шийку матки, отже, допомагає під час пологів. Він також пригнічує скорочення матки і може відігравати роль у розвитку молочних залоз. У невагітних жінок релаксин виявляють у жовтому тілі та ендометрії під час секреторної, однак не проліферативної фази місячного циклу. Його функція у невагітних жінок досі невідома. У чоловіків він міститься у спермі, де, можливо, допомагає підтримувати рухливість сперматозоїдів та їхню пенетрацію в яйцеклітину.
У більшості видів тварин є один ген релаксину, у людини ж виявлено два на 9-й хромосомі. Вони кодують структуру двох структурно відмінних поліпептидів, кожен з яких має активність релаксину. Проте тільки один з цих генів активний у яєчниках і простаті. Структура поліпептидів, що їх виробляють ці дві тканини, показана на рис. 23-36.